
Figure 8.
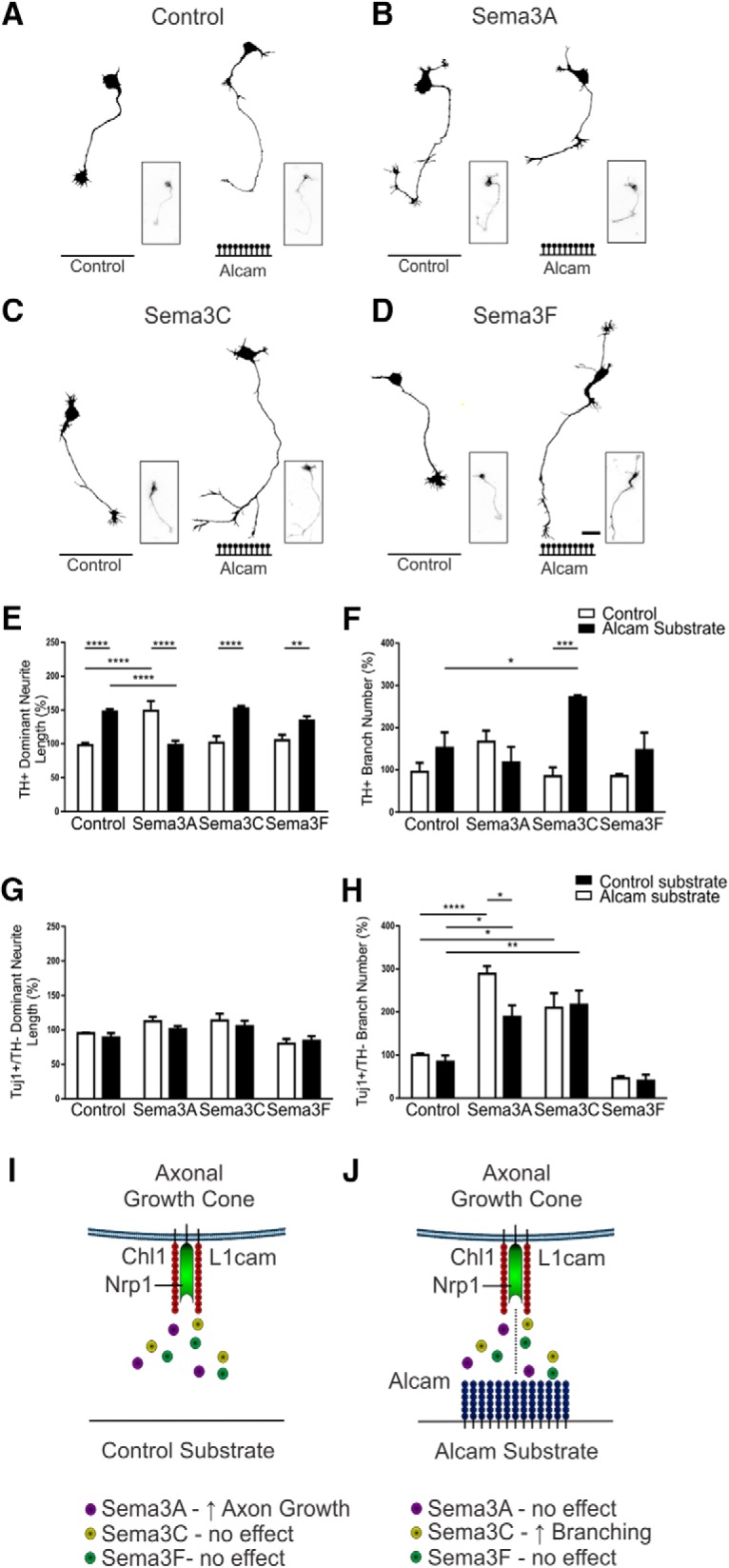
Functional analysis of Alcam substrate modulation of Semaphorin signaling. Representative dopaminergic (TH+) neuron traces of WT primary midbrain neurons cultured on control or Alcam substrate and treated with control (A), Sema3A (B), Sema3C (C), or Sema3F (D). Neurite length (E) of dopaminergic neurons showed a significant interaction (two-way ANOVA, F(3,16) = 28.94, p < 0.0001) and substrate (F(1,16) = 19.59, p = 0.0004) effect (n = 3). When grown on Alcam substrate, neurite length was significantly increased in control (t = 5.580, df = 16, p = 0.0002), significantly decreased with Sema3A treatment (t = 5.677, df = 16, p = 0.0001), significantly increased with Sema3C treatment (t = 5.697, df = 16, p = 0.0001), and significantly increased with Sema3F treatment (t = 3.253, df = 16, p = 0.0198). Direct comparison of Semaphorin-treated neurite length on control substrate significantly increased only with Sema3A treatment (t = 5.729, df = 16, p = 0.0009). The significant growth promoting effect of Sema3A and Alcam substrate on neurite length was reversed in the presence of both Sema3A and Alcam substrate (t = 5.529, df = 16, p < 0.0013). Branch number (F) of dopaminergic neurons showed a significant interaction (two-way ANOVA, F(3,16) = 6.166, p = 0.0055) and substrate (F(1,16) = 10.80, p = 0.0046) effect (n = 3). Branch number significantly increased only in dopaminergic neurons grown on an Alcam substrate and treated with Sema3C (t = 4.808, df = 16, p = 0.0008). Branch number increased significantly in a direct comparison of control and Sema3A-treated neurons grown on Alcam substrate (t = 3.093, df = 16, p = 0.0300), with no significant effect observed in control and Sema3A or Sema3F treated cultures. In nondopaminergic neurons (TH−/Tuj1+), a significant effect on neurite length due to Alcam substrate following Semaphorin treatment (G) (two-way ANOVA, Sema treatment, F(3,16) = 7.213, p = 0.0028, n = 3) was not significant in post hoc tests. A significant effect on branch number due to Alcam substrate following Semaphorin treatment (H) (two-way ANOVA, Sema treatment, F(3,16) = 37.66, p < 0.0001, n = 3) was also observed. Sema3A treatment resulted in a significant decrease in branch number (H) in nondopaminergic neurons when grown on an Alcam substrate (t = 3.260, df = 16, p = 0.0195). Branch number increased significantly in a direct comparison of control and Sema3A-treated neurons grown on control substrate (t = 6.142, df = 16, p = 0.0004), but not in Alcam substrate (t = 3.404, df = 16, p = 0.0968). Schematic of the regulation of Semaphorin signaling the absence (I) or presence (J) of Alcam substrate indicating the proposed trans-heterophilic interactions with the semaphorin receptor complex (comprised of Nrp1, L1cam and Chl1) on the growth cone. Scale bars in A–D, 20 μm. Data are shown as mean ± SEM. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001.