

Abstract
Fibronectin is a multifunctional, extracellular matrix glycoprotein that exists either as an insoluble multimeric fibrillar component of the extracellular matrix or as a soluble monomer. Cells attach to fibronectin through transmembrane integrin receptors and form a variety of cell-matrix contacts. Here we show that primary fibroblasts can use fibronectin to organize a specific cell-cell contact – “stitch adhesions.” This contact is formed by short parallel fibronectin fibrils connecting adjacent cells above the level of the focal adhesions that attach the cells to the substrate. Stitch adhesions contain integrin α5β1 but not αVβ3, align with actin filament bundles, and contain talin, tensin, α-actinin, vinculin, paxillin and a phosphorylated form of focal adhesion kinase. This combination of components differs from the described constituents of the known cell adhesions. Stitch adhesions are organized when protein synthesis and secretion are inhibited by cycloheximide and exogenous fibronectin is provided to the cells. The adhesion stitches described here provide an attractive model system for studying fibronectin fibrillogenesis and the mechanisms governing the formation of cellular adhesions.
Keywords: fibronectin, cell-cell contacts, adhesion, cycloheximide, human fibroblasts
1. Introduction
Fibronectin (FN) is a multifunctional glycoprotein of the extracellular matrix (ECM) with crucial roles in many physiological and pathological processes. The molecule is composed of two nearly identical subunits, linked covalently near their C-termini by a pair of disulfide bonds. Although it is the product of a single gene, FN exists in several isoforms that arise from alternative splicing [1]. The FN molecule is built on a modular principle, and each of its subunits contains 12 type I repeats, two type II repeats and 15-17 type III repeats, which together account for approximately 90% of the FN sequence [2,3]. FN is widely expressed by multiple cell types and throughout all stages of life. It is essential for vertebrate development, as demonstrated by the early embryonic lethality of mice with FN gene disruption [4].
A distinctive feature of fibronectin is its ability to bind to a large number of molecules, including cell surface receptors, growth factors and components of the extracellular matrix. FN is a ligand for a dozen members of the integrin family – transmembrane heterodimeric receptors that link the ECM with the intracellular cytoskeleton [5,6]. The major receptor for fibronectin is α5β1 integrin, which binds to the RGD sequence in the 10th type III repeat [7] and the synergy site in the adjacent 9th type III repeat of the FN molecule [8]. The same receptor is the primary player in the conversion of soluble, unorganized FN into linear or branched interconnected meshwork of fibrils around cells – a process known as fibronectin fibrillogenesis [9]. The formation of FN fibrils depends on actin-driven centripetal translocation of α5β1 integrins out of focal adhesions towards the cell body. Using these adhesions as an anchor, moving integrins unfold fibronectin that is bound to the cell surface, and form a different type of cell–matrix adhesion, the fibrillar adhesion [10,11]. The stretching forces applied to FN lead to exposure of cryptic fibronectin self-assembly sites, thus promoting self-association and further polymerization of FN fibrils [12,13,14]. Ongoing fibrillogenesis leads to accumulation of a thick, three-dimensional (3D) fibronectin meshwork. Cells, embedded in this ECM lose their contacts to the artificial flat and rigid surface of the tissue culture dish and adapt to the three-dimensional environment by substituting 3D matrix adhesions for focal and fibrillar adhesions [15,16]. This generally accepted notion implicates fibronectin and integrins as major players in establishment of the cell-substratum adhesions in two- as well as in three-dimensional environments.
Nevertheless, under specific experimental settings, integrins have been associated with the development of cell-cell types of contacts [17,18]. The integrin ligand – fibronectin – has also been observed to connect adjacent cells. The initial reports came from the Hynes lab, demonstrating that normal and FN-deficient HSV-transformed NIL cells, grown in low serum, organize fibronectin (LETS protein) as “stitches” of variable size, connecting adjacent cells [19,20]. Using the same HSV-transformed cells, Singer described an association between extracellular fibronectin and intracytoplasmic actin, which he termed the fibronexus [21]. Unlike the more robust ventral arrangement of fibronectin and integrins in a focal adhesion that contains vinculin, the more delicate vinculin-negative fibronexus is detected on the dorsal cell surface in the form of fibronectin stitches at sites of intercellular contact [22,23]. An interesting observation connects the formation of fibronectin stitches to the absence of organized collagen in the extracellular space. Similar to transformed fibroblasts, chondrocytes do not deposit collagen fibers, and they organize FN as short intercellular strands, while other cells that organize collagen form extensive extracellular fibronectin networks [24]. In support of this notion, Dzamba demonstrated that Movl3 fibroblasts, which do not express endogenous α1(I) collagen chains due to a retroviral insertion, produce a sparse matrix containing short fibronectin fibrils localized between cells [25]. This deficiency can be rescued by the addition of collagen I, suggesting that elongation of FN fibrils is dependent on the presence of this collagen type in this system.
Consequently, the formation of fibronectin-mediated stitch-like fibrillar connections between cells appears to be a frequently observed phenomenon. However, the molecular nature of the cell-matrix adhesion associated with these cell-cell spanning matrix connections has not been established nor compared to known cell-matrix adhesions. Here, we identify specific experimental conditions for reproducible induction of this class of “stitch adhesions,” which allowed us to characterize their morphology, dynamics of formation and main constituents. Evaluation of their properties indicates that they differ from previously described cell-cell and cell-matrix connections and can be regarded as a separate type of cell adhesion.
2. Experimental procedures
2.1. Antibodies and proteins
The following antibodies were used: antibody against human β1 integrin, isotype antibody control IgG2a,κ, anti-FN, anti-tensin (all from BD Biosciences), anti-human integrin-α5 nonfunction-blocking mAb11 [26], anti-human integrin-α5 inhibitory mAb16 [27], anti-human integrin-β1 inhibitory mAb13 [28], anti-human integrin-αV L230 (ATCC), anti-human FN mAb 13G12 [28], anti-β3 integrin (sc-7311; Santa Cruz Biotechnologies), anti-phospho-FAK (Fischer Scientific). Other antibodies used were from Sigma-Aldrich: anti-actin, anti-talin, anti-α-actinin, anti-vinculin and anti-paxillin. Secondary species–specific FITC-, Cy3- or AMCA-conjugated antibodies were from Jackson ImmunoResearch Laboratories. Rhodamine-phalloidin was from Fischer Scientific.
Human plasma FN was purified according to Miekka [29] and 70kD fibronectin fragment was obtained as described [30]. The 120kD fragment from plasma FN was purchased from Merck. Fibronectin-depleted FBS was obtained by the use of Gelatin-Sepharose 4B (LKB) as described by Knox [31]. Cellular fibronectin was purified according to Yamada et al. [32]. HiLyte Fluor™ 488 labeled bovine FN was purchased from Cytoskeleton Inc.
2.2. Cells, cycloheximide treatment and microscopy
Primary human foreskin fibroblasts (HFF) were a gift from Susan Yamada (NIDCR, NIH) and were cultured as previously described [10].
For preparation of cycloheximide (CHX)-treated samples, cells grown to 80% confluence were trypsinized, washed with trypsin inhibitor, washed once with CHX medium (DMEM, containing 1% FN-depleted FBS, penicillin (100 U/ml), streptomycin (100 μg/ml) and 25 μg/ml cycloheximide) and incubated as a suspension in the same medium for 30 min at 37°C with rotation. Cell suspensions were plated on coverslips or culture plates that had been pretreated for half an hour with the same medium. After overnight incubation in CHX to deplete intracellular fibronectin, cells were treated with 25 μg/ml full-length plasma or cellular fibronectin or the same amount of 70kD and 120kD FN fragments for four hours. When function-disrupting anti-integrin antibodies were used, the cells were pretreated with 200 μg/ml of each antibody for 30 min, followed by 25 μg/ml HiLyte Fluor™ 488 labeled bovine FN in the presence of the same concentrations of antibodies for four hours. The formation of cell-to-cell contacts was monitored with Axiovert 25 microscopes (Carl Zeiss MicroImaging, Inc.) using 5x NA 0.12 or 10x NA 0.25 A-Plan objectives. Images were collected with CCD video camera at 15-min intervals and digitized.
2.3. Pulse-chase formation of stitch adhesions with labeled fibronectin
Plasma fibronectin labeled with Alexa 488 and Alexa 594 were obtained by using Alexa Fluorophore Labeling kits (Thermo Fisher Scientific) according to the manufacturer’s recommendations.
Coverslips with CHX-treated fibroblasts were incubated with 25 μg/ml human FN-488 in CHX medium for one hour, washed quickly two times with warm PBS and incubated for an additional hour with 25 μg/ml FN-594 in the sane medium. At the end of the incubation, the coverslips were washed, fixed and stained with 5 μg/ml Hoechst 33342 (Thermo Fisher Scientific) in PBS for 5 min before mounting.
2.4. Immunofluorescence
Cells for immunofluorescence were fixed in 4% paraformaldehyde in PBS containing 5% sucrose for 15 min and permeabilized with 0.5% Triton X-100 in PBS for 4 min. Immunostaining was performed with the indicated antibodies at room temperature for 1 h, followed by appropriate secondary FITC-, CY3- or AMCA-conjugated antibodies. Stained samples were mounted in Fluoromount™ Aqueous Mounting Medium (Merck). Immunofluorescence microscopy images were obtained using a Leica confocal microscope (Leica TCS SPE) or a Zeiss Axiophot microscope equipped with a Photometrix CH 350 cooled CCD camera and fluorescence optics.
2.5. Coimmunoprecipitation and Western Blotting
Control and FN-incubated CHX-treated fibroblasts were solubilized on ice in RIPA buffer (150 mM NaCl, 2 mM EDTA, 1% sodium deoxycholate, 0.1% SDS, 1% Triton X-100, 10% glycerol, 50 mM HEPES, pH 7.5) containing Complete Inhibitory Cocktail (Merck). Homogenates were cleared of cellular debris by centrifugation at 20,000 g for 15 min at 4°C. Immunoprecipitates using anti-β1 integrin antibodies, isotype antibody control and Gammabind Plus Sepharose (Merck) were resolved on 8% gels. After electrotransfer to UltraCruz nitrocellulose membranes (Santa Cruz Biotechnology), the filters were blocked (5% non-fat dry milk in TBST: 150 mM NaCl, 50 mM Tris HCl, 0.1% Tween 20, pH 7.4) for 1 h. Membranes were than incubated with appropriate primary and HRP-conjugated secondary antibodies (Enzo Life Sciences). Immunoblots were visualized using Western Blotting Luminol Reagent (Santa Cruz).
2.6. Transfection
Full-length human paxillin cDNA and full-length chicken tensin cDNA, ligated into the GFP expression vector pGZ21, were provided by Kazue Matsumoto (NIDCR, NIH). Subconfluent cultures of fibroblasts were trypsinized and suspended in electroporation buffer (20 mM HEPES, 137 mM NaCl, 5 mM KCl, 0.7 mM Na2HPO4 and 6 mM dextrose, pH 7.05). GFP-paxillin or GFP-tensin vectors (30 μg) were added to 1.5×106 cells, and electroporated at 170 V and 960 mF using a Bio-Rad Gene Pulser. Following electroporation, cells were collected by centrifugation, resuspended in culture medium containing 5 mM sodium butyrate to enhance expression of transfected genes, and plated.
2.7. Statistics
The average and standard deviation (SD) of immunofluorescence images and analysis by one way ANOVA were calculated using GraphPad Instat. Probability (P) values were considered significant if <0.05.
3. Results
3.1. Fibronectin induces cell-cell contacts in cycloheximide-treated fibroblasts via α5β1 integrin
Fibroblasts synthesize and secrete a large number of proteins [33] including the components of extracellular matrix (ECM). To study the conversion of soluble fibronectin into fibrils in a controlled system, particularly in the absence of endogenous fibronectin and other secreted proteins of the ECM, we treated human foreskin fibroblasts with cycloheximide (CHX). CHX is a eukaryotic-specific inhibitor of protein synthesis that suppreses translation elongation [34]. Incubation of trypsinized HFF (human foreskin fibroblasts) in suspension for 30 min, followed by plating and overnight treatment with 25 μg/ml CHX (see section 4.2) effectively depleted all endogenous FN from the culture as demonstrated by immunofluorescence and Western blotting (see below). Under these conditions, fibroblasts preserved their viability but acquired an elongated morphology (Fig. 1A), slowed their migration and were often observed to move smoothly over the top of one another (Fig. 1A, inset). Addition of 25 μg/ml purified human plasma, or cellular fibronectin, induced marked changes in their behavior and morphology. Within several hours, cells organized extensive cell-cell contacts, became flattened and formed epithelia-like connected islands (Fig. 1B), and given enough time (depending on the plating cell density), all single cells joined the islands. This process was most pronounced during the first hour after addition of FN when most of the touching cells developed contacts (Fig.1C, 0’ to 60’). During the next several hours, the formation of cell-cell contacts gradually slowed, and the cells became well-flattened (Fig. 1C, 75’ to 180’).
Figure 1.

Full-length fibronectin induction of formation of cell-to-cell contacts in cycloheximide-treated fibroblasts. Phase-contrast images of CHX-treated cells revealed elongated morphologies in the absence of FN (A), but after incubation with 25 μg/ml human plasma FN for 4 hours (B), they developed extended cell-cell contacts, combined with an epithelial-like morphology. Time-lapse images acquired at 15 min intervals demonstrated quick initial establishment of cell-to-cell contacts that gradually increased (C). Arrows indicate developing cell-cell contacts. Bar = 50μm.
Immunofluorescence analysis with anti-fibronectin antibody demonstrated the absence of secreted endogenous FN after CHX treatment (Fig. 2A) and specific localization of exogenously added FN at the sites where cells had developed cell-cell contacts (Fig. 2B). Short FN fibrils, organized as “stitches,” connected adjacent cells and were absent from the free cell edges. Formation of these “stitch adhesions” (StA) was characteristic of full-length fibronectin, since repeating the incubation with 70 kD or 120 kD fragments of the FN molecule did not induce formation of cell-cell connections. Although both fragments contain integrin binding sites and encompass regions involved in interactions with other parts of the FN molecule that are important for fibrillogenesis [35], no organized fibronectin was detected in these samples (Fig. 2C and D). Instead, the staining for fibronectin was diffuse and localized to the areas of coverslips free of cells, most probably representing FN fragments attaching directly to the substrate (Fig. 2C and D, insets).
Figure 2.

Immunofluorescence of CHX-treated fibroblasts incubated with full length FN or fragments of FN and stained for fibronectin (green) and actin (red). (A) The control CHX-treated cells did not stain with anti-fibronectin antibodies, indicating that synthesis and secretion of FN were completely blocked. (B) Prominent fibronectin stitches were observed between adjacent cells after incubation with full-length plasma FN. No organized FN was detected in samples incubated for the same period of time with 70kD (C) or 120kD (D) fibronectin fragments. Nuclei (blue) were stained with Hoechst. Insets present only fibronectin staining. Arrowheads in B indicate free cellular edges devoid of FN. Bar = 50μm.
To gain further insight into the dynamics of formation of the stitch adhesions, we conducted pulse-chase experiments with fibronectin labeled with different fluorochromes (Fig. 3A). This technique permits determination of the direction of fiber growth as indicated by the relative distribution of the two fluorescent signals [35]. Incubation of CHX-treated HFF with fibronectin labeled with green Alexa 488 for one hour, followed by additional hour with red Alexa 594, revealed that fibers formed during the first hour of incubation (green stitches in Fig. 3A, FN-488) did not extend considerably afterwards. The length of the fibers changed only from 8 to about 9.5 μm during the subsequent hours (in Fig. 3B, compare 1 hour to 2 and 3 hours). Pulse-chase experiments revealed that the red FN after 1 hour did not add substantial red-labeled extensions, but instead associated with preexisting green fibers, so that they became thicker and yellow in appearance (indicating co-localization, Fig. 3A, FN-488/FN-594). New stitches organized during this second hour of incubation were also detected. They were easily distinguished by their red color and absence during the first hour of incubation with FN (Fig. 3A, arrows). Notably, after incubation with red FN, the samples contained some stitches that were only green-labeled, indicating that they had ceased their FN polymerization.
Figure 3.

Dynamics of FN stitch development. (A) Pulse-chase analysis of the formation of stitch adhesions using labeled fibronectin preparations. CHX-treated fibroblasts were incubated with fibronectin labeled with green Alexa 488 (FN-488) for 1 hour, followed by another hour of incubation with fibronectin labeled with red Alexa 594 (FN-488/FN-594). Arrows indicate stitches formed during the second hour of incubation. Merged fluorescence and phase contrast images show localization of labeled stitches at places of cell-cell contacts. Nuclei were visualized with Hoechst (blue). Bar = 20μm. (B) Determination of stitch length. CHX-treated fibroblast were incubated with fibronectin for different periods, fixed and stained with anti-fibronectin antibodies. The lengths of FN stitches were measured on immunofluorescence images and presented as box plots.
Although fibronectin can serve as a ligand for many integrins [6], the major receptor responsible for binding and reorganization of FN in fibroblasts is α5β1. To test directly the role of this integrin heterodimer in the formation of stitch adhesions, we used specific anti-α5 and anti-β1 antibodies previously shown to interfere with their function. CHX-treated fibroblasts were incubated with inhibitory anti-α5 antibody (mAb16), inhibitory anti-β1antibody (mAb13), nonfunction-blocking anti-α5 antibody (mAb11) or isotype antibody control (IgG) before induction of StA formation by the addition of FN. The same antibodies were kept present in the medium during the organization of fibronectin stitches after FN addition (Fig. 4). Non-inhibitory mAb11 as well as control isotype antibody did not inhibit the development of StA. The fibronectin stitches organized in these samples were similar to the stitches formed by fibroblasts in the control samples (Fig. 4, compare control with IgG and mAb11). In contrast, blocking normal function of either the α5 integrin subunit with mAb16 or of integrin β1 with mAb13 completely prevented FN organization. The observed immunofluorescence results were consistent with the changes in cell morphology. The cells in samples with organized StA were clustered into epithelia-like islands, while treatment with inhibitory antibodies prevented the formation of cell-to-cell contacts and preserved the typical elongated fibroblast morphology, even though fibronectin was present in the medium (Fig. 4, phase). These results demonstrated that α5β1 integrin was the primary receptor responsible for the organization of StA.
Figure 4.

Blocking integrin α5β1 prevents the formation of StA and cell-to-cell contacts. Cycloheximide-treated human foreskin fibroblasts were incubated for 30 minutes with 200 μg/ml IgG2a,κ isotype antibody control (IgG), nonfunction-blocking anti-α5 antibody (mAb11), inhibitory anti-α5 antibody (mAb16), and inhibitory anti-β1antibody (mAb13) or were left untreated (control). Stitch adhesions were induced in all samples by incubation with 25 μg/ml HiLyte Fluor™ 488-labeled bovine FN in presence of the same concentrations of antibodies for an additional 4 hours. Fibronectin was observed using the green fluorescence channel (FN), rhodamine-phalloidin was used for visualization of the actin cytoskeleton (red channel), and nuclei were stained with Hoechst (blue channel). Merged images with the corresponding colors are shown under “FN/actin/nuclei”. General morphology of the cells was determined using phase contrast microscopy (phase). Bar = 50μm.
3.2. Fibronectin stitches align with actin bundles and contain major adhesome proteins
It has been extensively documented that fibronectin polymerization requires a well-developed and functionally active actin cytoskeleton, capable of generating sufficient tension for unfolding the FN molecule and exposing cryptic self-association sites. Morphologically, this relationship is indicated by alignment between fibronectin fibrils and underlying bundles of actin filaments beneath the cell membrane. Thorough examination of samples stained for fibronectin and actin demonstrated a similar pattern of distribution for stitch adhesions. The fibronectin fibrils from the stitches aligned with clearly distinguishable actin bundles (Fig. 5A arrows). In some cases, the fibrils even bridged the small gaps between adjacent cells (Fig. 5A arrowheads). These results prompted us to search for molecular constituents that could provide a physical link between FN stitches and the actin cytoskeleton.
Figure 5.

Adhesion stitches aligned with actin filament bundles and associated with proteins typical of the integrin adhesome. Immunofluorescence images of (A) CHX-treated fibroblasts incubated with 25 μg/ml human plasma FN for 4 hours and then stained with anti-fibronectin antibodies (FN) and rhodamine-phalloidin (actin). Merged images (overlay) demonstrated alignment between actin bundles and fibronectin stitches (arrows). Fibronectin fibrils occasionally bridged the gap between neighboring cells (arrowheads). Bar = 20μm. Co-immunoprecipitation experiments of (B) CHX-treated cells in the absence (−FN) or presence (+FN) of fibronectin performed with anti-β1 integrin antibody 9EG7 (IP:9EG7) and rat IgG2a,κ isotype antibody control (IP:IgG), followed by Western blotting (WB) with antibodies against the indicated proteins revealed strong enrichment of adhesome proteins in β1 integrin complexes after incubation of cells with FN. Loading controls prior to immunoprecipitation (Lysate) are also shown.
In all types of contacts between cells and extracellular matrix proteins, a central role is played by transmembrane integrins [36]. Inside cells, they anchor bundles of actin microfilaments through an adhesion plaque containing the integrin adhesome, which consists of many different proteins [37,38]. Since α5β1 is the major integrin involved in FN polymerization, we performed co-immunoprecipitation experiments with 9EG7 antibody, which recognizes the activated form of α5β1. When immunoprecipitates were prepared from CHX-treated HFF grown in the absence of FN, there were only traces of actin and the adhesion plaque proteins we examined (Fig. 4B, −FN). Incubation of the cells with FN, which induced the formation of stitch adhesions, led to the appearance of prominent bands of actin, talin, tensin, α-actinin, vinculin, paxillin and FN as coimmunoprecipitating proteins (Fig. 5B, +FN). These findings were specific, since immunoprecipitation with isotype antibody control produced only a faint actin band (Fig. 5B, IgG). Because the 9EG7 antibody recognizes activated β1 integrin subunits, which can heterodimerize with a variety of different α subunits, we cannot entirely exclude the possibility that the immunoprecipitates might contain minor contaminants associated with other than α5β1 heterodimers. For example, αVβ1 that binds osteopontin might have been activated, since 1% FN-depleted serum is present in our experimental system. In fact, such a possibility may explain the faint bands of adhesome proteins detected in the immunoprecipitated samples that are not incubated with fibronectin (Fig. 5B, −FN).
To further clarify the participation of specific integrins and adhesome proteins in stitch adhesions, we next performed extensive immunofluorescence studies (Fig. 6). Staining with antibodies against the integrin subunits α5 and β1 clearly indicated their localization with fibrils of the stitches (Fig. 6, arrows on panels α5 and β1). The fluorescence signal from the stitches was noticeably brighter than staining of the focal adhesions (Fig. 6, arrowheads on panels β1 and α5), probably due to the absence of fibronectin coated on the substrate. Unlike α5β1, αV and β3 integrin subunits were detected in focal adhesions and were completely absent from the StA (Fig. 6, panels β3 and αV), indicating that αVβ3 and αVβ1 integrin heterodimers did not participate directly in their formation.
Figure 6.

Identification of molecular components of the stitch adhesions. Cycloheximide-treated human foreskin fibroblasts were incubated with fibronectin and stained with antibodies against the indicated integrins, adhesome proteins and FN. The primary antibodies were visualized with secondary antibodies conjugated to FITC (green channel) or Cy3 (red channel). Merged images are presented in the righthand column (overlay). Arrows indicate stitch adhesions, and arrowheads focal adhesions. Bar = 20μm.
Vinculin, an adhesome protein involved in linkage of integrin molecules to the actin cytoskeleton, and the activated form of focal adhesion kinase (p-FAK) – one of the major kinases operating at cell adhesion sites, were also observed in the stitch adhesions (Fig. 6, panels Vinc and p-FAK). The intensity of staining for both proteins was not uniform along FN fibrils, and some of the stitches were more strongly stained than others, suggesting the dynamic nature of this localization. Judging by the plane of focus during microscopy, StA were positioned above the plane of substrate-attached focal adhesions, indicating their dorsal localization.
Further experiments with cells transiently transfected with GFP-labeled tensin and paxillin allowed simultaneous visualization of these proteins together with two additional antigens of interest (Fig. 7). A combination between GFP-tensin and antibodies against FN and integrin αV confirmed the presence of tensin in stitch adhesions as indicated initially by co-immunoprecipitation experiments (Fig. 7, left column). The images in this column represent two cells situated with one on top of the other. From the distribution of focal adhesions (Fig. 7, αV panel), which are the places of closest contacts between cell and substrate, and the phase contrast image, it appears that the outlined cell is the most ventral cell. This unusual placement of cells demonstrated again the dorsal localization of StA and indicated that they could be formed not only between adjacent cells but also between cells positioned with one on top of the other.
Figure 7.
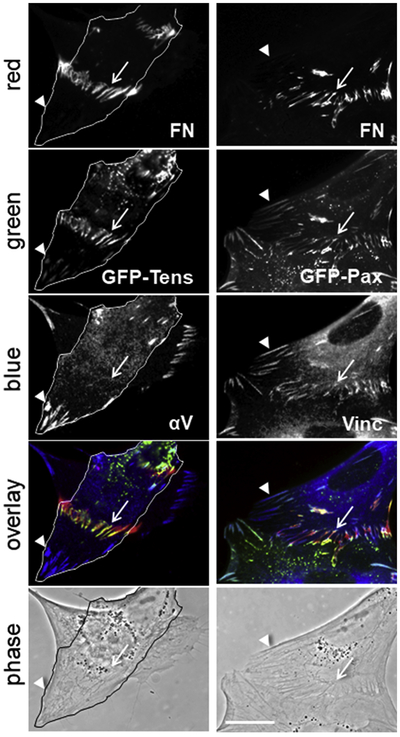
GFP-tagged tensin and paxillin localize at stitch adhesions. Transiently transfected HFF with GFP-tensin (left column) and GFP-paxillin (right column) were stained with antibodies against fibronectin (FN), αV integrin (αV) and vinculin (Vinc). The primary antibodies were visualized with secondary antibodies conjugated to Cy3 (red) or AMCA (blue). GFP fluorescence was detected on the green channel (green). Merged images are presented under “overlay” and phase contrast images under “phase”. Arrows indicate stitch adhesions and arrowheads - focal adhesions. The cell positioned closer to the substrate in the images in the left column is outlined. Bar = 20μm.
GFP-paxillin also localized at fibronectin fibrils connecting adjacent cells and, as expected, was found at focal adhesions (Fig. 7, arrowheads on the right column). Merged images confirmed previous observations of the nonuniform distribution of adhesome proteins along fibronectin fibrils. Green and blue colors, corresponding to paxillin and vinculin, respectively, were concentrated at the edges of the stitches, leaving the middle to appear as only red from the signal for anti-fibronectin antibodies (Fig. 7, overlay in the right column).
3.3. Removal of cycloheximide restores normal fibronectin fibrillogenesis.
The unusual organization of FN and formation of stitch adhesions we observed resulted from the absence of synthesis and secretion of proteins, especially fibronectin, due to the cycloheximide treatment. We were interested to clarify whether this treatment had long-lasting effects on the ability of treated cells to handle fibronectin, or whether removal of the drug would restore normal fibrillogenesis. To answer these questions, we performed experiments where different combinations of CHX treatment and addition of labeled bovine FN were used (Fig. 8A, upper panels). Since the effect of CHX on translation is reversible [39], all samples were allowed to restore (CHX-treated) or continue (non CHX-treated) their normal secretion and organization of endogenous FN overnight by incubation in media supplemented with 1% FN-depleted serum only (Fig. 8A, lower panels). The endogenous human FN was distinguished from the exogenously added green bovine FN by staining with anti-human FN-specific antibody and secondary antibody conjugated with a red fluorophore. The results indicated that when cycloheximide was not present, exogenous bovine FN was successfully incorporated into the FN fibrils organized by the cells using endogenous human FN. Hybrid FN fibrils containing both fluorochromes were orange or yellow in appearance (Fig. 8A, −CHX +bFN, arrowheads) and could be easily distinguished after overnight incubation, although the matrix assembled by endogenous FN was more pronounced. When cycloheximide was present, endogenous human FN could not be detected, and the exogenous bovine FN assembled into typical StA (Fig. 8A, +CHX, upper panels). Comparisons between FN fibrils in StA and fibrils organized in the absence of CHX revealed noticeable differences. In the samples with normal or restored fibrillogenesis, the FN fibrils were situated predominantly on the cell bodies, while the stitches were localized exclusively between cells. In addition, individual fibrils in stitch adhesions were mostly uniform in length and thicker than the fibrils organized by non-treated cells or after removal of CHX. Washing away cycloheximide restored normal fibrillogenesis (Fig. 8A, compare +CHX −bFN lower panel with −CHX −bFN) and led to the disappearance of StA. Green FN fibrils from the stitches were reorganized and embedded into the developing FN matrix formed by secreted human FN (Fig. 8A, +CHX +bFN, lower panel, arrowheads). Occasionally, contracted green fibronectin fibers originating from StA were also detected (Fig. 8A +CHX +bFN, lower panel, arrow). These results demonstrated that after removal of CHX, cells successfully resumed deposition of fibronectin matrix. Moreover, the general morphology of the five days-old matrix (Fig. 8B +CHX) was similar to the matrix organized by HFF that had not been subjected to any treatments (Fig. 8B −CHX), suggesting that CHX did not induce long-lasting changes in fibronectin fibrillogenesis.
Figure 8.

Removal of cycloheximide restores normal fibronectin fibrillogenesis. (A) Fibroblasts treated (+CHX) or not treated (−CHX) with cycloheximide overnight in media containing 1% FN-depleted serum were incubated (+bFN) or not (−bFN) with 25 μg/ml HiLyte Fluor™ 488-labeled bovine FN for 4 hours. All samples were washed and left overnight in the above medium in the absence of both cycloheximide and labeled FN (O.N. in medium −CHX −bFN). To visualize the endogenously produced human fibronectin, samples were stained with anti-human FN antibody and secondary Cy3-conjugated antibody. The green channel was used to detect bovine FN (bFN), the red channel for human FN (hFN), and the blue channel for Hoechst-stained nuclei (Hoechst). Arrowheads indicate hybrid FN fibers containing both bovine and human fibronectin; arrows indicate FN fibers originating from StA. (B) Fibroblasts treated (+CHX) or not treated (−CHX) with cycloheximide as above were grown in media containing 1% FN-depleted serum for an additional 5 days. Fibronectin matrix was stained with anti-human FN antibody (red) and nuclei were stained with Hoechst (blue). Bar = 50μm.
To gain further insight into the process of stitch reorganizations after restoration of normal FN fibrillogenesis, we used CHX-treated fibroblasts connected with stitch adhesions formed by green-labeled bovine FN and studied them shortly after a release from cycloheximide block (Fig. 9). Two hours after removal of CHX, StA became slightly disorganized and some of their FN fibrils were released from the cells, as judged by the loss of their stretched fibrillar appearance (Fig. 9, 2 h, arrows). At the same time, secreted human FN became detectable, demonstrating faint accumulation at StA (Fig. 9, 2 h, red channel, arrowheads) or appearance at StA-free cell areas (Fig. 9, 2 h, red staining, overlay). At four hours, the disintegration of StA was almost complete, leaving green bovine FN either embedded into newly formed bovine/human FN fibrils (Fig. 9, 4 h, arrowheads) or as small non-fibrillar aggregates (Fig. 9, 4 h, arrows). A similar distribution of the bovine FN was detected after 24 hours, although at this late time point the matrix was comprised predominantly of human FN (Fig. 9, 24 h). Our results suggested that the presence of StA did not influence the process of normal FN fibrillogenesis, nor the morphology of the assembled FN matrix. In addition, they imply that normal fibronectin fibrillogenesis depends on protein synthesis and secretion and occurs in a different manner when these processes are blocked.
Figure 9.

Reorganization of stitch adhesions after restoration of normal fibronectin fibrillogenesis. Fibroblasts with organized StA (0 h) were washed and incubated in CHX-free DMEM containing 1% FN-depleted serum for the indicated times. Stitch adhesions formed by HiLyte Fluor™ 488-labeled bovine FN were detected in the green channel and the secreted human FN was observed in the red channel after staining with anti-human FN antibody and secondary Cy3-conjugated antibody. Nuclei were stained with Hoechst. Merged fluorescent images are presented under “overlay”. Arrowheads indicate hybrid FN fibers containing bovine and human fibronectin, and arrows indicate – FN fibers originating from StA. Bar = 50μm.
4. Discussion
Fibroblasts are a type of connective tissue cells that play important roles in secretion and organization of ECM, as well as in the proliferative and remodeling phases of wound healing. They connect to the substrate through a well-developed system of adhesions [16,40,41], which has made them a widely used model to study cell-extracellular matrix types of adhesions. Unlike epithelial cells, their cell-to-cell contacts are much less, and when induced artificially, can even lead to non-apoptotic programmed cell death [42]. Here we describe results demonstrating that primary human foreskin fibroblasts can organize extensive cell-cell contacts by using integrin α5β1 and fibronectin in stitch adhesions (Fig. 10). Development of these stitch contacts in this model system was induced under specific conditions requiring two important steps. First, cells were grown under conditions inhibiting protein synthesis and secretion. This was achieved by treatment with cycloheximide, which effectively inhibited the production of endogenous FN. Second, addition of exogenous full-length FN was necessary for induction of cell-cell contacts. The added FN engaged in polymerization and formed characteristic fibrils connecting adjacent cells, which we named “stitch adhesions” because of their specific appearance.
Figure 10.

Schematic representation of the stitch adhesion. Unlike focal adhesions (FA), which are located at the ventral cell surface and attach cells to the substrate mainly through the vitronectin receptor αVβ3, stitch adhesions are positioned dorsally and connect adjacent cells by short fibronectin fibrils bound exclusively to α5β1 integrins. Both types of adhesions connect to the actin cytoskeleton through protein plaques composed of similar members of the integrin adhesome.
The morphology of the fibrils of the stitches differs significantly from the fibrils organized by fibroblasts with active protein synthesis and secretion (Fig. 8). They were short, with a similar length of approximately 9.5 μm, thick and parallel to each other. The stitches aligned with actin bundles, they stained for both subunits of integrin α5β1 (Fig. 6), and their formation was strongly dependent on the functional activity of this integrin heterodimer (Fig. 4). A number of integrin adhesome proteins co-localized with the stitches, although their distribution along fibrils were not equal and often concentrated at the edges of the fibrils (Fig. 6 and 7). In particular, the stitch adhesions were positive for the major adhesion plaque proteins responsible for linking integrins to actin cytoskeleton, namely α-actinin, talin, vinculin and tensin [38]. This suggests that, analogous to the other adhesions, cell-generated tension could be transmitted to integrins and bound fibronectin molecules. Another common member of focal adhesions that was identified in adhesion stitches was focal adhesion kinase. FAK is a protein tyrosine kinase with important role in development, cell migration, cell proliferation and tumorigenesis [43]. Moreover, we identified the activated/phosphorylated form of FAK (Fig. 6, p-FAK), implying that stitch adhesions may be involved in cell signaling, similar to focal adhesions. Comparing the characteristics of the already described types of adhesions with stitch adhesions (StA), it became evident that they share many similarities but did not match completely with any of them (Table 1). Focal adhesions contain αVβ3 integrin, which was missing from stitch adhesions (Fig. 6). While fibronectin and α5 integrin are detected only at the medial periphery of focal adhesions [15], the immunofluorescence signal from α5 was strong and covered the entire length of the FN fibrils of the stitches (Fig. 6). Unlike fibrillar adhesions, StA were positive for paxillin and vinculin in both immunoprecipitation (Fig. 5B) and immunofluorescence (Fig. 6 and 7) assays. As evident from Table 1, the closest similarity was found to 3D adhesions, except that the latter do not contain the phosphorylated form of FAK [15], which was present in the stitch adhesions. The differences in molecular composition we established, combined with the specific morphology and localization, define stitch adhesions as a distinct class of cell adhesions.
Table 1.
Comparison of the molecular composition of the known cell-matrix adhesions (adapted from [15]) with the addition of stitch adhesions (righthand column)
| Component | Focal adhesions |
Fibrillar adhesions | 3D adhesions | Stitch adhesions | |
|---|---|---|---|---|---|
| β1 | + | + | + | + | (IF/IP) |
| α5 | −* | + | + | + | (IF) |
| β3 | + | − | − | − | (IF) |
| αV | + | − | − | − | (IF) |
| F-actn | + | + | + | + | (IF/IP) |
| tensin | + | + | + | + | (IF/IP) |
| talin | + | + | + | + | (IP) |
| paxillin | + | − | + | + | (IF/IP) |
| Vinculin | + | − | + | + | (IF/IP) |
| p-FAK | + | − | − | + | (IF) |
| fibronectin | −* | + | + | + | (IF/IP) |
IF – immunofluorescence; IP – immunoprecipitation
except at the periphery
In the experimental system we employed, the formation of adhesion stitches was triggered by addition of purified fibronectin. Both plasma and cellular FN organized similar stitches, which indicated that the EDA, EDB and the variable regions were not essential for the process, since plasma FN lacks these regions [44]. Added fibronectin was captured by α5β1, as this is the integrin capable of binding soluble fibronectin molecules in their compact inactive conformation [45]. The availability of α5β1 on the cell surface was most probably not affected by CHX treatment, because its degradation depends on binding to FN [46] and cycloheximide even appears to stabilize the mature β1 subunit [47]. Our immunofluorescence experiments also detected both of these integrin subunits on the surface of CHX-treated fibroblasts with preferential localization at stitch adhesions and focal contacts (Fig. 6).
The presence of soluble FN in the culture medium might suggest that binding to integrins could occur over the entire cell surface, but instead, the stitches developed in the area of closest contact between adjacent cells, which typically included the leading edge of cells. This specific localization could be explained by the findings of Sheetz’s group, who demonstrated that FN-liganded integrins attach to the cytoskeleton preferentially at the dorsal portion of the leading edge and are released at the endoplasm–ectoplasm boundary [48]. Connecting to actin bundles, which was also documented by us, is necessary for transmission of tension forces, required for unfolding of bound fibronectin molecules and formation of the stitch adhesions.
According to the generally accepted view, attachment of FN to the substrate or anchoring at focal adhesions is necessary to provide support for FN, which can then be stretched and engaged in polymerization [3]. We speculate that during the formation of stitches, support for FN molecules is provided by integrins belonging to adjacent/different cells (Fig. 10). This allows the actin cytoskeletons of the neighboring cells to work against each other, thus providing simultaneous stretching forces and anchoring for the forming stitches. At the same time, each of the cells is firmly attached to the vitronectin substrate through αVβ3-positive focal contacts (Fig. 6). Because this model demands at least two sites for binding of at least two integrin heterodimers, fragments of the FN molecule, even those containing integrin-binding sites, would be predicted to be ineffective for the formation of stitches. This prediction was confirmed by our results with 70 kD and 120kD FN fragments, which were not able to trigger establishment of stitch-type contacts (Fig. 2, C and D).
An interesting characteristic of stitch adhesions is the almost uniform length of their fibrils. The established model for fibronectin fibrillogenesis postulates that ligated α5β1 integrins translocate centripetally along stress fibers, using focal adhesions as an anchor to stretch FN molecules and induce formation of FN fibrils [10]. The end of the fibrils towards the cell edge is immobile, whereas the end toward the cell center extends [49]. Detachment of the extending end from the cell surface halts the growth, leads to eventual shrinking and deposition of the fibril on the cell surface or the substrate. This model does not pose any limit on the length of the FN fibrils being generated and indeed, cells produce fibrils of variable size. Studying the formation of the stitches, however, we found that the almost uniform final length of approximately 9.5 μm was reached within three hours, and the incorporation of new FN molecules led to their thickening but not substantial elongation (Fig. 3).
The observed effect of CHX on fibronectin fibrillogenesis was reversible, and restoration of protein synthesis and secretion led to a recovery of normal fibronectin matrix deposition (Fig. 8). Interestingly, we did not observe any noticeable effects of preexisting StA on the morphology (length, orientation or thickness) of the FN fibrils that were organized by fibroblasts after release of the CHX inhibitory block. Absence of influence on matrix formation suggests that StA may instead fulfill a role related to establishment of cell-to-cell contacts. Some physiological processes require strong associations between neighboring cells. During wound healing, attachment between myofibroblasts in granulation tissue is necessary to secure wound contraction. Such associations are provided by the fibronexus, a complex between fibronectin and actin in the form of fibronectin stitches at sites of intercellular contact [50]. Fibronectin stitches have also been attributed to formation of epithelioid colonies by chondrocytes, an essential process for cartilage matrix synthesis [24]. Thus, stitch adhesions may represent a different form of FN utilization in which fibronectin fibrils connect adjacent cells instead of building up the extracellular matrix environment.
Our results demonstrated that fibroblasts organize exogenous FN in a different way when protein synthesis and secretion were inhibited. This raises the possibility that normal fibrillogenesis may depend on the presence of specific protein(s) secreted by fibroblasts. The simplest scenario involves a secreted protein that interacts with FN and helps to present it to the cell in a form suitable for the typical fibrillogenesis. A possible candidate for such a protein supporting FN fibrillogenesis is collagen. Formation of collagen fibrils not only depend on preexisting FN matrix [51,52] but can also stimulate its production [25]. Both proteins are secreted together [53] and in addition, Dessau at al. have shown that chondrocytes form only short intercellular fibronectin stitches in the absence of extracellular collagen fibers [24]. Whether this is true also for fibroblasts remains to be elucidated.
In conclusion, we show here that when protein synthesis and fibronectin secretion are blocked, fibronectin fibrillogenesis is directed to a mechanism of stitch adhesion formation with specific fibronectin-containing cell-cell adhesions connecting neighboring fibroblasts. This distinct type of adhesions offers an interesting, useful model system for studying conversion of soluble fibronectin into fibrils and the effects of organized FN on cell physiology.
Highlights.
Fibroblasts with blocked protein synthesis use exogenous fibronectin to organize a new type of cell-cell adhesions – “stitch adhesions”.
Stitch adhesions are positioned at the dorsal cell surface and connect adjacent cells by short (9.5 μm), uniform and parallel fibronectin fibrils.
Stitch adhesions contain exclusively α5β1 integrins and connect to the actin cytoskeleton through adhesion plaques containing talin, tensin, α-actinin, vinculin, paxillin and a phosphorylated form of focal adhesion kinase.
Acknowledgements
This work was supported by the Intramural Research Program of the NIH, NIDCR and in part by Bulgarian Ministry of Education and Science, Contract DO1-154/28/08/2018/”Scientific Infrastructure on Cell Technologies in Biomedicine (SICTB)”
Footnotes
Competing interests
Authors declare no competing interests.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- [1].Hynes RO, Fibronectins. New York: Springer-Verlag, 1990. http://www.springer.com/la/book/9781461279402 [Google Scholar]
- [2].Pankov R, Yamada KM, Fibronectin at a glance, J. Cell Sci. 115 (2002) 3861–3863. http://jcs.biologists.org/content/115/20/3861.long [DOI] [PubMed] [Google Scholar]
- [3].Singh P, Carraher C, Schwarzbauer JE, Assembly of fibronectin extracellular matrix, Annu. Rev. Cell Dev. Biol. 26 (2010) 397–419. https://www.annualreviews.org/doi/abs/10.1146/annurev-cellbio-100109-104020 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [4].George EL, Georges-Labouesse EN, Patel-King RS, Rayburn H, Hynes RO, Defects in mesoderm, neural tube and vascular development in mouse embryos lacking fibronectin, Development, 119 (1993) 1079–1091. http://dev.biologists.org/content/119/4/1079.short [DOI] [PubMed] [Google Scholar]
- [5].Plow EF, Haas TA, Zhang L, Loftus J, Smith JW, Ligand binding to integrins, J. Biol. Chem. 275 (2000) 21785–21788. http://www.jbc.org/content/275/29/21785.short [DOI] [PubMed] [Google Scholar]
- [6].Humphries JD, Byron A, Humphries MJ, Integrin ligands at a glance, J. Cell Sci. 119 (2006) 3901–3903. http://jcs.biologists.org/content/119/19/3901.short [DOI] [PMC free article] [PubMed] [Google Scholar]
- [7].Ruoslahti E, Öbrink B, Common principles in cell adhesion. Exp. Cell Res. 227 (1996) 1–11. https://www.sciencedirect.com/science/article/pii/S0014482796902438 [DOI] [PubMed] [Google Scholar]
- [8].Aota SI, Nomizu M, M Yamada K, The short amino acid sequence Pro-His-Ser-Arg-Asn in human fibronectin enhances cell-adhesive function, J. Biol. Chem. 269 (1994) 24756–24761. http://www.jbc.org/content/269/40/24756.short [PubMed] [Google Scholar]
- [9].Schwarzbauer JE, Sechler JL, Fibronectin fibrillogenesis: a paradigm for extracellular matrix assembly, Curr. Opin. Cell Biol. 11 (1999) 622–627. https://www.sciencedirect.com/science/article/pii/S0955067499000174 [DOI] [PubMed] [Google Scholar]
- [10].Pankov R, Cukierman E, Katz BZ, Matsumoto K, Lin DC, Lin S, Hahn C, Yamada KM, Integrin dynamics and matrix assembly: tensin-dependent translocation of α5β1 integrins promotes early fibronectin fibrillogenesis, J. Cell Biol. 148 (2000) 1075–1090. http://jcb.rupress.org/content/148/5/1075 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [11].Zamir E, Katz M, Posen Y, Erez N, Yamada KM, Katz BZ, Lin S, Lin DC, Bershadsky A, Kam Z, Geiger B, Dynamics and segregation of cell-matrix adhesions in cultured fibroblasts, Nat. Cell Biol. 2 (2000) 191–196. https://www.nature.com/articles/ncb0400_191 [DOI] [PubMed] [Google Scholar]
- [12].Aguirre KM, McCormick RJ, Schwarzbauer JE, Fibronectin self-association is mediated by complementary sites within the amino-terminal one-third of the molecule, J. Biol. Chem. 269 (1994) 27863–27868. http://www.jbc.org/content/269/45/27863.short [PubMed] [Google Scholar]
- [13].Hocking DC, Sottile J, McKeown-Longo PJ, Fibronectin's III-1 module contains a conformation-dependent binding site for the amino-terminal region of fibronectin, J. Biol. Chem. 26 (1994) 19183–19187. http://www.jbc.org/content/269/29/19183.short [PubMed] [Google Scholar]
- [14].Ingham KC, Brew SA, Huff S, Litvinovich SV, Cryptic self-association sites in type III modules of fibronectin, J. Biol. Chem. 272 (1997). 1718–1724. http://www.jbc.org/content/272/3/1718.short [DOI] [PubMed] [Google Scholar]
- [15].Cukierman E, Pankov R, Stevens DR, Yamada KM, Taking cell-matrix adhesions to the third dimension, Science, 294 (2001) 1708–1712. http://science.sciencemag.org/content/294/5547/1708 [DOI] [PubMed] [Google Scholar]
- [16].Cukierman E, Pankov R, Yamada KM, Cell interactions with three-dimensional matrices, Curr. Opin. Cell Biol. 14 (2002) 633–640. https://www.sciencedirect.com/science/article/pii/S0955067402003642 [DOI] [PubMed] [Google Scholar]
- [17].Whittard JD, Akiyama SK, Activation of β1 integrins induces cell-cell adhesion, Exp. Cell Res. 263 (2001) 65–76. https://www.sciencedirect.com/science/article/pii/S0014482700950997 [DOI] [PubMed] [Google Scholar]
- [18].Lefort CT, Wojciechowski K, Hocking DC, N-cadherin cell-cell adhesion complexes are regulated by fibronectin matrix assembly, J. Biol. Chem. 286 (2011) 3149–3160. http://www.jbc.org/content/286/4/3149.short [DOI] [PMC free article] [PubMed] [Google Scholar]
- [19].Ali IU, Mautner V, Lanza R, Hynes RO, Restoration of normal morphology, adhesion and cytoskeleton in transformed cells by addition of a transformation-sensitive surface protein, Cell, 11(1977) 115–126. https://www.sciencedirect.com/science/article/pii/0092867477903221 [DOI] [PubMed] [Google Scholar]
- [20].Mautner V, Hynes RO, Surface distribution of LETS protein in relation to the cytoskeleton of normal and transformed cells, J. Cell Biol. 75 (1977) 743–768. http://jcb.rupress.org/content/75/3/743.abstract [DOI] [PMC free article] [PubMed] [Google Scholar]
- [21].Singer II, The fibronexus: a transmembrane association of fibronectin-containing fibers and bundles of 5 nm microfilaments in hamster and human fibroblasts, Cell, 16 (1979) 675–685. https://www.sciencedirect.com/science/article/pii/0092867479900400 [DOI] [PubMed] [Google Scholar]
- [22].Singer II, Paradiso PR, A transmembrane relationship between fibronectin and vinculin (130 kd protein): serum modulation in normal and transformed hamster fibroblasts, Cell, 24 (1981) 481–492. https://www.sciencedirect.com/science/article/pii/0092867481903391 [DOI] [PubMed] [Google Scholar]
- [23].Singer II, Fibronexus formation is an early event during fibronectin-induced restoration of more normal morphology and substrate adhesion patterns in transformed hamster fibroblasts, J. Cell Sci 56 (1982) 1–20. https://europepmc.org/abstract/med/6820020 [DOI] [PubMed] [Google Scholar]
- [24].Dessau W, Sasse J, Timpl R, Jilek F, von der Mark K, Synthesis and extracellular deposition of fibronectin in chondrocyte cultures. Response to the removal of extracellular cartilage matrix, J. Cell Biol. 79 (1978) 342–355. http://jcb.rupress.org/content/79/2/342 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [25].Dzamba BJ, Wu H, Jaenisch R, Peters DM, Fibronectin binding site in type I collagen regulates fibronectin fibril formation, J. Cell Biol. 121 (1993) 1165–1172. http://jcb.rupress.org/content/121/5/1165 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [26].Miyamoto S, Akiyama SK, Yamada KM, Synergistic roles for receptor occupancy and aggregation in integrin transmembrane function, Science 267 (1995) 883–885. http://science.sciencemag.org/content/267/5199/883 [DOI] [PubMed] [Google Scholar]
- [27].Akiyama SK, Yamada SS, Chen WT, Yamada KM, Analysis of fibronectin receptor function with monoclonal antibodies: roles in cell adhesion, migration, matrix assembly, and cytoskeletal organization, J. Cell Biol. 109 (1989) 863–875. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2115712/ [DOI] [PMC free article] [PubMed] [Google Scholar]
- [28].Nagai T, Yamakawa N, Aota SI, Yamada SS, Akiyama SK, Olden K, Yamada KM, Monoclonal antibody characterization of two distant sites required for function of the central cell-binding domain of fibronectin in cell adhesion, cell migration, and matrix assembly, J. Cell Biol. 114 (1991) 1295–1305. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2289135/ [DOI] [PMC free article] [PubMed] [Google Scholar]
- [29].Miekka SI, Ingham KC, Menache D, Rapid methods for isolation of human plasma fibronectin, Thromb. Res. 27 (1982) 1–14. http://www.thrombosisresearch.com/article/0049-3848(82)90272-9/fulltext [DOI] [PubMed] [Google Scholar]
- [30].McKeown-Longo PJ, Mosher DF, Interaction of the 70,000-mol-wt amino-terminal fragment of fibronectin with the matrix-assembly receptor of fibroblasts. J. Cell Biol. 100 (1985) 364–374. http://jcb.rupress.org/content/100/2/364 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [31].Knox P, Crooks S, Rimmer CS, Role of fibronectin in the migration of fibroblasts into plasma clots, J. Cell Biol. 102 (1986) 2318–2323. http://jcb.rupress.org/content/102/6/2318 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [32].Yamada KM, Olden K, Pastan I, Transformation-sensitive cell surface protein: isolation, characterization, and role in cellular morphology and adhesion, Ann. N. Y. Acad. Sci, 312 (1978) 256–277. https://nyaspubs.onlinelibrary.wiley.com/doi/abs/10.1111/j.1749-6632.1978.tb16807.x [DOI] [PubMed] [Google Scholar]
- [33].Won CH, Kwon OS, Kang YJ, Yoo HG, Lee DH, Chung JH, Kim KH, Park WS, Park NH, Cho K, Kwon SO, Choi JS, Eun HC, Comparative secretome analysis of human follicular dermal papilla cells and fibroblasts using shotgun proteomics, BMB Rep. 45 (2012) 253–8. http://www.bmbreports.org/journal/view.html?volume=45&number=4&spage=253 [DOI] [PubMed] [Google Scholar]
- [34].Schneider-Poetsch T, Ju J, Eyler DE, Dang Y, Bhat S, Merrick WC, Green R, Shen B, Liu JO, Inhibition of eukaryotic translation elongation by cycloheximide and lactimidomycin, Nat. Chem. Biol. 6 (2010) 209 https://www.nature.com/articles/nchembio.304 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [35].Pankov R, Momchilova A, Fluorescent Labeling Techniques for Investigation of Fibronectin Fibrillogenesis (Labeling Fibronectin Fibrillogenesis), in: Even-Ram S, Artym V (Eds.), Extracellular Matrix Protocols. Methods in Molecular Biology (Methods and Protocols), vol 522 Humana Press; (2009) pp. 261–274. https://link.springer.com/protocol/10.1007/978-1-59745-413-1_18#citeas [DOI] [PubMed] [Google Scholar]
- [36].Sun Z, Guo SS, Fässler R, Integrin-mediated mechanotransduction, J. Cell Biol. 215 (2016) 445–456. http://jcb.rupress.org/content/215/4/445 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [37].Winograd-Katz SE, Fässler R, Geiger B, Legate KR, The integrin adhesome: from genes and proteins to human disease, Nat. Rev. Mol. Cell Biol. 15 (2014) 273–288. https://www.nature.com/articles/nrm3769 [DOI] [PubMed] [Google Scholar]
- [38].Horton ER, Humphries JD, James J, Jones MC, Askari JA, Humphries MJ, The integrin adhesome network at a glance, J. Cell Sci. 129 (2016) 4159–4163. http://jcs.biologists.org/content/129/22/4159.eLetters [DOI] [PMC free article] [PubMed] [Google Scholar]
- [39].Rebelo S, Vieira SI, da Cruz e Silva EF, da Cruz e Silva OA, Monitoring‘‘De Novo"APP Synthesis by Taking Advantage of the Reversible Effect of Cycloheximide, Am. J. Alzheimers Dis. Other Demen.23 (2009) 602–608. http://journals.sagepub.com/doi/abs/10.1177/1533317508323572 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [40].Berrier AL, Yamada KM, Cell–matrix adhesion J Cell Physiol. 213 (2007) 565–573. https://onlinelibrary.wiley.com/doi/full/10.1002/jcp.21237 [DOI] [PubMed] [Google Scholar]
- [41].Costa P, Parsons M, New insights into the dynamics of cell adhesions. Int. Rev. Cell Mol. Biol. 283, (2010) 57–91. https://www.sciencedirect.com/science/article/pii/S1937644810830023 [DOI] [PubMed] [Google Scholar]
- [42].Bizik J, Kankuri E, Ristimäki A, Taieb A, Vapaatalo H, Lubitz W, Vaheri A, Cell–cell contacts trigger programmed necrosis and induce cyclooxygenase-2 expression, Cell Death Differ. 11 (2004) 183–195. https://www.nature.com/articles/4401317 [DOI] [PubMed] [Google Scholar]
- [43].Schaller MD, Cellular functions of FAK kinases: insight into molecular mechanisms and novel functions, J. Cell Sci. 123 (2010) 1007–1013. http://jcs.biologists.org/content/123/7/1007.short [DOI] [PubMed] [Google Scholar]
- [44].Ffrench-Constant C, Alternative Splicing of Fibronectin–Many Different Proteins, Exp. Cell Res. 221 (1995) 261–271. https://www.sciencedirect.com/science/article/pii/S0014482785713742 [DOI] [PubMed] [Google Scholar]
- [45].Huveneers S, Truong H, Fässler R, Sonnenberg A, Danen EH, Binding of soluble fibronectin to integrin α5β1–link to focal adhesion redistribution and contractile shape, J. Cell Sci. 121 (2008) 2452–2462. http://jcs.biologists.org/content/121/15/2452.short [DOI] [PubMed] [Google Scholar]
- [46].Lobert VH, Brech A, Pedersen NM, Wesche J, Oppelt A, Malerød L, Stenmark H, Ubiquitination of α5β1 integrin controls fibroblast migration through lysosomal degradation of fibronectin-integrin complexes, Dev. Cell, 19 (2010) 148–159. https://www.sciencedirect.com/science/article/pii/S153458071000300X [DOI] [PubMed] [Google Scholar]
- [47].Vekeman S, Jaspers M, Cassiman JJ, Inhibition of the degradation of the precursor and of the mature β1 integrin subunit by different protein synthesis inhibitors and by ATP depletion, FEBS Lett. 327 (1993) 207–212. https://febs.onlinelibrary.wiley.com/doi/abs/10.1016/0014-5793%2893%2980171-P [DOI] [PubMed] [Google Scholar]
- [48].Nishizaka T, Shi Q, Sheetz MP, Position-dependent linkages of fibronectin–integrin–cytoskeleton, Proc. Natl. Acad. Sci. USA, 97 (2000) 692–697. http://www.pnas.org/content/97/2/692.short [DOI] [PMC free article] [PubMed] [Google Scholar]
- [49].Ohashi T, Kiehart DP, Erickson HP, Dual labeling of the fibronectin matrix and actin cytoskeleton with green fluorescent protein variants, J. Cell Sci. 115 (2002) 1221–1229. http://jcs.biologists.org/content/115/6/1221.short [DOI] [PubMed] [Google Scholar]
- [50].Singer II, Kawka DW, Kazazis DM, Clark RA, In vivo co-distribution of fibronectin and actin fibers in granulation tissue: immunofluorescence and electron microscope studies of the fibronexus at the myofibroblast surface, J Cell Biol. 98 (1984) 2091–2106. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2113047/ [DOI] [PMC free article] [PubMed] [Google Scholar]
- [51].McDonald JA, Kelley DG, Broekelmann TJ, Role of fibronectin in collagen deposition: Fab'to the gelatin-binding domain of fibronectin inhibits both fibronectin and collagen organization in fibroblast extracellular matrix, J Cell Biol. 92 (1982) 485–492. http://jcb.rupress.org/content/92/2/485 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [52].Velling T, Risteli J, Wennerberg K, Mosher DF, Johansson S, Polymerization of type I and III collagens is dependent on fibronectin and enhanced by integrins α11β1and α2β1. J Biol. Chem. 277 (2002) 37377–37381. http://www.jbc.org/content/277/40/37377 [DOI] [PubMed] [Google Scholar]
- [53].Ledger PW, Uchida N, Tanzer ML, Immunocytochemical localization of procollagen and fibronectin in human fibroblasts: effects of the monovalent ionophore, monensin, J Cell Biol. 87 (1980) 663–671. http://jcb.rupress.org/content/87/3/663 [DOI] [PMC free article] [PubMed] [Google Scholar]