
Figure 6. A model for microvillar motility in brush border assembly.
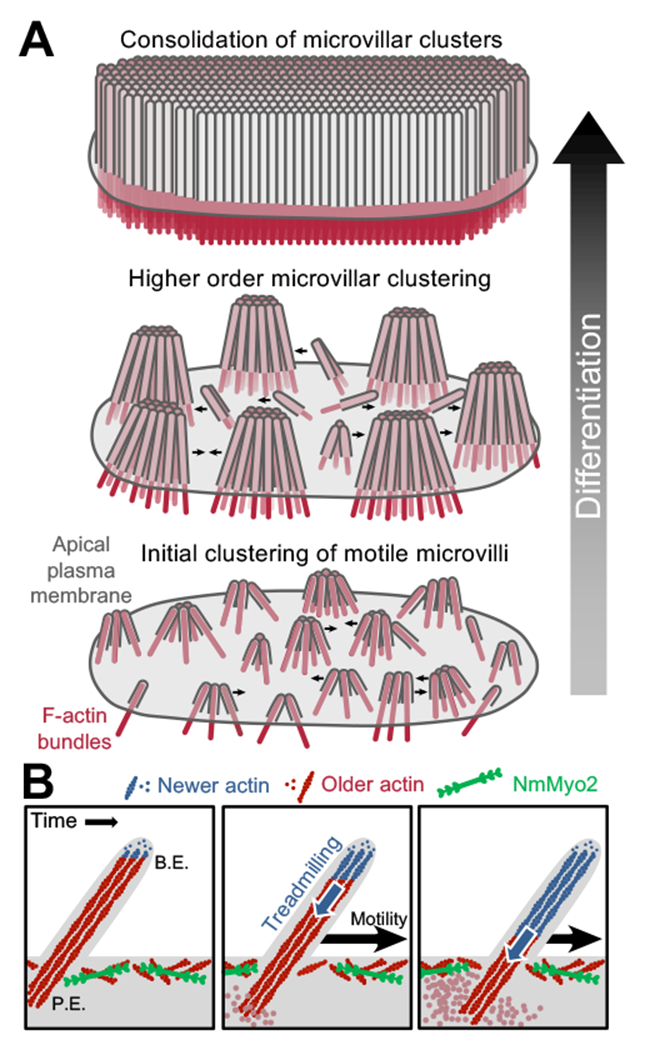
(A) A progressive clustering model for microvillar remodeling and brush border formation during differentiation. Here, nascent microvilli emerge from the apical surface and undergo persistent motility, which promotes collisions between protrusions. These encounters allow adhesion links to form between microvillar tips, resulting in characteristic tepee-shaped structures (originally reported in Crawley et al. Cell 2014). As maturation proceeds, clusters grow by moving across the apical surface, colliding with other clusters and consolidating their numbers until eventually the entire surface is occupied by one continuous large-scale cluster, i.e. a mature brush border. (B) Enlarged view of a single motile microvillus. The microvillar core is comprised of bundled F-actin (red) with the barbed-ends oriented toward the distal tip. New actin monomers incorporate at the barbed-ends (B.E., blue F-actin), which is balanced by monomer disassembly from the pointed-ends (P.E.). This results in ‘treadmilling’ of actin through the microvillar core which provides a pushing force against the membrane that powers microvillar motility. NmMyo2 is localized at the sub-apical cortex and may interact with microvilli rootlets to provide a counterforce translating microvillus actin core treadmilling into movement across the cell surface.