
Fig. 6: Mechanism enforcing the initiation of stomatal cell lineages via SCRM_KiDoK-MAPK interaction module.
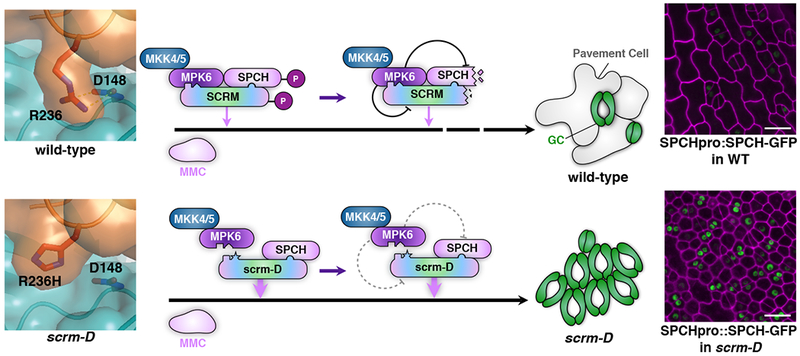
Shown is a schematic diagram depicting the molecular interaction mechanism between MPK3/6, SCRM, and SPCH for proper specification of stomatal cell-fate on the developing epidermis. In wild-type plants, the KiDoK motif of SCRM associates with MPK6 through its bipartite Docking and KRAAM motifs. This association of MPK6 and SCRM triggers subsequent phosphorylation and degradation of the SCRM protein by an unknown proteosomal pathway. Through heterodimerization with SPCH, SCRM brings SPCH to be in molecular proximity with MPK6, thereby allowing the phosphorylation-mediated downregulation of SPCH to inhibit stomatal cell fate. Not depicted in this schematic is the binding mechanism of MPK3 (due to lack of AtMPK3 structural data) with the SCRM KiDoK motif, wherein SCRM primarily utilizes its KRAAM motif to associate with MPK3, that triggers subsequent phosphorylation and degradation of the SCRM protein. In scrm-D, since the scrm-D_KiDoK motif (KHAAM) is unable to associate with MPK6, the scrm-D protein cannot recruit MPK6 to interact with and modulate the stability of itself and SPCH, resulting in a highly-stabilized SPCH protein in the scrm-D background (Right). The absence of proper downregulation of SCRM and SPCH confers constitutive stomatal differentiation in the entire epidermis.