
Figure 7.
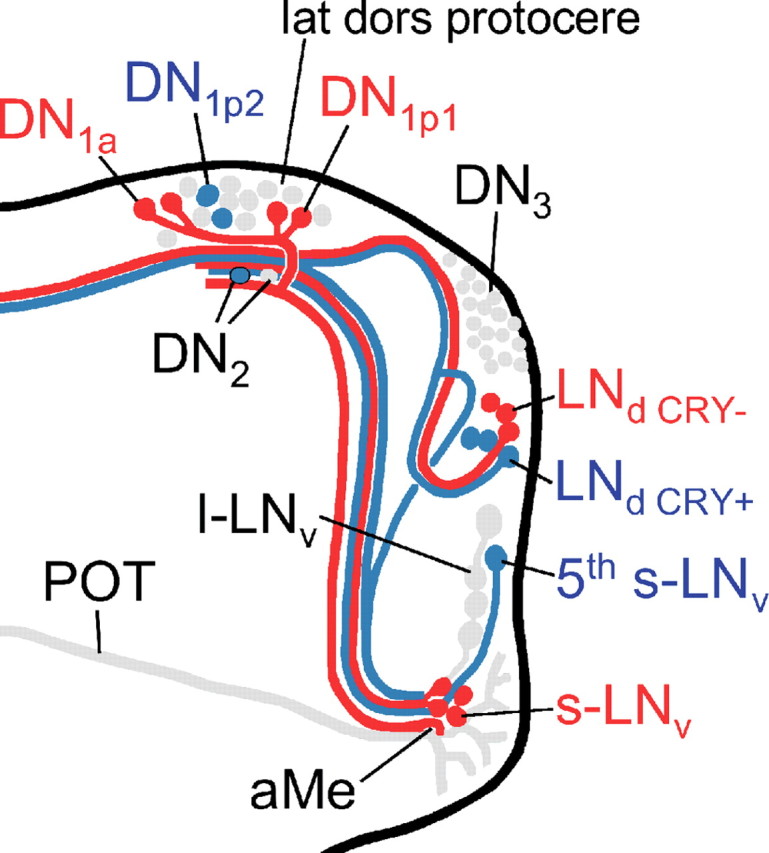
Diagram summarizing our present view on the differential action of PDF on the clock neurons in somda mutants (only the right brain hemisphere is shown). Neurons that lengthen their period on excessive amounts of PDF in the dorsal brain are shown in blue. Neurons that shorten their period under the same conditions are shown in red. Neurons not assessed in the present study or neurons with unclear response to PDF are shown in gray. Note that the aberrant arborizations of the l-LNv are omitted for better clarity. The s-LNv, fifth s-LNv, l-LNv, DN1a, and the three CRY-positive LNd neurons arborize in the aMe, in which PDF secreted by the l-LNv cells might serve as coupling factor in wild-type flies (Wülbeck et al., 2008, their Discussion). Furthermore, fibers from the s-LNv, the DN1a, DN1p1, and the CRY-positive and -negative LNd cells may contact each other in the lateral dorsal protocerebrum (lat dors protocere), in which PDF seems to be rhythmically released from the s-LNv cells (Park et al., 2000). The fifth s-LNv cell was also found to project toward the dorsolateral protocerebrum, but it is not clear whether it reached this area or whether it terminated before (Helfrich-Förster et al., 2007). The l-LNv cells do not project into the dorsolateral brain in wild-type flies, but do so extensively in somda mutants (Wülbeck et al., 2008). We suppose that extensive amounts of PDF secreted into this brain region shorten the periods of the CRY-positive DN1a and DN1p1 cells. The latter two groups may feedback on the s-LNv forcing them also to free-run with short period (for details, see supplemental Discussion, available at www.jneurosci.org as supplemental material).