

Abstract
Coral reefs are degrading globally leading to a catastrophic loss of biodiversity. While shifts in the species composition of communities have been well documented associated with habitat change, the mechanisms that underlie change are often poorly understood. Our study experimentally examines the effects of coral degradation on the trait-mediated effects of predators on the morphology, behaviour and performance of a juvenile coral reef fish. Juvenile damselfish were exposed to predators or controls (omnivore or nothing) in seawater that had flowed over either live or dead-degraded coral over a 45d period. No interaction between water source and predator exposure was found. However, fish exposed to degraded water had larger false eyespots relative to the size of their true eyes, and were more active, both of which may lead to a survival advantage. Non-consumptive effects of predators on prey occurred regardless of water source and included longer and deeper bodies, large false eyespots that may distract predator strikes away from the vulnerable head region, and shorter latencies in their response to a simulated predator strike. Research underscores that phenotypic plasticity may assist fishes in coping with habitat degradation and promote greater resilience to habitat change than may otherwise be predicted.
Subject terms: Behavioural ecology, Climate-change ecology
Introduction
Habitat degradation is one of the key drivers of a global catastrophic loss of biodiversity1–3. The loss and modification of ecosystem engineers, such as large canopy trees or rapidly growing hard corals, impacts communities through changes in the diversity of the landscape and services it provides4. Because habitat degradation is usually a gradual process, its impact on communities is first detected through changes in the physiology of species5. Physiological challenges sum to whole-animal changes in body condition by affecting an individual’s overall health and ability to obtain limited resources6,7. The way an individual changes physiologically, and how these changes alter interactions within a community, determines an individual’s fitness and the resilience of the species to habitat degradation8. While shifts in the species composition of communities have been well documented associated with habitat change (e.g.9,10), the mechanisms that underlie change are often poorly understood.
Changes to the composition of landscape-forming species alters the sensory environment of the animals that live within the habitat11,12. The interactions between individuals that make up community dynamics are framed within a foundation of current and historical information from their local environment13,14. The information within which to frame decisions changes as the availability of sensory information changes. For instance, a loss of a forest canopy alters the visibility and passage of air through the environment, altering the information available from visual and olfactory senses15. This change in the availability of fundamental information can alter the outcome of interactions between individuals and species, which are the basis of community dynamics. Predator-prey interactions have evolved to be information-demanding, because predators have usually coevolved with their prey, making both predator attack and prey escape strategies highly efficient16, Alterations to the flow and types of information through the community are therefore very likely to affect predator-prey dynamics and may be one of the fundamental mechanisms that underlie the rapid change in community composition that comes with habitat change.
Prey have evolved to be highly responsive to the threat of predation, and predation is viewed as the driving force underlying physiology, morphology and behaviour through selective loss. Not only do predators have direct (lethal or consumptive) effects on prey, but they also have indirect (non-consumptive), trait-mediated effects on prey through the threat of predation. Indeed, meta-analyses find that these non-consumptive ‘fear-effects’ have more of an impact on prey ecology, and consequently community dynamics, than their consumptive effects alone. This has been well demonstrated in terrestrial17, freshwater18 and marine systems19,20. Studies of aquatic organisms have found that the sight or smell of a predator can lead to changes in life history traits (e.g., developmental rates and hatching times21;), behaviour (e.g., development of neophobic phenotypes22;) and alter body morphology23 to forms that are best suited to surviving in a location where the threat of predation is high. Landmark examples of this are the deepening of the body in crucian carp (Carassius carassius) to the smell of predatory pike (Esox lucius)24, and the increase in the relative size of the false eyespot in damselfish (Pomacentrus amboinensis) prey, believed to attract predator strikes to the less vulnerable tail region23. Developmental plasticity in these morphological characteristics has been shown to affect performance and survival23,25,26, underscoring the importance of trait-mediated effects in prey ecology and fitness.
Many aspects of predator-prey interactions in aquatic environments are mediated by olfactory cues and changes in the chemistry of the surrounding environment can have impacts on the efficacy of cues27. Learning and updating information about predators principally occurs for many aquatic organisms through chemical associative learning, whereby damage-released chemical cues (known as alarm odours) label other unrelated stimuli (e.g., sight, sound or smell of a predator) as a threat when they co-occur28. Recent research has shown that many fishes lose their ability to use alarm odours in degraded coral reef habitats due to changes in the background chemistry of the environment, which directly impacts survival in the field29. While studies have shown that prey survival is compromised in degraded habitats due to uninformed decisions30,31, it is unknown whether the degradation of habitats such as coral reefs alters the action of trait-mediated antipredator solutions.
The current study examines the effects of habitat degradation on trait-mediated antipredator traits in a coral reef fish. We focus on the coral reef ecosystem as it is one of the most biodiverse on the planet and because it is subject to degradation on a global scale, changing from hard-coral to algal dominated landscapes9. Previous studies have highlighted the importance for prey of olfactory information on the activity of predators28, and the smell of predators alone have been previously shown to alter prey behaviour and morphology in water from healthy coral reefs19,23,25. The current study examines how water from healthy and dead-degraded habitats can influence the development of body morphology and escape performance in a habitat generalist damselfish, Pomacentrus amboinensis. This is a particularly interesting model species as it has a false eyespot (i.e., ocellus) as a juvenile whose size has been found to correlate with survival23. Predictions from previous research suggest water source will interact with the type of stimulus fish to which focal prey fish were exposed (i.e., blank control, herbivore or predator), because degraded water has been found to prevent the labelling of the predator as a threat32,33. Consequently, we expect that predator-associated trait-mediated effects, such as changes to body depth and the size of the false eyespot, would be negligible in degraded habitats compared to fish reared in water that has flowed over live coral.
Materials and Methods
Study system and species
The study was conducted at the Lizard Island research station (14°40'43.07"S, 145°26'53.35"E), on the northern Great Barrier Reef, Australia from October to December 2017. Newly metamorphosed damselfish (family Pomacentridae) were collected from light traps that had been deployed overnight at least 30 m from the reef edge and over a water column 8–20 m deep. The focal damselfish species, Pomacentrus amboinensis, is a common component of Indo-Pacific coral reefs where it is an omnivorous planktivore34.
The predator species used in the experiment was the dusky dottyback, Pseudochromis fuscus (family: Pseudochromidae), which is a common mid-sized predator on the shallow reefs throughout the Indo-Pacific. The species is a voracious predator of recently settled fishes during the recruitment season35 and is common in the same habitats used by P. amboinensis. The omnivorous goby, Amblygobius phalaena36, was used as the stimulus fish in experimental controls to account for the effect of exposing P. amboinensis to visual and olfactory cues of any heterospecific fish species. It is similar in body shape to the predator, Ps. fuscus, and is common on the base of the reef where P. amboinensis occurs34. Both species were caught by SCUBA divers using a small hand net and a dilute clove oil solution that anaesthetizes the fish and facilitates their capture from the reef.
Live bushy hard coral (Pocillopora damicornis) and dead-degraded coral covered in a mixture of algae and sessile invertebrates (450–500 ml in volume; dimensions ~200 × 150 × 150 mm) were collected from fringing reefs around Lizard Island, and placed in aerated 60 L header tanks of flow-through seawater37. Live and dead Po. damicornis are common nursery habitats for newly recruited damselfish, and commonly found within Ps. fuscus territories35.
Experimental design and protocol
To examine the effect of water source on a prey’s morphological responses to a predation threat, individual P. amboinensis were exposed to six experimental treatments combining olfactory and visual cues of a predator (the dottyback, Ps. fuscus), a non-predator (the herbivorous goby, A. phalaena), or a blank control (receiving no cue from adult fish sources) in either seawater that had flowed over live coral or over dead-degraded coral, in a 3 × 2 design. Given that degraded water sources can limit the labelling of predators as threat31–33, we expected P. amboinensis from this treatment would not develop trait-mediated responses towards the dottyback and be morphologically similar to prey exposed to the control or herbivore fish.
Prior to being randomly allocated to one of the 6 treatment combinations, light trap caught P. amboinensis individuals were conditioned to recognize the sight and olfactory cues of P. fuscus in either water from live coral or dead-degraded coral. Conditioning was undertaken by chemical associative learning, using a commonly used protocol. Conspecific chemical alarm odours from five light trap caught P. amboinensis were added to 15 ml of treatment water (from live or dead-degraded coral header tanks) and 500 ml seawater from a 35 L tank that had 4 Ps. fuscus sitting under aeration, but no water flow for 1 hour. Alarm odours were collected from fish killed by cold shock (see animal welfare below) and superficially lacerated with five vertical cuts per side from a scalpel38. Ps. fuscus were also added to the tanks within a plastic bag filled with aerated seawater so that P. amboinensis were trained to associate the sight of a dottyback as a threat. This training procedure has been found to substantially increase the probability of survival in the Ambon damselfish39, and is necessary to make sure that prey can recognise the cues of the predator species within the appropriate environmental context into which they settled. Fish were specifically trained in the water treatment where they were going to be reared and tested, in order to maintain a realistic and consistent scenario throughout this study, as well as to capture any effect that the water treatment has on the ability of P. amboinensis to recognise predators32,33. This approach ensured that all fish within a water type (live coral vs. degraded coral) had the same baseline predator experience before the commencement of the study.
Trained damselfish (N = 144) were transferred to purpose-built experimental predator–prey tanks ( 52 L; see Supplementary Fig. S1). Each experimental tank consisted of four 6.1 L sections for the stimulus species (i.e., dottyback, goby or blank control) and 12 individual prey compartments (2.3 L). The design of the experimental tank allowed each of the stimulus compartments to exchange visual and chemical cues only with the three prey compartments in front (see Supplementary Material). The experimental tank included a 1.5 cm layer of sand on the bottom, PVC tubes providing shelter to the stimulus species and the prey, and a plastic branching coral skeleton (Pocillopora sp. ~5 × 3 × 8 cm) at the back of each P. amboinensis compartment to provide topography. Experimental tanks were situated in an outdoor wet laboratory to ensure that all fish received natural temporal cues and the seawater was supplied via header tanks containing either live coral or dead-degraded coral (as described above). P. amboinensis were individually fed twice daily with 10 ml of Artemia sp. nauplii (mean 1245 Artemia/ml), while dottyback predators were fed two euthanized P. amboinensis recruits at the start and end of the day. Gobies were fed 5 ml of small crustaceans from the light trap catches at the same time of day as the predators. A small number of fish were lost in the first 2 d after the initiation of the experiment (17 out of 144 due to plumbing issues; representing 11 fish from live and 6 fish within dead coral water sources). These were restocked with fish that had been conditioned together with the original fish used to stock the tanks.
Water from the Lizard Island research station seawater system was used for this study. We expect that the chemistry of the seawater fed into the system that formed the background water source for all treatments represented a mixture of healthy and degraded coral odours, given that the seawater is sourced from the island’s shallow backreef. Seawater from the station was passed through a one micron filter bag before entering the 60 L header tanks containing either the live Po. damicornis or the dead-degraded corals as above. A pipe on the opposite side of the header tank, allowed water to flow into the central chamber of the experimental tank that contained the stimulus species (i.e., dottyback, goby or nothing). Water then flowed through six holes in the Perspex divider to each of the compartments containing individual P. amboinensis and exited through holes in the back of their compartments. Dye trials confirmed this flow pattern. Given the arrangement of the tanks, water flow, and panels, each P. amboinensis received visual cues from the stimulus species and chemical cues from a combination of the water source (live or degraded coral) and the stimulus (dottyback, goby or blank control). P. amboinensis could neither see nor smell conspecifics in the nearby chambers due to the design of the tanks and the direction of the water flow.
Stimulus fishes (i.e., Ps. fuscus or A. phalaena) were replaced every 2 weeks, and specific individuals rotated every 2 days within treatments. Coral substrata were kept in the aquarium system for a maximum of one week prior to being replaced with newly collected material. In this way, each P. amboinensis was exposed to 42 days of continuous treatment, but the specific identity of the stimulus fishes and coral was frequently shifted within treatment. P. amboinensis were returned to their original compartments after being periodically photographed.
Morphometric measures
P. amboinensis were individually placed inside a clip-seal bag containing aerated seawater (either live or degraded) and photographed on their side against a centimeter grid at the start (day one), midway through (day 22) and at the end (day 45) of the experiment. Using the software ImageJ, a variety of morphometric measures were taken from the images: (i) standard length (SL), (ii) body depth, (iii) area of the eye, (iv) area of the black portion of the eye, (v) area of ocellus, and (vi) area of the black portion of the ocellus (Fig. S2). At the end of the study, fish were weighed by transferring fish onto tissue paper prior to placing them in a tared quantity of seawater. Upon recovery from stress fish were released onto the fringing reef.
Behavioural assessments
Behavioural assessments were conducted at the start (day 5), midway through (day 23) and at the end of the experiment (day 42). The procedure involved suspending a mirror (80 × 40 cm) over each tank at 45° so that focal fish could be filmed from above with a video camera positioned parallel to the tank and pointing at the mirror. The camera stood on a tripod and was always out of sight of the focal fish. After a 10 min acclimation period, individual P. amboinensis were fed 10 ml of Artemia sp. nauplii (mean 1245 Artemia/ml), while the video camera recorded their behaviour for 5 minutes at 30 fps. Similar to a previous study by Lönnstedt and colleagues23, the total number of feeding strikes and activity (i.e., number of seconds fish were not in shelter) were quantified from the first 120 s after the feeding event.
Routine swimming
After 45 days individual P. amboinensis were removed from their compartments and exposed to a repeatable drop stimulus using the methodology of McCormick and Allan40. The testing arena consisted of a transparent circular acrylic area (diameter 200 mm; height 70 mm), within a large opaque-sided plastic tank (585 × 420 × 330 mm; 60 L) with a transparent Perspex bottom to allow responses to be filmed from below as a silhouette. The water level was low (60 mm) to reduce movements in the vertical plane and the arena was lit by an LED light strip attached to the outer surface of the opaque tank. Fish were not fed overnight (~16 h) and the water within the arena matched the water within the holding tanks of the tested fish. Fish were placed in a 1-litre opaque tank of matching water within a water bath in a quiet area for 30–40 minutes prior to being carefully transferred into the test arena.
After a five minute acclimation period within the arena, fish were videoed at 30 fps for two minutes to obtain an estimate of their activity. Activity measured as the total distance moved over the 120 s period measured at one second intervals.
Escape performance assessment
Once routine swimming had been recorded, fish were exposed to a repeatable startle stimulus which followed the protocol of McCormick and Allan40. Fish were only startled when they moved to the middle portion of the tank and were oriented toward the drop stimulus, which is located in the center of the arena, allowing an individual to move an equal distance in any direction and standardising for fish position relative to the stimulus. A weight was released from an electromagnet and was governed by a piece of fishing line that was long enough such that the tapered tip only just touched the surface of the water. To avoid the occurrence of a premature escape response associated with visual stimulation, the weight was released from above through a 550 mm piece of 48.5 mm diameter PVC pipe with the bottom edge at a distance of 10 mm above the water level. Escape variables were only measured when prey performed a C-start (i.e., commencement of fast-start that results in the individual forming a C-shape). Escape responses were recorded at 480 frames per second (Casio EX-ZR1000) as a silhouette from below.
Kinematic variables associated with the fast-start response were analysed using the image-analysis software Image-J, with a manual tracking plug-in following standard protocols40. For analysis of the high-speed movements each fish was reduced to a single moving point. The point where each fish was tracked was standardised by following the same point on each fish (i.e., the position directly behind the eyes which corresponds to the widest part of the body). We chose to standardise tracking based on this point of the body as it the most stable and easiest to track owing to the small size of the juveniles. Distance to the drop stimulus was measured at the point of contact of the weight with the water surface. The following fast-start variables were measured40:
Response latency (ms) was measured as the time interval between the stimulus touching the water surface and the first detectable movement of the fish.
Response speed (m/s) was measured as the distance covered within a fixed time (24 ms). This fixed duration was based on the average duration (22.8 ms) of stage one and two (as defined above).
Maximum response speed (m/s) was measured as the maximum speed achieved at any time during stage one and stage two.
Ethics statement
This research was undertaken in accordance with the animal ethics guidelines and regulations of the James Cook University, and all protocols were approved by the James Cook University Animal Ethics Committee (Approval Numbers: A2408). Animal welfare issues are detailed in the Supplementary File.
Statistical analyses
Standard length (SL) was tested between water sources (Live coral, Dead-degraded coral), treatments (Control, Goby, Dottyback) and their interaction separately for the start and end of the experiment using general linear models (GLMs). Body weight was only measured at the end of the experiment, so had a more abbreviated analysis. Other body variables (e.g., body depth, max eye diameter) that were correlated with standard length were tested for effects of water source, treatments and their interaction using analysis of covariance, using SL as a covariate. A test of the assumption of homogeneity of slopes were run prior to running the reduced model. SL was not a significant covariate for tests on the proportion of ocellus to eye maximum diameter.
The routine swimming variable total distance moved was initially analysed with the fixed factors water source and treatment and SL as a covariate. SL and its interactions accounted for no significant variation so were dropped from the final analyses. Fast-start latency was correlated with distance to the drop stimulus, while mean speed and maximum speed had positive correlations (covariance) with SL, so these correlated variables were used as covariates in analyses. Because of the interdependence of kinematic variables an α’ = 0.0125 was used as the cut-off for interpretation to reduce the impact of type I error inflation.
All analyses were originally run as linear mixed effects models accounting for the tank effect, but this term was found to be of negligible importance (variance was usually <30% of the residual) and subsequently dropped from the analyses. For all analyses, assumptions of normality and homogeneity of variance were examined using residual analyses. Due to minor differences in sample sizes among treatment by water source combinations (n = ~24, though a small number died; for exact N see figure legends) Type III sums of squares were used in GLM analyses. Effect sizes are given as partial-eta-squared, which represents the proportion of the total variance in a dependent variable that is associated with the membership of different groups.
Results
Morphology
At the start of the experiment, there was a significant difference in the length of damselfish between water sources (F1,125 = 5.78, P = 0.02, ηp2 = 0.04), with fish in the live coral water treatment being slightly longer than those exposed to dead-degraded seawater (mean 12.0, 11.8 mm SL respectively, SE 0.07). This was likely due to the restocking of fish that were lost at the start of the experiment within the treatments (11 live vs 6 dead coral treatments). There was no difference in size-corrected body depth among treatments, water sources or their combination at the start of the experiment (P > 0.2). Initial trends in means size were very different from their patterns at the end of the experiment (below), suggesting that the small initial differences (i.e., 0.2 mm in length) had little effect on the final patterns found.
There was a significant difference in the size of damselfish at the end of the experiment among treatments (F2,126 = 3.77, P = 0.026, ηp2 = 0.06), though no difference between water sources, and no interaction between treatment and water source (P > 0.05). Fish were significantly longer when exposed to the predatory dottybacks, compared to when exposed to the goby or the blank controls (Fig. 1a). Body weight at the end of the experiment showed exactly the same patterns of significance among treatments as standard length (F2,127 = 3.86, P = 0.02, ηp2 = 0.06). There was also a similar pattern in the body depth with the effect of standard length removed. Body depth of experimental P. amboinensis differed among treatments (F2,126 = 4.07, P = 0.019, ηp2 = 0.06; Fig. 1b), though not between water sources, and there was no interaction between treatment and water source (P > 0.05).
Figure 1.
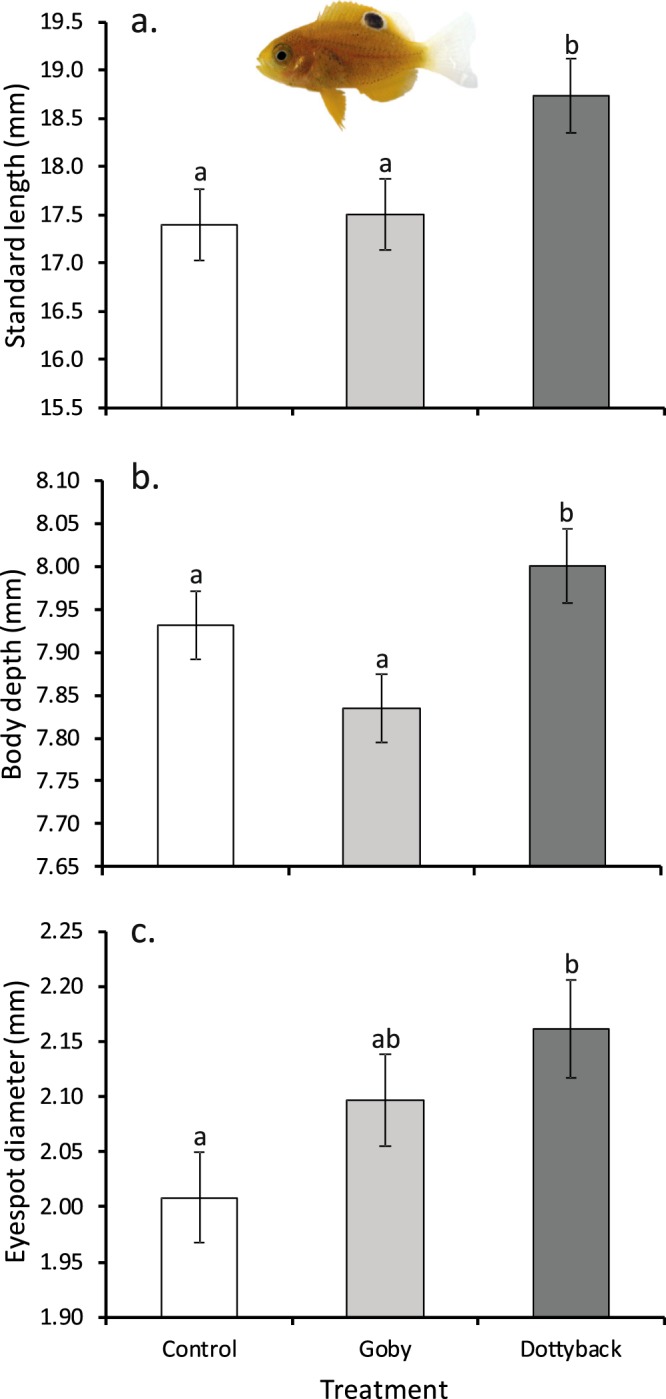
Comparison of (a) standard length, (b) body depth and (c) false eyespot diameter (mean ± SE) of juvenile Pomacentrus amboinensis exposed to an empty control, an omnivorous goby, or a predatory dottyback for 45 days. Data are averaged over water source as this had no effect on the variables. Values for body depth and eyespot diameter were adjusted for their covariation with standard length. Letters above bars represent post-hoc groupings of means. N = 47, 45, 41 (left to right). Photographic credit M. McCormick.
The maximum diameter of the eye was larger for damselfish reared in live coral water compared to those from dead coral water at the end of the experiment (F1,128 = 8.33, P = 0.005, ηp2 = 0.06; mean ± SE, Live 1.60 ± 0.01 mm, n = 66; Dead 1.56 ± 0.01 mm, n = 67), but there was no treatment effect or interaction (P > 0.3). The false eyespot diameter differed only among treatments (F2,127 = 3.23, P = 0.04, ηp2 = 0.05, with SL as a significant covariate; Fig. 1c), with those damselfish exposed to dottybacks possessing larger eyespots compared to controls. There was a difference in the proportion of the eyespot to eye maximum diameter among treatments (F2,127 = 4.0, P = 0.02, ηp2 = 0.06; Fig. 2a) and by water source (F1,127 = 4.31, P = 0.04, ηp2 = 0.03; Fig. 2b), but there was no interaction between factors (P = 0.25). False eyespots were proportionately larger in fish exposed to dottybacks, and in those reared in degraded water (Fig. 2b). The relationship between body weight and standard length (i.e., length standardized weight) differed at the level of the interaction between treatment and water source (F2,121 = 3.37, P = 0.04). Further analyses showed that this difference was caused by fish exposed to live-coral water and dottybacks being different in slope and intercept than all other relationships (ANCOVA Interaction F1,129 = 20.5, P < 0.0001), which did not differ from one another. P. amboinensis exposed live coral with dottybacks gained weight at a rate of 42.9 mg/mm, compared to 34.7 mg/mm of all the other treatment combinations combined.
Figure 2.
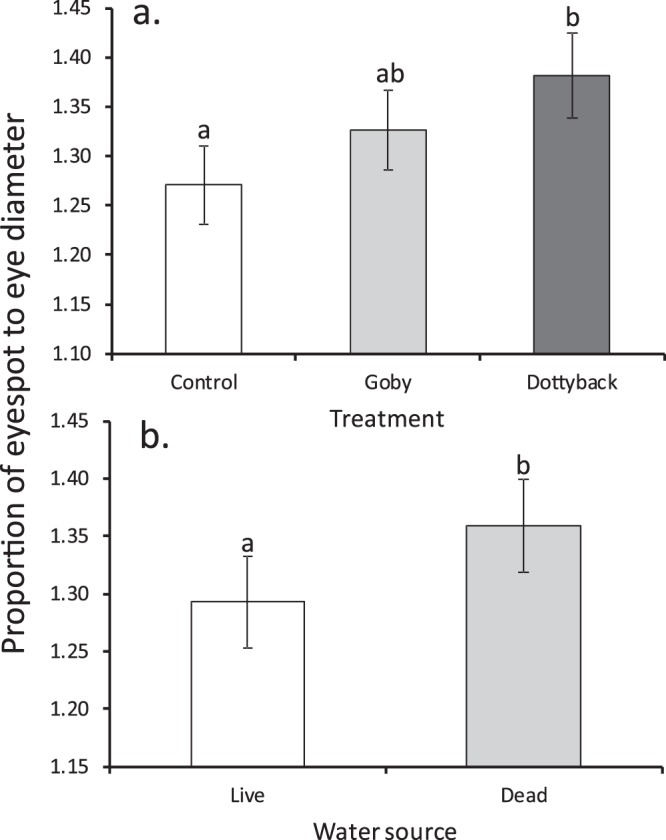
Comparison of proportion of the false eyespot to eye diameter of juvenile Pomacentrus amboinensis (mean ± SE) exposed for 45 days to: (a) three treatment levels of adult fish (blank control, an omnivorous goby, or a predatory dottyback), and (b) two water sources (live coral or dead-degraded coral). Letters above bars represent Fisher’s LSD groupings of means. N = 47, 45, 41 (top); 66, 67 (bottom).
Behaviour
Feeding rate on the fourth day after the initiation of the experiment differed among treatments (F2,126 = 5.61, P = 0.005, ηp2 = 0.08), but not by water source or their interaction (P > 0.2). P. amboinensis exposed to the goby or dottyback had one-third greater feeding rates than fish within the control treatment (Fig. 3). By the end of the experiment, the lack of effect of water source was still evident (P > 0.3), but the pattern among treatments had changed to a trend in which fish in the dottyback treatment eat more than the control fish (F2,127 = 3.07, P = 0.05, ηp2 = 0.05), while goby-exposed juveniles had an intermediate feeding rate.
Figure 3.
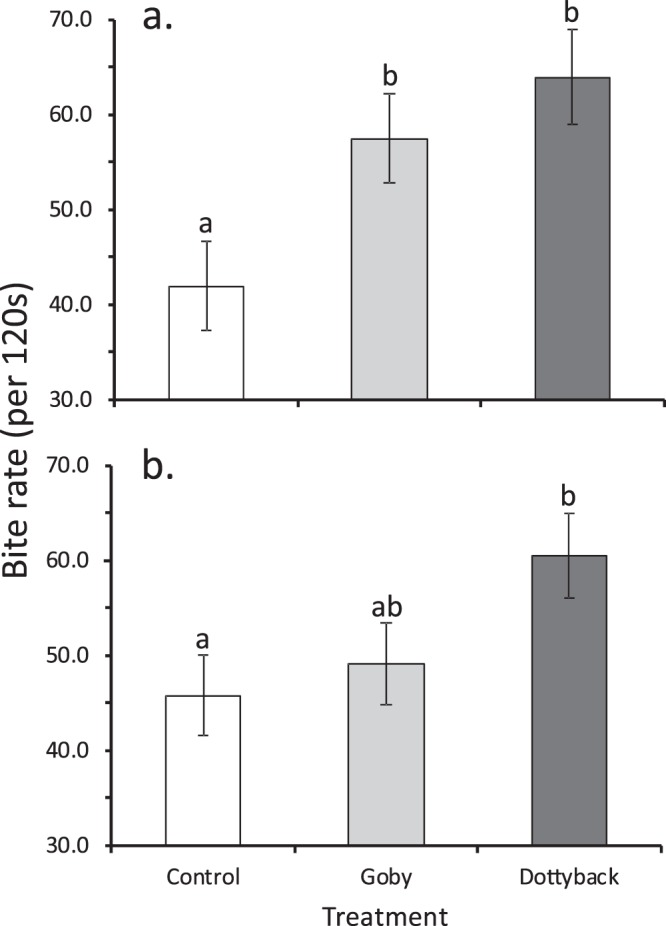
Bite rate (per 120 s; mean ± SE) of juvenile Pomacentrus amboinensis (a) 4 days after the start of the experiment and (b) at the end (day 45) of the experiment. Juvenile damselfish were exposed to three treatment levels: blank control, an omnivorous goby, or a predatory dottyback. Letters above bars represent Fisher’s LSD groupings of means. N = 46, 45, 41 (top), 47, 45, 41 (bottom).
Routine swimming and escape performance
The total distance moved by damselfish during routine swimming in the performance arena was affected by water source (F1,123 = 5.93, P = 0.01, ηp2 = 0.05), but not by treatment or their interaction (P > 0.7). Fish that had been reared in water that had flowed over dead-degraded coral moved 13% further during their routine swimming compared to those that had been reared in water flowed over live coral (5.1 ± 0.2 vs 4.4 ± 0.2 m/120 s).
There was a positive relationship between distance of the focal fish to startle stimulus and response latency, but this differed significantly among treatments (F2,119 = 4.12, P = 0.019, ηp2 = 0.06), with the relationship being much weaker and shallower for fish exposed to dottybacks (r2 = 0.07, n = 40) compared to the other two treatments (control r2 = 0.37, n = 47; goby r2 = 0.47, n = 41).
Average and maximum response speed were positively correlated with length, so SL was used as a significant covariate in the analyses of these variables. Average response speed was affected by treatment (F2,121 = 5.38, P = 0.006, ηp2 = 0.08; Fig. 4), but not by water source or their interaction (P > 0.14). Response speed was greater for fish with a history of exposure to gobies than either dottyback or nil controls (Fig. 4). The relationship between maximum speed and standard length differed among treatments (F2,119 = 3.79, P = 0.02, ηp2 = 0.06), and once corrected for the effect of the covariate there was a trend towards an effect of treatment on maximum speed (F2,119 = 3.17, P = 0.046, ηp2 = 0.05), with those fish exposed to gobies having higher maximum speeds compared to the other two treatments.
Figure 4.
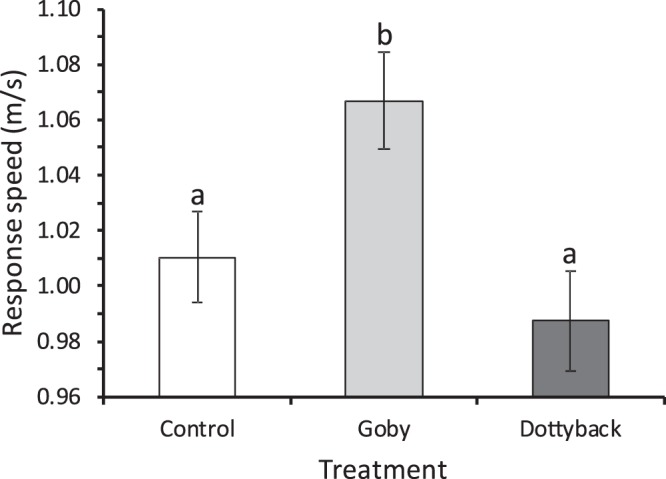
Response speed (mean ± SE) of juvenile Pomacentrus amboinensis to a drop stimulus when reared under three treatment levels of adult fish: a blank control, an omnivorous goby, or a predatory dottyback. Values are corrected for their covariation with standard length. Letters above bars represent Fisher’s LSD groupings of means. N = 41, 47, 40.
Discussion
The widespread and increasing degradation of coral reefs has drawn attention to the importance of identifying the mechanisms that make one species more resilient to habitat change than others41,42, as it has for other ecosystems such as forests43. This requires detailed information about the interactions between individuals within the context of their environment, and the potential for phenotypic plasticity to aid in their adaption to new environmental conditions. The current experiment is one of the first studies to explore how the changing nature of an ecosystem may affect predator-induced trait-mediated effects on body shape, behaviour and performance. Our findings show that prey attributes were flexible to predator presence and environmental context, but the overall effect of degraded water on trait-mediated predator effects was weaker than predicted. While there was a relatively minor impact of habitat degradation compared to the effect of predator cues, these modifications to the physiology of early settlement-stage individuals may be important in influencing life-history and survival trajectories at this important life-history bottleneck44,45 and on subsequent life stages46.
Water from degraded habitat did affect some aspects of the morphology and behaviour of fish, but in general this effect was independent of (i.e., did not interact with) whether fish were exposed to predators or not. The maximum diameter of the eye was larger for fish from live coral water compared to those reared in dead coral water. Since smaller eyes tend to have lower visual acuity47, smaller eyes may represent a cost to fish in degraded water48. These findings meant that damselfish reared in dead-degraded coral water had larger ocellus sizes relative to their eyes compared to fish reared in water from live coral. Interestingly, the effect of degraded water on ocellus size had half the effect size of the effect of predators on relative ocellus size, which caused predator-exposed fish to have larger ocellus relative to the true eye in keeping with a previous study23. Having a proportionally larger false eyespot compared to the true eye may be an advantage as it may attract strikes to the tail rather than the vulnerable head region, as has been shown by birds preying on butterflies with wing eyespots49. Predators anticipate the direction prey will move after an attack is initiated, and an eyespot may lead predators to misjudge the prey’s centre of mass50. A prey attacked at the caudal area can also escape and survive51, however an attack on the head would damage vital parts reducing the likelihood of survival. McPhail (1977) demonstrated that caudal spots in a freshwater characid fish (Hyphessobrycon panamensis) deflect the aim of a predatory freshwater barracuda (Ctenolucius beani). McPhail found that more predators focused their attacks on the caudal area of prey fish that had an artificial caudal spot drawn on compared to fish with no spot. Prey with these artificial eyespots escaped predators more frequently than fish with no eyespots, demonstrating a clear advantage. The presence of strongly selective predation52, and the phenotypic plasticity displayed by juvenile fishes generally (e.g.53,54), suggests that the benefits of developing a larger relative false eyespot outweighs the costs of having a smaller true eye, though field test will be needed to support this assertion.
Fish reared in degraded coral water moved 13% more during routine activity assessments compared to fish from healthy coral water. Previous research suggests that whether greater movement is advantageous depends on the life stage of the individual and knowledge of the specific risks associated with the settlement site55. Marine organisms with complex life histories that settle after a pelagic larval phase, be they invertebrates or vertebrates like fishes, often lack information on the identity of predators in their specific settlement site56. Research on fishes suggest that they rapidly obtain information on local risks through their own interactions with the community (e.g.39) and through the publically available information57. One study that quantified the space use of newly settled fish in the field found that survival on habitat patches was higher for bolder, more active fish, but was negatively related to how far they were willing to stray from habitat55. Previous research examining the behaviour of fishes on habitat patches found that fish on dead-degraded coral are more active and stay further away from the potential shelter31,58. The inability of many fishes to use alarm odours to assess and learn about risk may mean that fish on dead coral may be less conservative in their space use because they are unable to judge risk effectively. The problem they face is the impossibility of knowing their risk judgement has been impaired because, unlike senses such as vision, they have no way of knowing that they are not detecting an important odour. Such risky behaviour has been shown to lead to 75% higher mortality rates of juvenile fish over the first 48 h after settlement31 and may lead fish on degraded habitats to have higher mortality (e.g.33,59).
Regardless of the effects of whether the nursery habitat was live or dead-degraded coral, our findings show strong effects of the presence of predators on body shape, pigmentation, behaviour and performance in line with the limited studies that have explored these trait-mediated predator effects on fishes23,24. Studies of other organisms have found that non-consumptive predator-induced morphological change occurs in diverse taxa including amphibian60, gastropods61 and ciliates62. In the present experiments, fish exposed to predators grew longer compared to fish exposed to omnivorous gobies or the blank control. Being larger than average at a specific developmental stage is commonly advantageous due to all the aspect of performance that correlate with length63,64. The current experiment found aspects of performance such routine activity, maximum and mean escape speed scaled positively with length, emphasizing the importance of size. Many studies have found that predators select for the smaller members of a prey group65, and numerous studies within the current system demonstrate a strong and ongoing advantage to being larger than average in the early juvenile stage66,67.
Our study also found that body depth was also promoted by exposure to predators, in support of previous studies. The gape-limited nature of many aquatic predators means that one of the key relationships underlying catch success may be the width of a predator’s mouth compared to prey body depth67–69. Our finding of the development of a deeper body when in close proximity with a predator matches the findings of Lönnstedt et al.’s (2013) study of the same species, and the response of the crucian carp to predatory pike cues24. In a laboratory experiment, Chivers, et al.25 found that the relative body depth of the crucian carp was dynamic and could be rapidly altered though exposure to olfactory cues. This and the current experiment emphasize the dynamic nature of morphological defenses to the non-consumptive effects of predators.
Being exposed to a predator affected aspects of body length and growth compared to blank controls. Having the smell and sight of a known predator affected growth in length and weight, but there was an interaction between water source and treatment when length-standardised weight (i.e., bulk) was considered. Fish exposed to predators in live coral water gained more body mass than other treatment combinations. Given that all fish were provided the same amount of food throughout the experiment, this suggest that the efficiency of food conversion to tissue may have been higher for predator-exposed fish in live coral water compared to those in water from dead-degraded coral. Although the mechanism that would support this is unclear, it is likely related to the stress hormone cortisol. Middlemis et al.70 found that tadpoles (Rana sylvatica) that were exposed to predators had higher cortisol, which directly induced substantial growth of the tail compared to controls not exposed to predators. Our findings contrast with a number of other studies that have examined the influence of predator presence on fish growth71. For instance, Fu et al.72 exposed juvenile Chinese bream (Parabramis pekinensis) prey to predatory southern catfish (Silurus meridionalis) and found reduced growth and higher metabolism in prey groups with predators. Differences in results among studies may be due to differences in the life stage of fish examined, with other studies examining late-stage juveniles. The current study was initiated when individuals were newly metamorphosed; a life stage previously shown to be highly flexible in morphology and behaviour (e.g.22,23,55). Clearly, the physiological mechanisms underlying this flexibility and how it was affected by water from degraded corals warrant further research.
Fish reared with predators tended to have a uniformly fast escape response latency, which differed compared to the other treatments, where there was a strong relationship between distance to the startle stimulus and latency. This shows fish that had been exposed to predators were more responsive and responded more quickly. Fish have recently been shown to exhibit a context-dependent escape response, such that they optimize when and how fast they respond to a startle stimulus (such as a predator strike) dependent on how highly they rank the threat40,73,74. It appears that predator-exposed fish are primed to burst regardless of the distance to the threat, while fish reared under more benign conditions had an optimized response. Interestingly, once the fast-start had been initiated the goby-reared fish had better performance once they had initiated a C-start. Given that the speed at which fish respond to a predator (i.e., latency) is one of the best predictors of survival in the field64, it appears likely that the priming that comes with being forewarned of the presence of a predator may lead to higher survival.
Given the findings of previous research on the influence of degraded water on the efficacy of alarm odours to label threats and assess risk, it was surprising that the present study found few interactions between water source and predator presence. Laboratory and field studies show that our focal species, like many fishes29, is unable to use chemical alarm odours to learn predator identity or update information in the presence of small amounts of water from dead-degraded coral32, or even on a live coral patch surrounded by degraded habitat33. Being on thermally bleached or dead corals also changes the space use, activity and interactions of fishes59. Given the ability of cues from degraded coral to modify the availability of cues and behaviour it was expected that such water would also have a major impact on trait-mediated predator effects. While the damselfish may not have initially recognized the dottybacks as predators, it is likely that the close proximity of the predator to prey, and the strikes of predators at the Perspex dividers may have elevated the ranking of dottybacks on the threat gradient75. Other non-olfactory learning mechanisms, such as visual strikes, have likely filled the information void that would usually be received from olfactory sources76. While research suggests that some form of sensory compensation may occur when visual cues are reduced (through turbulence or in darkness77;), this has not been demonstrated for olfactory cues because of the uncertain meaning of an absence of smell. Clearly however, the present study demonstrates that there is substantial compensation for documented loss of the olfactory threat information in water from dead-degraded habitats27. This information allows the trait-mediated predator effects to be fully demonstrated in degraded water. It remains to be determined how they achieve the compensation for the loss of the important associative-learning mechanism of predator learning and reinforcement, and when the compensation occurs to allow the development of trait-mediated predator effects. Understanding these mechanisms will be key to determining how species survive under environmental adversity and how species’ resilience is promoted.
Supplementary information
Details on the study system, animal welfare, tank setup and morphometrics
Acknowledgements
We are grateful to C. Hemingson and A. Giffin for their amazing support running the experiment and keeping all ‘hotel’ guests fed, clean and happy.
Author contributions
M.I.M. and M.M.P. conceived the ideas and designed methodology; M.M.P. and E.F. collected the data; M.I.M. analysed the data; M.I.M. wrote the initial draft of the manuscript and all co-authors revised the manuscript for submission.
Data availability
Data is accessible from the Tropical Data Hub (10.25903/5cb82367e7f39).
Competing interests
The authors declare no competing interests.
Footnotes
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
is available for this paper at 10.1038/s41598-019-51798-2.
References
- 1.Haddad NM, et al. Habitat fragmentation and its lasting impact on Earth’s ecosystems. Science Advances. 2015;1:e1500052. doi: 10.1126/sciadv.1500052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Watson JE, et al. Catastrophic declines in wilderness areas undermine global environment targets. Curr. Biol. 2016;26:1–6. doi: 10.1016/j.cub.2016.08.049. [DOI] [PubMed] [Google Scholar]
- 3.Betts MG, et al. Global forest loss disproportionately erodes biodiversity in intact landscapes. Nature. 2017;547:441. doi: 10.1038/nature23285. [DOI] [PubMed] [Google Scholar]
- 4.Pimm SL, Jenkins CN, Li BV. How to protect half of Earth to ensure it protects sufficient biodiversity. Science Advances. 2018;4:eaat2616. doi: 10.1126/sciadv.aat2616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Pankhurst NW. The endocrinology of stress in fish: an environmental perspective. General & Comparative Endocrinology. 2011;170:265–275. doi: 10.1016/j.ygcen.2010.07.017. [DOI] [PubMed] [Google Scholar]
- 6.Busch DS, Hayward LS. Stress in a conservation context: A discussion of glucocorticoid actions and how levels change with conservation-relevant variables. Biol. Conserv. 2009;142:2844–2853. doi: 10.1016/j.biocon.2009.08.013. [DOI] [Google Scholar]
- 7.Homyack JA. Evaluating habitat quality of vertebrates using conservation physiology tools. Wildl. Res. 2010;37:332–342. doi: 10.1071/wr08093. [DOI] [Google Scholar]
- 8.Cooke SJ, et al. Conservation physiology across scales: insights from the marine realm. Conservation Physiology. 2014;2:15. doi: 10.1093/conphys/cou024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Hughes TP, et al. Global warming transforms coral reef assemblages. Nature. 2018;556:492–497. doi: 10.1038/s41586-018-0041-2. [DOI] [PubMed] [Google Scholar]
- 10.Chace JF, Walsh JJ. Urban effects on native avifauna: a review. Landscape Urban Plann. 2006;74:46–69. doi: 10.1016/j.landurbplan.2004.08.007. [DOI] [Google Scholar]
- 11.Metcalfe NB. The effects of habitat on the vigilance of shorebirds: is visibility important? Anim. Behav. 1984;32:981–985. doi: 10.1016/S0003-3472(84)80210-9. [DOI] [Google Scholar]
- 12.Lima SL. Distance to cover, visual obstructions, and vigilance in house sparrows. Behaviour. 1987;102:231. doi: 10.1163/156853986X00144. [DOI] [Google Scholar]
- 13.Endler JA. Signals, signal conditions, and the direction of evolution. Am. Nat. 1992;139:S125–S153. doi: 10.1086/285308. [DOI] [Google Scholar]
- 14.Weissburg M, Smee DL, Ferner MC. The sensory ecology of nonconsumptive predator effects. Am. Nat. 2014;184:141–157. doi: 10.1086/676644. [DOI] [PubMed] [Google Scholar]
- 15.Andruskiw M, Fryxell JM, Thompson ID, Baker JA. Habitat-mediated variation in predation risk by the American marten. Ecology. 2008;89:2273–2280. doi: 10.1890/07-1428.1. [DOI] [PubMed] [Google Scholar]
- 16.Smith, R. J. F. In Behavioural ecology of teleost fishes (ed. Godin, J. J.) 163–190 (Oxford University Press, 1997).
- 17.Schmitz OJ, Krivan V, Ovadia O. Trophic cascades: the primacy of trait-mediated indirect interactions. Ecol. Lett. 2004;7:153–163. doi: 10.1111/j.1461-0248.2003.00560.x. [DOI] [Google Scholar]
- 18.Salice CJ, Plautz SC. Predator-induced defences in offspring of laboratory and wild-caught snails: prey history impacts prey response. Evol. Ecol. Res. 2011;13:373–386. [Google Scholar]
- 19.Palacios MM, Warren DT, McCormick MI. Sensory cues of a top-predator indirectly control a reef fish mesopredator. Oikos. 2016;125:201–209. doi: 10.1111/oik.02116. [DOI] [Google Scholar]
- 20.Haggerty MB, Anderson TW, Long JD. Fish predators reduce kelp frond loss via a trait-mediated trophic cascade. Ecology. 2018;99:1574–1583. doi: 10.1002/ecy.2380. [DOI] [PubMed] [Google Scholar]
- 21.Chivers DP, et al. Predator-induced life history changes in amphibians: egg predation induces hatching. Oikos. 2001;92:135–142. doi: 10.1034/j.1600-0706.2001.920116.x. [DOI] [Google Scholar]
- 22.Ferrari MCO, et al. Living in a risky world: the onset and ontogeny of an integrated antipredator phenotype in a coral reef fish. Scientific Reports. 2015;5:15537. doi: 10.1038/srep15537. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Lönnstedt OM, McCormick MI, Chivers DP. Predator-induced changes in the growth of eyeballs and false eyespots. Scientific Reports. 2013;3:2259. doi: 10.1038/srep02259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Bronmark C, Miner JG. Predator induced phenotypic change in body morphology in the crucian carp. Science. 1992;258:1348–1350. doi: 10.1126/science.258.5086.1348. [DOI] [PubMed] [Google Scholar]
- 25.Chivers DP, Zhao X, Brown GE, Marchant TA, Ferrari MCO. Predator-induced changes in morphology of a prey fish: the effects of food level and temporal frequency of predation risk. Evol. Ecol. 2008;22:561–574. doi: 10.1007/s10682-007-9182-8. [DOI] [Google Scholar]
- 26.Domenici P, Turesson H, Brodersen J, Bronmark C. Predator-induced morphology enhances escape locomotion in crucian carp. Proceedings of the Royal Society B-Biological Sciences. 2008;275:195–201. doi: 10.1098/rspb.2007.1088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.McCormick MI, Barry R, Allan BJM. Algae associated with coral degradation affects risk assessment in coral reef fishes. Scientific Reports. 2017;7:16937. doi: 10.1038/s41598-017-17197-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Ferrari MCO, Wisenden BD, Chivers DP. Chemical ecology of predator–prey interactions in aquatic ecosystems: a review and prospectus. Canadian Journal of Zoology. 2010;88:698–724. doi: 10.1139/z10-029. [DOI] [Google Scholar]
- 29.Ferrari MCO, McCormick MI, Allan BJM, Chivers DP. Not equal in the face of habitat change: closely related fishes differ in their ability to use predation-related information in degraded coral. Proceedings of the Royal Society B. 2017;284:20162758. doi: 10.1098/rspb.2016.2758. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.McCormick MI. Behaviourally mediated phenotypic selection in a disturbed coral reef environment. PLoS One. 2009;4:e7096. doi: 10.1371/journal.pone.0007096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Lönnstedt OM, McCormick MI, Chivers DP, Ferrari MCO. Habitat degradation is threatening reef replenishment by making fish fearless. J. Anim. Ecol. 2014;83:1178–1185. doi: 10.1111/1365-2656.12209. [DOI] [PubMed] [Google Scholar]
- 32.Lönnstedt OM, McCormick MI, Chivers DP. Degraded environments alter prey risk assessment. Ecology and Evolution. 2013;3:38–47. doi: 10.1002/ece3.388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.McCormick MI, Lönnstedt OM. Disrupted learning: habitat degradation impairs crucial antipredator responses in naïve prey. Proceedings of the Royal Society B. 2016;283:20160441. doi: 10.1098/rspb.2016.0441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.McCormick MI, Weaver CJ. It pays to be pushy: intracohort interference competition between two reef fishes. PLoS One. 2012;7:e42590. doi: 10.1371/journal.pone.0042590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Feeney WE, et al. High rate of prey consumption in a small predatory fish on coral reefs. Coral Reefs. 2012;31:909–918. doi: 10.1007/s00338-012-0894-z. [DOI] [Google Scholar]
- 36.Hernaman V, Probert P, Robbins W. Trophic ecology of coral reef gobies: interspecific, ontogenetic, and seasonal comparison of diet and feeding intensity. Mar. Biol. 2009;156:317–330. doi: 10.1007/s00227-008-1085-x. [DOI] [Google Scholar]
- 37.Natt M, Lönnstedt OM, McCormick MI. Coral reef fish predator maintains olfactory acuity in degraded coral habitats. PLoS ONE. 2017;12:e0179300. doi: 10.1371/journal.pone.0179300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Chivers DP, McCormick MI, Allan BJM, Ferrari MCO. Risk assessment and predator learning in a changing world: understanding the impacts of coral reef degradation. Scientific Reports. 2016;6:32542. doi: 10.1038/srep32542. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Lönnstedt OM, McCormick MI, Meekan MG, Ferrari MCO, Chivers DP. Learn and live: the role of predator experience in influencing prey behaviour and survival. Proceedings of the Royal Society London B. 2012;279:2091–2098. doi: 10.1098/rspb.2011.2516. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.McCormick M, Allan B. Interspecific differences in how habitat degradation affects escape response. Scientific Reports. 2017;7:426. doi: 10.1038/s41598-017-00521-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Keith SA, et al. Synchronous behavioural shifts in reef fishes linked to mass coral bleaching. Nature Climate Change. 2018;8:986–991. doi: 10.1038/s41558-018-0314-7. [DOI] [Google Scholar]
- 42.Wismer S, Tebbett SB, Streit RP, Bellwood DR. Spatial mismatch in fish and coral loss following 2016 mass coral bleaching. Sci. Total Environ. 2019;650:1487–1498. doi: 10.1016/j.scitotenv.2018.09.114. [DOI] [PubMed] [Google Scholar]
- 43.Stork NE, et al. Vulnerability and resilience of tropical forest species to land-use change. Conserv. Biol. 2009;23:1438–1447. doi: 10.1111/j.1523-1739.2009.01335.x. [DOI] [PubMed] [Google Scholar]
- 44.Wilbur HM. Complex life cycles. Ann. Rev. Ecol. Syst. 1980;11:67–93. doi: 10.1146/annurev.es.11.110180.000435. [DOI] [Google Scholar]
- 45.Almany GR, Webster MS. The predation gauntlet: early post-settlement mortality in coral reef fishes. Coral Reefs. 2006;25:19–22. doi: 10.1007/s00338-005-0044-y. [DOI] [Google Scholar]
- 46.O’Connor CM, Norris DR, Crossin GT, Cooke SJ. Biological carryover effects: linking common concepts and mechanisms in ecology and evolution. Ecosphere. 2014;5:1–11. doi: 10.1890/ES13-00182.1. [DOI] [Google Scholar]
- 47.Caves EM, Sutton TT, Johnsen S. Visual acuity in ray-finned fishes correlates with eye size and habitat. J. Exp. Biol. 2017;220:1586–1596. doi: 10.1242/jeb.151183. [DOI] [PubMed] [Google Scholar]
- 48.Corral-López A, Garate-Olaizola M, Buechel SD, Kolm N, Kotrschal A. On the role of body size, brain size, and eye size in visual acuity. Behav. Ecol. Sociobiol. 2017;71:179. doi: 10.1007/s00265-017-2408-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Wourms MK, Wasserman FE. Butterfly wing markings are more advantageous during handling than during the initial strike of an avian predator. Evolution. 1985;39:845–851. doi: 10.1111/j.1558-5646.1985.tb00426.x. [DOI] [PubMed] [Google Scholar]
- 50.Ruxton, G. D., Sherratt, T. N. & Speed, M. P. Avoiding attack: the evolutionary ecology of crypsis, warning signals, and mimicry. (Oxford University Press, 2004).
- 51.McPhail JD. A possible function of the caudal spot in characid fishes. Canadian Journal of Zoology. 1977;55:1063–1066. doi: 10.1139/z77-136. [DOI] [Google Scholar]
- 52.Perez KO, Munch SB. Extreme selection on size in the early lives of fish. Evolution. 2010;64:2450–2457. doi: 10.1111/j.1558-5646.2010.00994.x. [DOI] [PubMed] [Google Scholar]
- 53.Boel M, Brodersen J, Koed A, Baktoft H, Post DM. Incidence and phenotypic variation in alewife alter the ontogenetic trajectory of young-of-the-year largemouth bass. Oikos. 2018;127:1800–1811. doi: 10.1111/oik.05556. [DOI] [Google Scholar]
- 54.Raffini F, Meyer A. A comprehensive overview of the developmental basis and adaptive significance of a textbook polymorphism: head asymmetry in the cichlid fish Perissodus microlepis. Hydrobiologia. 2019;832:65–84. doi: 10.1007/s10750-018-3800-z. [DOI] [Google Scholar]
- 55.McCormick MI, Meekan MG. The importance of attitude: the influence of behaviour on survival at an ontogenetic boundary. Mar. Ecol. Prog. Ser. 2010;407:173–185. doi: 10.3354/meps08583. [DOI] [Google Scholar]
- 56.Vail A, McCormick MI. Metamorphosing reef fishes avoid predator scent when choosing a home. Biol. Lett. 2011;7:921–924. doi: 10.1098/rsbl.2011.0380. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Manassa RP, McCormick MI. Social learning improves survivorship at a life history transition. Oecologia. 2013;171:845–852. doi: 10.1007/s00442-012-2458-x. [DOI] [PubMed] [Google Scholar]
- 58.Bostrom-Einarsson L, Bonin MC, Munday PL, Jones GP. Loss of live coral compromises predator-avoidance behaviour in coral reef damselfish. Scientific Reports. 2018;8:7795. doi: 10.1038/s41598-018-26090-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.McCormick MI. Lethal effects of habitat degradation on fishes through changing competitive advantage. Proc Roy Soc Lond B. 2012;279:3899–3904. doi: 10.1098/rspb.2012.0854. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Benard MF, Fordyce JA. Are induced defenses costly? Consequences of predator-induced defenses in western toads, Bufo boreas. Ecology. 2003;84:68–78. doi: 10.1890/0012-9658(2003)084[0068:AIDCCO]2.0.CO;2. [DOI] [Google Scholar]
- 61.Trussell GC, Nicklin MO. Cue sensitivity, inducible defense, and trade-offs in a marine snail. Ecology. 2002;83:1635–1647. doi: 10.1890/0012-9658(2002)083[1635:CSIDAT]2.0.CO;2. [DOI] [Google Scholar]
- 62.Kuhlmann, H., Kusch, J. & Heckmann, K. In The ecology and evolution of inducible defenses (eds Tollrian, R. & D. Harvell, C.) 142–159 (Princeton University Press, 1999).
- 63.Dial KP, Greene E, Irschick DJ. Allometry of behavior. Trends Ecol. Evol. 2008;23:394–401. doi: 10.1016/j.tree.2008.03.005. [DOI] [PubMed] [Google Scholar]
- 64.McCormick MI, Fakan E, Allan BJM. Behavioural measures determine survivorship within the hierarchy of whole-organism phenotypic traits. Funct. Ecol. 2018;32:958–969. doi: 10.1111/1365-2435.13033. [DOI] [Google Scholar]
- 65.Sogard SM. Size-selective mortality in the juvenile stage of teleost fishes: a review. Bull. Mar. Sci. 1997;60:1129–1157. [Google Scholar]
- 66.Figueira WF, Booth DJ, Gregson MA. Selective mortality of a coral reef damselfish: role of predator-competitor synergisms. Oecologia. 2008;156:215–226. doi: 10.1007/s00442-008-0985-2. [DOI] [PubMed] [Google Scholar]
- 67.Holmes T, McCormick MI. Size-selectivity of predatory reef fish on juvenile prey. Mar. Ecol. Prog. Ser. 2010;399:273–283. doi: 10.3354/meps08337. [DOI] [Google Scholar]
- 68.Werner EE, Gilliam JF. The ontogenetic niche and species interactions in size-structured populations. Annu. Rev. Ecol. Syst. 1984;15:393–425. doi: 10.1146/annurev.es.15.110184.002141. [DOI] [Google Scholar]
- 69.Mihalitsis M, Bellwood DR. A morphological and functional basis for maximum prey size in piscivorous fishes. PloS one. 2017;12:e0184679. doi: 10.1371/journal.pone.0184679. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Middlemis Maher J, Werner Earl E, Denver Robert J. Stress hormones mediate predator-induced phenotypic plasticity in amphibian tadpoles. Proceedings of the Royal Society B: Biological Sciences. 2013;280:20123075. doi: 10.1098/rspb.2012.3075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Bourdeau PE, Johansson F. Predator-induced morphological defences as by-products of prey behaviour: a review and prospectus. Oikos. 2012;121:1175–1190. doi: 10.1111/j.1600-0706.2012.20235.x. [DOI] [Google Scholar]
- 72.Fu C, Fu SJ, Wu QY, Cao ZD. Predation threat modifies relationships between metabolism and behavioural traits but not their ecological relevance in Chinese bream. Mar. Freshwat. Behav. Physiol. 2017;50:329–344. doi: 10.1080/10236244.2017.1411158. [DOI] [Google Scholar]
- 73.Domenici P. Context-dependent variability in the components of fish escape response: integrating locomotor performance and behavior. Journal of Experimental Zoology. 2010;313A:59–79. doi: 10.1002/jez.580. [DOI] [PubMed] [Google Scholar]
- 74.Herbert-Read JE, Ward AJW, Sumpter DJT, Mann RP. Escape path complexity and its context dependency in Pacific blue-eyes (Pseudomugil signifer) J. Exp. Biol. 2017;220:2076–2081. doi: 10.1242/jeb.154534. [DOI] [PubMed] [Google Scholar]
- 75.Ferrari MCO, Sih A, Chivers DP. The paradox of risk allocation: a review and prospectus. Anim. Behav. 2009;78:579–585. doi: 10.1016/j.anbehav.2009.05.034. [DOI] [Google Scholar]
- 76.Hartman EJ, Abrahams MV. Sensory compensation and the detection of predators: the interaction between chemical and visual information. Proceedings of the Royal Society B. 2000;267:571–575. doi: 10.1098/rspb.2000.1039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Leahy SM, McCormick MI, Mitchell M, Ferrari MCO. To fear or to feed: the effects of turbidity on perception of risk by a marine fish. Biol. Lett. 2011;7:811–813. doi: 10.1098/rsbl.2011.0645. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Details on the study system, animal welfare, tank setup and morphometrics
Data Availability Statement
Data is accessible from the Tropical Data Hub (10.25903/5cb82367e7f39).