
Fig. 9.
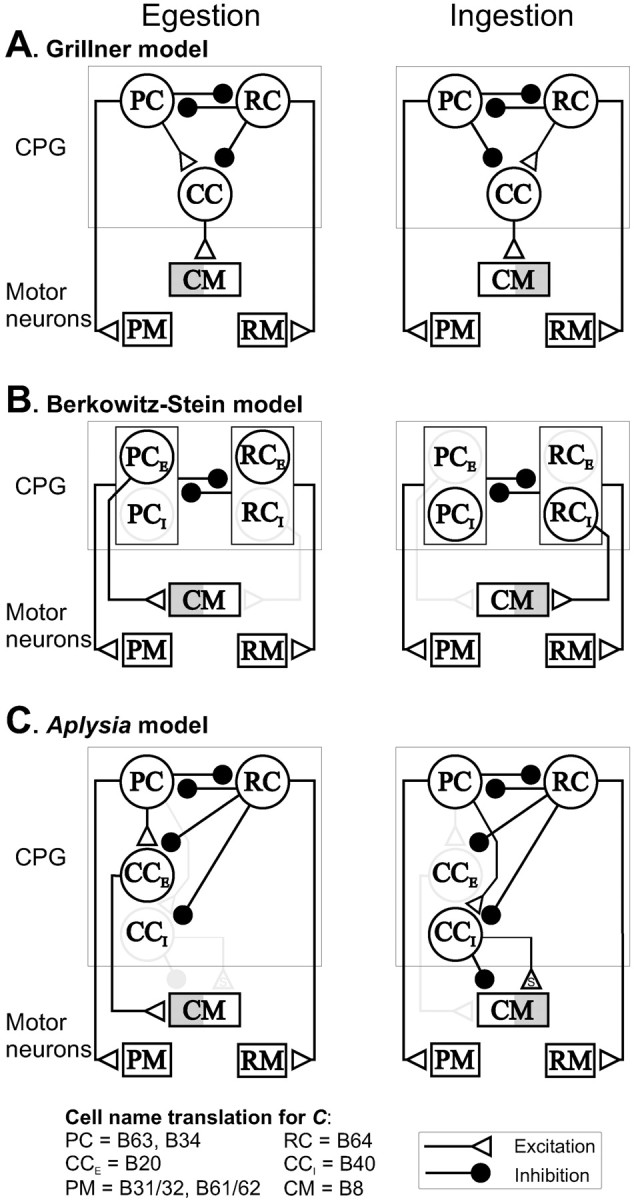
Alternative network models of the generation of egestive and ingestive motor programs in Aplysiafeeding. The two motor programs require coordination of two sets of radula movements, protraction–retraction and opening–closing. Each rhythmic cycle consists of alternating protraction and retraction phases that are represented by protraction motor neurons (PM) and retraction motor neurons (RM), respectively, in the diagrams. In each cycle, protraction precedes retraction, whereas the radula closure shifts its phase relative to protraction–retraction. Thus, in egestive motor programs (left panels), the radula closure motor neuron (CM, B8) is active in protraction (depicted as gray shades in the left half of CM), whereas in ingestive motor programs (right panels), CM is active in retraction (depicted as gray shades in theright half of CM). The phasing of motorneuronal activity is mediated by interneurons of a central pattern generator (CPG). PM and RM are controlled by protraction controller interneurons (PC, B63, and B34) and retraction controller interneuron (RC) (e.g., B64), respectively. PC and RC reciprocally inhibit each other (Hurwitz et al., 1997), thus forming a half-center oscillator. A, A model based on the Grillner (1981,1985) hypothesis in spinal circuit (Grillner, 1981, 1985). The main feature of the model is that interneurons (i.e., controllers) for radula movements [PC, RC, as well as closure controller interneurons (CC)] are shared between ingestion and egestion. Phase shifting of CM between protraction and retraction in the two programs is mediated by phase shifting of CC through a change of sign of coupling that is implemented by descending fibers. Specifically, in egestion, CC is driven by PC and inhibited by RC, whereas in ingestion, CC is driven by RC and inhibited by PC. B, A model based on the Berkowitz and Stein (1994) model of the turtle-scratching spinal circuit. In this model, protraction and retraction are mediated by partially overlapping sets of PC and RC interneurons. In egestion, PCs, which are maximally tuned to egestion (PCE), excite both PM and CM, thus CM is active together with PM. Similarly in ingestion, RCs, which are maximally tuned to ingestion (RCI), excite both RM and CM, thus B8 is active together with RM. C, TheAplysia feeding network model that is based on present and earlier studies on feeding interneurons (Hurwitz and Susswein, 1996; Hurwitz et al., 1997; Jing et al., 1999; Jing and Weiss, 2001). Translation of cell names that are mentioned in the paper to functional terms (PC, RC, CCE, CCI, PM, CM) is provided at the bottom. Similar to the Grillner (1981, 1985) model, PC and RC remain the same in both egestion and ingestion. On the other hand, similar to theBerkowitz and Stein (1994) model, two separate CCs (CCE for egestion and CCI for ingestion) are recruited to mediate CM phasing in the two motor programs. In egestion, CCE (B20) receives excitation from PC and inhibition from RC, so it is active in protraction. CCE drives CM directly through fast excitation so CM is active in protraction. In addition, a separate neuron B4/5, which is preferentially active in egestion and active in retraction, inhibits B8 to prevent B8 firing in retraction (Jing and Weiss, 2001) (not depicted for clarity). In ingestion, CCI (B40) also receives excitation from PC and inhibition from RC, so it is active in protraction. However, unlike CCE, CCIdoes not use fast excitation; instead, it uses slow excitation to promote CM activity in retraction. In addition, CCI exerts fast inhibition on CM to precisely activate CM at the onset of retraction and prevent excessive B8 activity in protraction that may be promoted by CCE (see Discussion). An swithin the connection symbols (C) depicts “slow synaptic connections.” Functionally ineffective synaptic connections and weakly active neurons are shown ingray.