

Abstract
Background and Purpose
Both types of cannabinoid receptors—CB1 and CB2—regulate brain functions relating to addictive drug‐induced reward and relapse. CB1 receptor antagonists and CB2 receptor agonists have anti‐addiction efficacy, in animal models, against a broad range of addictive drugs. Δ9‐Tetrahydrocannabivarin (Δ9‐THCV)—a cannabis constituent—acts as a CB1 antagonist and a CB2 agonist. Δ8‐Tetrahydrocannabivarin (Δ8‐THCV) is a Δ9‐THCV analogue with similar combined CB1 antagonist/CB2 agonist properties.
Experimental Approach
We tested Δ8‐THCV in seven different rodent models relevant to nicotine dependence—nicotine self‐administration, cue‐triggered nicotine‐seeking behaviour following forced abstinence, nicotine‐triggered reinstatement of nicotine‐seeking behaviour, acquisition of nicotine‐induced conditioned place preference, anxiety‐like behaviour induced by nicotine withdrawal, somatic withdrawal signs induced by nicotine withdrawal, and hyperalgesia induced by nicotine withdrawal.
Key Results
Δ8‐THCV significantly attenuated intravenous nicotine self‐administration and both cue‐induced and nicotine‐induced relapse to nicotine‐seeking behaviour in rats. Δ8‐THCV also significantly attenuated nicotine‐induced conditioned place preference and nicotine withdrawal in mice.
Conclusions and Implications
We conclude that Δ8‐THCV may have therapeutic potential for the treatment of nicotine dependence. We also suggest that tetrahydrocannabivarins should be tested for possible anti‐addiction efficacy in a broader range of preclinical animal models, against other addictive drugs, and eventually in humans.
Abbreviations
- CPA
conditioned place avoidance
- CPP
conditioned place preference
- FR
fixed ratio
- MP
minipump
- SA
self‐administration
- Δ8‐THCV
Δ8‐tetrahydrocannabivarin
- Δ9‐THCV
Δ9‐tetrahydrocannabivarin
What is already known
CB1 receptor antagonists and CB2 receptor agonists each show anti‐addiction profiles in animal models.
What does this study add
Δ8‐tetrahydrocannabivarin, a combined CB1 antagonist/CB2 agonist, shows potent anti‐nicotine effects in seven different animal models.
What is the clinical significance
A new and potent anti‐nicotine pharmacotherapy for humans may evolve, based on medicinal cannabis.
1. INTRODUCTION
Tobacco smoking is the leading cause of preventable deaths worldwide (U.S. Department of Health and Human Services, 2010; World Health Organization, 2013) and is largely driven by the dependence‐producing properties of https://www.guidetopharmacology.org/GRAC/LigandDisplayForward?ligandId=2585 (Stolerman & Jarvis, 1995). Although several medications are available to aid smoking cessation, such as https://www.guidetopharmacology.org/GRAC/LigandDisplayForward?ligandId=7135, nicotine replacement, and https://www.guidetopharmacology.org/GRAC/LigandDisplayForward?ligandId=5459), high relapse to smoking is seen (Harmey, Griffin, & Kenny, 2012; Hughes, Peters, & Naud, 2008; Rose, 2009). Thus, new pharmacotherapeutic treatments are needed. Furthermore, with the increasing legalization of both “medical” and recreational marijuana (e.g., Government of Canada/Gouvernement du Canada, 2018; National Conference of State Legislatures, 2018), it is essential to learn which of the more than 400 biologically active chemicals in cannabis (Grotenhermen & Russo, 2002) have verifiable medicinal value and which do not.
The endocannabinoid system is involved in drug addiction—not only to cannabinoids but also to virtually all addictive drugs (Maldonado, Valverde, & Berrendero, 2006), including nicotine (Gamaleddin et al., 2015). https://www.guidetopharmacology.org/GRAC/ObjectDisplayForward?objectId=56 cannabinoid receptors (Matsuda, Lolait, Brownstein, Young, & Bonner, 1990) are widely expressed in brain, while the CB2 receptors (Howlett et al., 2002; Matsuda, 1997; Munro, Thomas, & Abu‐Shaar, 1993) are widely expressed in the peripheral immune system. CB1 receptors regulate the dopaminergic reward system—mediating addictive drug reward and relapse to drug use after successful abstinence (Gardner, 2002; Gardner, 2005). https://www.guidetopharmacology.org/GRAC/ObjectDisplayForward?objectId=57 2 receptors are now known to also be expressed in brain (Van Sickle et al., 2005; albeit in much lower density than CB1 receptors), to regulate dopaminergic neuronal function (Zhang et al., 2014), and to mediate addictive drug‐seeking behaviours (Jordan & Xi, 2019; Manzanares et al., 2018; Xi et al., 2011). CB1 receptor antagonists have anti‐addiction efficacy in animal models, against a broad range of addictive substances (Cohen, Kodas, & Griebel, 2005; De Vries et al., 2001; Lupica, Riegel, & Hoffman, 2004; Maldonado et al., 2006; Tanda & Goldberg, 2003; Xi et al., 2006; Xi et al., 2008). CB2 agonists have similar anti‐addiction efficacy in animal models (Delis et al., 2017; Jordan & Xi, 2019; Manzanares et al., 2018; Navarrete, García‐Gutiérrez, & Manzanares, 2018; Xi et al., 2011; Zhang et al., 2014).
Δ9 https://www.guidetopharmacology.org/GRAC/LigandDisplayForward?ligandId=6418 (Δ9‐THCV)—a cannabis‐derived phytocannabinoid (Gill, Paton, & Pertwee, 1970)—has CB1 antagonist action combined with CB2 agonist action (Bolognini et al., 2010; McPartland, Duncan, Di Marzo, & Pertwee, 2015; Pertwee, 2008). Δ8‐Tetrahydrocannabivarin (Δ8‐THCV) is a synthetic, more stable, and easier‐to‐synthesize analogue of Δ9‐THCV with a similar pharmacological profile of combined CB1 antagonist and CB2 agonist action (Bátkai et al., 2012).
Therefore, in the present study, we have investigated the possible anti‐nicotine efficacy of Δ8‐THCV in seven different preclinical animal (rodent) models relevant to nicotine addiction and dependence.
2. METHODS
2.1. Animals
All animal care and experimental procedures were conducted in accordance with the Guide for the Care and Use of Laboratory Animals, 8th Edition (National Research Council, 2011). All experiments using rats were approved by the Animal Care and Use Committee of the National Institute on Drug Abuse of the U.S. National Institutes of Health. All experiments using mice were approved by the Virginia Commonwealth University Institutional Animal Care and Use Committee. Animal studies are reported in compliance with the ARRIVE guidelines (Kilkenny, Browne, Cuthill, Emerson, & Altman, 2010) and with the recommendations made by the British Journal of Pharmacology.
For self‐administration and relapse experiments, adult male alcohol‐preferring (P) rats (RGD Cat# 2302666, https://doi.org/info:x-wiley/rrid/RRID:RGD2302666; Lumeng, Hawkins, & Li, 1977) were used, in view of our previous success with nicotine self‐administration in this rat strain (Wang et al., 2015). The rats were obtained from the Indiana University Medical Center, Indianapolis, IN, USA. All rats were housed individually in a climate‐controlled room under a 12‐hr light/dark cycle. For conditioned place preference (CPP) and nicotine‐withdrawal experiments, adult (9 weeks of age upon arrival) male drug‐naïve ICR (Institute of Cancer Research; https://doi.org/info:x-wiley/rrid/RRID:SCR_011417) mice (Harlan Laboratories, Indianapolis, IN, USA) were used. Mice were group‐housed (five per cage) in a climate‐controlled room on a 12‐hr light/dark cycle. Food and water were available ad libitum throughout the experiments.
2.2. Intravenous nicotine self‐administration
2.2.1. Surgery
Animals were prepared for intravenous nicotine self‐administration by surgical catheterization of the right external jugular vein. Each jugular catheter was constructed of microrenathane (Braintree Scientific, Braintree, MA, USA); catheterization was performed under sodium pentobarbital anaesthesia using standard aseptic surgical techniques as described previously (Xi et al., 2008; Xi et al., 2011). Each catheter ran subcutaneously to the top of the rodent skull, where it connected to a stainless steel cannula that was fixed to the skull with four stainless steel jeweller's screws (Small Parts, Miami Lakes, FL, USA) and dental acrylic cement. Each stainless steel cannula was fused to a screw‐on, screw‐off connector in such a manner as to allow rapid connection and disconnection to an infusion pump via tubing encased in a protective metal spring from the head‐mounted connector to the top of the experimental chamber. To help prevent clogging, catheters were flushed daily with a gentamicin–heparin saline solution (0.1 mg·ml−1 gentamicin and 30 IU·ml−1 heparin; ICN Biochemicals, Cleveland, OH, USA).
2.2.2. Self‐administration apparatus
Experiments were conducted in operant response test chambers (Med Associates, Georgia, VT, USA). Each test chamber had two levers: one active and one inactive. Depression of the active lever activated an infusion pump; depression of the inactive lever was counted but had no consequence. A cue light and a speaker were located 12 cm above the active lever. The house light was turned on at the start of each 3‐hr test session. Scheduling of experimental events and data collection was accomplished using Med Associates software (Med Associates, Georgia, VT, USA).
2.2.3. Self‐administration procedure
Animals were allowed 7 days to recover from surgery and were then initially trained to self‐administer nicotine (30 μg·kg−1 per infusion) under fixed‐ratio 1 (FR‐1) reinforcement. Each nicotine infusion delivered a volume of 0.08 ml over 5 s and was paired with presentation of a stimulus light and tone. Each self‐administration session lasted 3 hr. Reliable nicotine self‐administration was considered to have been achieved when the following criteria were met: (a) >10 nicotine infusions per 3‐hr session; (b) <20% variability in daily nicotine infusions across two consecutive sessions; and (c) an active/inactive lever‐press ratio exceeding 2:1. To confirm that the operant lever response was reinforced by nicotine, a switch between the active and inactive levers was conducted in a subset of animals, in which the previous nicotine‐paired active lever became inactive, while the previous inactive lever became active. After confirming reliable nicotine‐reinforced operant self‐administration behaviour, the effects of Δ8‐THCV (3 mg·kg−1 or 10 mg·kg−1) on nicotine self‐administration were evaluated.
2.2.4. Effects of Δ8‐THCV on nicotine self‐administration
Δ8‐THCV was administered 30 min prior to the start of nicotine self‐administration. After each test, animals received 3–5 days of self‐administration of nicotine alone until stable self‐administration was re‐established. Δ8‐THCV was administered by intraperitoneal injection. The order of testing for the two doses of Δ8‐THCV was counterbalanced.
2.3. Relapse to nicotine‐seeking after a 14‐day forced abstinence period
Relapse to drug‐seeking behaviour can be measured by many different animal models (Venniro, Caprioli, & Shaham, 2016). For the present work, we chose a variant of the “forced abstinence” model (Venniro et al., 2016), in which the ability of the environmental context plus the conditioned cues (lights and tones) previously associated with drug self‐administration to evoke drug‐seeking behaviour after a period of involuntary withdrawal from drug‐taking behaviour is measured. After stable intravenous nicotine self‐administration was achieved, the animals were returned to their home colony cages for a 14‐day period of behavioural and pharmacological withdrawal. There was no explicit behavioural extinction of the nicotine‐taking habit. After the 14‐day withdrawal period, each animal was returned to its self‐administration chamber and allowed access for 3 hr to the levers that formerly—upon being depressed—activated the pump that delivered intravenous nicotine. During this post‐withdrawal test day, depression of the active lever delivered saline and activated the conditioned cues (the light and tone previously paired with each nicotine infusion). Thus, context‐ and conditioned cue‐induced relapse to nicotine‐seeking behaviour was assessed. We then observed the effects of Δ8‐THCV (10 or 20 mg·kg−1 i.p.) or vehicle (5% Cremophor®) on this behaviour.
2.4. Nicotine‐triggered relapse to nicotine‐seeking behaviour using the reinstatement model
The reinstatement animal model of relapse to drug‐seeking behaviour differs from the forced abstinence model in that animals are deliberately behaviourally extinguished from their prior drug‐taking habit by (a) substitution of saline or vehicle for the addictive drug in the pump of the self‐administration apparatus and (b) the drug‐associated cue light and tone are turned off during the extinction period (e.g., Xi et al., 2006). This behavioural extinction is continued until the animals reach a criterion of non‐response on the active lever that previously activated the intravenous delivery of drug. In the present study, daily extinction sessions continued until lever pressing was <10 per 3 hr session for three consecutive days. Then animals were divided into three experimental groups, and reinstatement testing was begun 24 hr later. On the reinstatement test day, one group of animals received pretreatment with saline, a second group received 10 mg·kg−1 i.p. Δ8‐THCV pretreatment, and a third group received 20 mg·kg−1 Δ8‐THCV i.p. pretreatment—each 30 min prior to a priming (triggering) injection of nicotine (0.15 mg·kg−1, s.c.). Active lever presses were then recorded for 3 hr.
2.5. Nicotine‐induced CPP
2.5.1. CPP apparatus
The CPP apparatus (Med Associates, St. Albans, VT, USA) consisted of white‐ and black‐coloured chambers (20 × 20 × 20 cm each) with differing floor textures (white mesh versus black rod) to allow the animals to differentiate between the two environmental contexts on the basis of visual and tactile cues. These two place conditioning chambers were separated by a smaller intermediate grey compartment with a smooth polyvinyl chloride floor and partitions that could be raised to allow access from the intermediate grey chamber to the black and white chambers.
2.5.2. CPP procedure
An unbiased CPP procedure was used, as we previously described (Kota et al., 2007). On Day 1, animals were confined to the middle grey chamber for a 5‐min habituation period and then allowed to move freely between all three chambers for 15 min. Time spent in each chamber was recorded, and those data were used to populate groups of approximately equal bias in baseline chamber preference. Twenty‐minute CPP acquisition sessions occurred twice a day (Days 2–4). During conditioning sessions, animals were confined to one of the larger chambers. The saline groups received saline in one large chamber in the morning and saline in the other large chamber in the afternoon. The nicotine group received nicotine in one large chamber and saline in the other large chamber. For the nicotine‐treated groups, CPP conditioning began 5 min after nicotine administration. Treatments were counterbalanced to ensure that some animals received the drug and paired environmental stimuli in the morning while others received them in the afternoon. The nicotine‐paired chamber was randomized amongst all groups. Sessions were 7 hr apart and were conducted by the same investigator.
2.5.3. Effects of Δ8‐THCV on nicotine‐induced CPP
To determine the effect of Δ8‐THCV on nicotine place conditioning, separate cohorts were generated by pretreating with either vehicle (5% Cremophor®) or Δ8‐THCV (0.03, 0.3, 3, or 30 mg·kg−1) by subcutaneous administration 30 min before nicotine. Day 5 was the drug‐free test day, and the procedure was the same as on Day 1—animals were allowed to freely explore the apparatus after the 5‐min habituation period. Locomotor activity counts and time spent on each side of the CPP apparatus were recorded. Data are expressed as a preference score: time spent on the drug‐paired side minus time spent on the saline‐paired side. A positive number indicates a preference for the drug‐paired side; a negative number indicates an aversion to the drug‐paired side. A number at or near zero indicates no preference for either side.
2.5.4. Effects of Δ8‐THCV on animal activity as measured on CPP test day
To confirm our previous impressions that Δ8‐THCV does not alter locomotor activity of test animals, we measured activity counts (seconds) on the CPP test day—comparing the locomotor activity effects of Δ8‐THCV (at four different doses) to that of vehicle (5% Cremophor®).
2.6. Nicotine withdrawal
2.6.1. Induction of nicotine withdrawal
Osmotic minipumps (model 2000; Alzet Corporation, Cupertino, CA, USA) that delivered continual infusion of 24 mg·kg−1·day−1 s.c. nicotine or saline for 14 days were surgically implanted under isoflurane anaesthesia, as we previously described (Damaj et al., 2003). Nicotine withdrawal was induced by removing the osmotic minipumps after 14 days of continuous nicotine administration, a regimen that we have previously shown to produce a significant nicotine withdrawal syndrome (Damaj et al., 2003). No analgesic was given in conjunction with the minipump removal, as this would have interfered with hyperalgesia testing (see below). One day after minipump removal, animals were treated with either vehicle or Δ8‐THCV at 0.3 mg·kg−1 s.c.—the dose that was found to most completely block the development of nicotine CPP— and then tested for nicotine withdrawal starting 30 min after vehicle or Δ8‐THCV administration.
2.6.2. Measurement of nicotine withdrawal
Nicotine withdrawal was measured in three different ways—(a) measurement of withdrawal‐induced anxiety‐like behaviour, (b) measurement of somatic signs of nicotine withdrawal, and (c) withdrawal‐induced hyperalgesia. In each instance, all ratings of nicotine withdrawal were performed by an observer blinded to the experimental treatment. The specific testing sequence (anxiety‐like behaviour; somatic signs of withdrawal; hyperalgesia) was based on our prior determination that this order of testing reduced within‐group variability and produced the most consistent results (Jackson, Martin, Changeux, & Damaj, 2008).
2.6.3. Nicotine withdrawal‐induced anxiety‐like behaviour
Animals were first evaluated in the plus maze test for anxiety‐like behaviour over a 5‐min period, as we have previously described (Damaj et al., 2003). Time spent on the closed arms of the maze was interpreted as a measure of anxiety‐like behaviour (Campos, Fogaça, Aguiar, & Guimarães, 2013). The number of crossings between the open and closed arms was counted as a measure of locomotor activity.
2.6.4. Observation and rating of overt somatic signs of nicotine withdrawal
Immediately following the plus maze testing, animals were evaluated for the characteristic overt somatic signs of nicotine withdrawal (Kwilasz, Harris, & Vann, 2009)—paw and body tremors, head shakes, retrograde locomotion, jumps, curls, and ptosis—as we have previously described (Damaj et al., 2003)—for 20 min. The total number of somatic signs was tallied for each animal, and the mean number of somatic signs during the observation period was calculated for each group.
2.6.5. Nicotine withdrawal‐induced hyperalgesia
Hyperalgesia is a well‐recognized component of nicotine withdrawal (Schmidt, Tambeli, Gear, & Levine, 2001). In the present experiments, nicotine withdrawal‐induced hyperalgesia was evaluated using the hot plate pain assay immediately following the somatic sign observation period. Animals were placed into a 10‐cm‐wide glass cylinder atop a hot plate (Thermojust Apparatus, Richmond, VA, USA) that was maintained at 52°C. Latency to reaction time (primarily paw licking) was recorded.
2.7. Data and statistical analyses
The data and statistical analysis comply with the recommendations of the British Journal of Pharmacology on experimental design and analysis in pharmacology. Animal group sizes were chosen on the basis of extensive previous experience with the animal models used. No data points were excluded from the analysis in any experiment. Where variation in group size occurred, this was due to animals being dropped from the experiment due to obstruction or clogging of intravenous catheters. Data are expressed as means ± SEM for each group. All experimental data were analysed using one‐way or two‐way ANOVA (Prism 6; GraphPad Software, La Jolla, CA, USA). Where a significant difference amongst group means was revealed by ANOVA (P < .05), between‐group individual comparisons were analysed using the Student–Newman–Keuls post hoc multiple comparisons procedure (Kirk, 1982). Values of P = .05 or P > .05 were taken to indicate no statistically significant differences (N.S.) among or between sample means.
2.8. Materials
(−)‐Nicotine hydrogen tartrate, (−)‐1‐methyl‐2‐(3‐pyridyl)pyrrolidine (+)‐bitartrate, was purchased from Sigma‐Aldrich Inc. (St. Louis, MO, USA) and was dissolved in physiological saline. The nicotine solution pH was neutralized with sodium bicarbonate as needed. Freshly prepared solutions were used in all experiments. Nicotine doses are expressed as the free base of the drug. For experiments involving mice, nicotine was given at a volume of 10 ml·kg−1 s.c. For CPP experiments, nicotine was given in a 0.5 mg·kg−1 s.c. bolus because we previously found that this dose produced robust CPP in ICR mice (Kota, Martin, Robinson, & Damaj, 2007). For nicotine withdrawal studies, 24 mg·kg−1·day−1 nicotine or saline was continuously infused for 14 days using subcutaneous osmotic minipumps (model 2000; Alzet Corporation, Cupertino, CA, USA) that were surgically implanted under isoflurane anaesthesia. We had previously found that this chronic nicotine regimen produces a significant withdrawal syndrome upon abrupt cessation of nicotine (Damaj, Kao, & Martin, 2003). Δ8‐THCV was obtained from Organix Inc. (Woburn, MA, USA) and was dissolved in 5% polyethoxylated castor oil (Cremophor®; purchased from Sigma‐Aldrich). The doses of Δ8‐THCV were chosen based on pilot studies, indicating efficacy in each experiment without significant adverse effects such as sedation or locomotor impairment.
2.9. Nomenclature of targets and ligands
Key protein targets and ligands in this article are hyperlinked to corresponding entries in http://www.guidetopharmacology.org/, the common portal for data from the IUPHAR/BPS Guide to PHARMACOLOGY (Harding et al., 2018), and are permanently archived in the Concise Guide to PHARMACOLOGY 2017/18 (Alexander et al., 2017).
3. RESULTS
3.1. Δ8‐THCV inhibits intravenous nicotine self‐administration
Systemic administration of Δ8‐THCV significantly inhibited intravenous nicotine self‐administration—measured as active lever presses for intravenous nicotine infusions (Figure 1, central panel), or total numbers of intravenous nicotine infusions received (Figure 1, left panel). Δ8‐THCV had no effect on inactive lever‐pressing (Figure 1, right panel).
Figure 1.
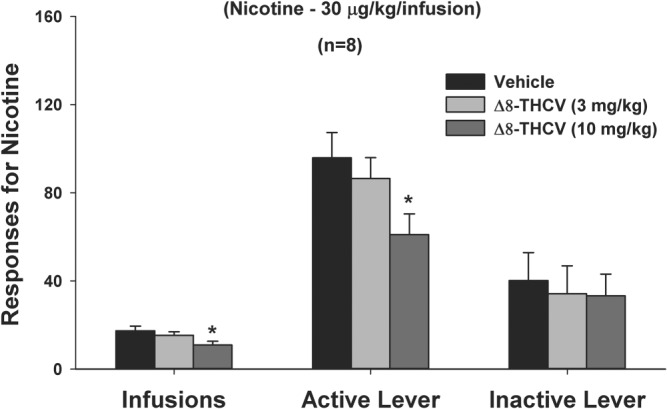
Effect of intraperitoneal administration of Δ8‐tetrahydrocannabivarin (Δ8‐THCV) on intravenous nicotine self‐administration. Δ8‐THCV significantly and dose‐dependently reduced intravenous nicotine self‐administration—measured as either total numbers of intravenous nicotine infusions received (left bars) or as active lever‐presses for intravenous nicotine infusions (central bars), while having no significant effect on lever‐pressing that resulted in no intravenous nicotine delivery (right bars). * P < .05, significantly different from vehicle (0 mg·kg−1 Δ8‐THCV), as determined by individual group comparisons. Sample size n = 8
3.2. Δ8‐THCV inhibits cue‐induced nicotine‐seeking after a 14‐day “drug holiday” from intravenous nicotine self‐administration
Δ8‐THCV significantly inhibited conditioned‐cue/context‐induced nicotine‐seeking behaviour—measured as active nicotine‐seeking lever presses after a 14‐day period of forced abstinence from nicotine‐taking behaviour (Figure 2, right panel). Active lever presses during the test session delivered only intravenous infusions of saline plus re‐exposure to the nicotine‐associated cue lights and tone. Δ8‐THCV's protective effect at 20 mg·kg−1 constituted a greater than 90% decrease in relapse to nicotine‐seeking.
Figure 2.
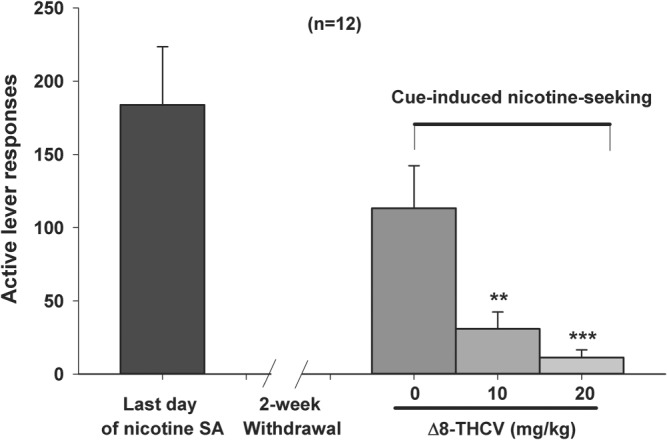
Effect of intraperitoneal administration of Δ8‐tetrahydrocannabivarin (Δ8‐THCV) on cue‐triggered nicotine‐seeking behaviour using the “forced abstinence” animal model of relapse to drug‐seeking. Δ8‐THCV significantly and dose‐dependently attenuated context‐triggered relapse to nicotine‐seeking behaviour after a two‐week period of nicotine withdrawal (right bars). * P < .05, significantly different from vehicle (0 mg·kg−1 Δ8‐THCV), as determined by individual group comparisons. Sample size n = 12
3.3. Δ8‐THCV inhibits nicotine‐triggered relapse to nicotine‐seeking in the “reinstatement” animal model of relapse
Δ8‐THCV dose‐dependently inhibited nicotine‐triggered relapse to nicotine‐seeking behaviour in the “reinstatement” (Venniro et al., 2016) model of relapse—measured as active nicotine‐seeking lever presses (Figure 3, right panel). Active lever presses during the test session delivered only vehicle. Δ8‐THCV's protective effect at 10 or 20 mg·kg−1 constituted a 70% decrease in relapse to nicotine‐seeking.
Figure 3.
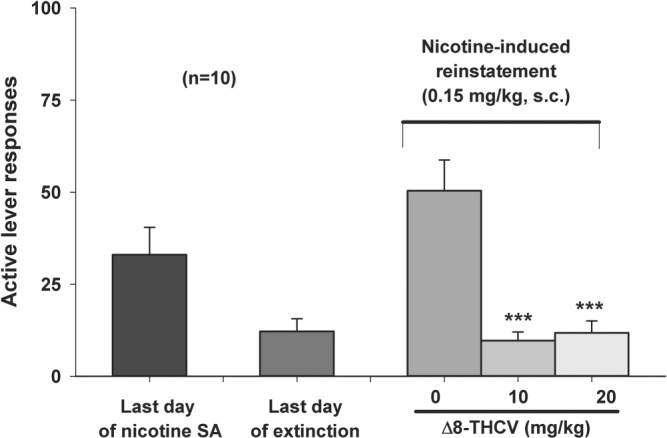
Effect of intraperitoneal administration of Δ8‐tetrahydrocannabivarin (Δ8‐THCV) on nicotine‐triggered relapse to nicotine‐seeking behaviour in animals behaviourally extinguished and (per force) pharmacologically detoxified from their prior nicotine‐taking behaviour, using the “reinstatement” animal model of relapse to drug‐seeking. Δ8‐THCV significantly and dose‐dependently reduced nicotine‐triggered relapse to nicotine‐seeking behaviour (right bars). * P < .05, significantly different from vehicle (0 mg·kg−1 Δ8‐THCV), as determined by individual group comparisons. Sample size n = 10
3.4. Δ8‐THCV inhibits context‐induced nicotine CPP
Δ8‐THCV—administered prior to nicotine on CPP conditioning days—dose‐dependently inhibited environmental context‐induced nicotine‐seeking behaviour on CPP test days—measured as time spent preferentially in the nicotine‐paired CPP test chambers (Figure 4). Two‐way ANOVA followed by Newman–Keuls multiple comparisons showed that nicotine produced a significant CPP and that there was a dose‐dependent reduction in nicotine‐paired context‐induced CPP on the CPP test day in those animals that had received Δ8‐THCV pretreatment prior to nicotine on their CPP conditioning days (Figure 4). Specifically, nicotine‐induced CPP was very robustly blocked by all doses of Δ8‐THCV. Δ8‐THCV had no effect—at any dose—on vehicle‐paired place preference (Figure 4); that is, Δ8‐THCV by itself produced neither a CPP nor a conditioned place avoidance (CPA).
Figure 4.
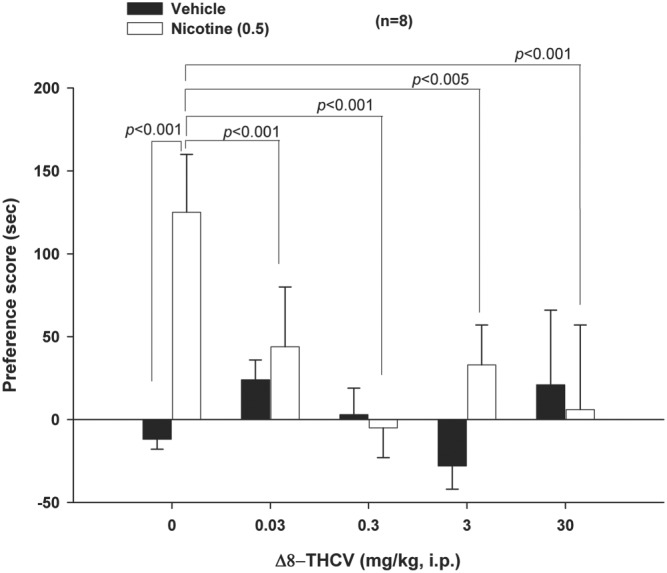
Effect of subcutaneous administration of Δ8‐tetrahydrocannabivarin (Δ8‐THCV) on acquisition of nicotine‐induced conditioned place preference (CPP). Δ8‐THCV, administered prior to nicotine during CPP conditioning days, significantly and dose‐dependently inhibited context‐induced nicotine‐seeking behaviour on CPP test days. Δ8‐THCV by itself was motivationally neutral—producing neither a CPP nor a conditioned place avoidance (CPA). * P < .05, significantly different as indicated. Sample size n = 8
3.5. Δ8‐THCV does not alter animal activity as measured on CPP test day
Animals treated with Δ8‐THCV (0.03 ‐ 30 mg·kg−1) did not display altered locomotor behaviour compared to their vehicle counterparts (see Table 1).
Table 1.
Δ8‐THCV did not alter locomotor activity on conditioned place preference test day
| Group | Locomotor activity counts | |
|---|---|---|
| Vehicle‐treated animals | Nicotine‐treated animals | |
| Vehicle | 1,451 ± 197 | 1,397 ± 132 |
| Δ8‐THCV (0.03 mg·kg−1) | 1,445 ± 134 | 1,362 ± 103 |
| Δ8‐THCV (0.3 mg·kg−1) | 1,404 ± 158 | 1,234 ± 127 |
| Δ8‐THCV (3 mg·kg−1) | 1,391 ± 168 | 1,305 ± 131 |
| Δ8‐THCV (30 mg·kg−1) | 1,293 ± 198 | 1,275 ± 158 |
Note. Animals pretreated with Δ8‐THCV did not show altered locomotor activity compared to vehicle‐pretreated animals. Numbers are presented as means ± SEM for n = 6–8 per group.
Abbreviation: Δ8‐THCV, Δ8‐tetrahydrocannabivarin.
3.6. Δ8‐THCV inhibits anxiety‐like signs of nicotine withdrawal
As noted above, rodents undergoing acute nicotine withdrawal display anxiety‐like behaviour, which is easily evaluated using a standard rodent plus maze test (Damaj et al., 2003). Time spent on the open arms of the plus maze is interpreted as constituting anti‐anxiety‐like behaviour (Campos et al., 2013). As shown in Figure 5, animals undergoing nicotine withdrawal showed high levels of anxiety‐like behaviour in the plus maze. This nicotine‐withdrawal‐induced anxiety‐like behaviour was significantly ameliorated by 0.3 mg·kg−1 Δ8‐THCV. Moreover, Δ8‐THCV did not affect total crossings between arms (Table 2), suggesting that Δ8‐THCV's effect on nicotine‐withdrawal‐induced anxiety‐like behaviour was not due to motor effects. Importantly, Δ8‐THCV (0.3 mg·kg−1) did not alter behaviour in animals that were treated with saline instead of nicotine (Table 2).
Figure 5.
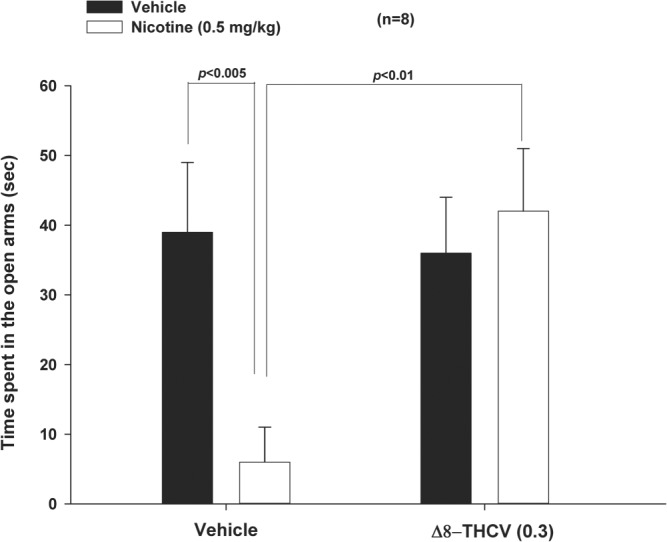
Effect of subcutaneous administration of Δ8‐tetrahydrocannabivarin (Δ8‐THCV) on nicotine‐withdrawal‐induced anxiety‐like behaviour. Animals undergoing nicotine withdrawal showed high levels of anxiety‐like behaviour in the plus maze, that is, significantly less time spent in the open arms of the maze, which was significantly ameliorated by 0.3 mg·kg−1 (s.c.) Δ8‐THCV. * P < .05, significantly different as indicated. Sample size n = 8
Table 2.
Δ8‐THCV did not alter number of arm crossings in the elevated plus maze
| Treatment | Arm crosses ± SEM |
|---|---|
| Saline MP, vehicle | 7.3 ± 0.8 |
| Saline MP, Δ8‐THCV (0.3 mg·kg−1) | 7.1 ± 1.2 |
| Nicotine MP, vehicle | 6.7 ± 1.3 |
| Nicotine MP, Δ8‐THCV (0.3 mg·kg−1) | 8.1 ± 1.8 |
Note. Animals undergoing spontaneous nicotine withdrawal were treated with vehicle or 0.3 mg·kg−1 Δ8‐THCV s.c., and number of crossings between arms of the plus maze was counted. Numbers are presented as means ± SEM for n = 6–8 per group.
Abbreviations: Δ8‐THCV, Δ8‐tetrahydrocannabivarin; MP, minipump.
3.7. Δ8‐THCV inhibits somatic signs of acute nicotine withdrawal
As also noted above, rodents undergoing acute nicotine withdrawal display a distinctive set of overt somatic withdrawal signs—including paw and body tremors, head shakes, retrograde locomotion, jumps, curls, and ptosis (Damaj et al., 2003; Kwilasz et al., 2009). In the present study, total number of somatic signs was tallied for each animal, and the mean number of somatic signs during the 20‐min observation period was calculated for each group. As shown in Figure 6, animals undergoing nicotine withdrawal showed high levels of overt somatic withdrawal signs, which was robustly ameliorated by 0.3 mg·kg−1 of Δ8‐THCV. Δ8‐THCV did not alter behaviour in animals that were treated with saline instead of nicotine. The specific somatic signs of nicotine withdrawal in the present experiment—and their counts—are shown in Table 3.
Figure 6.
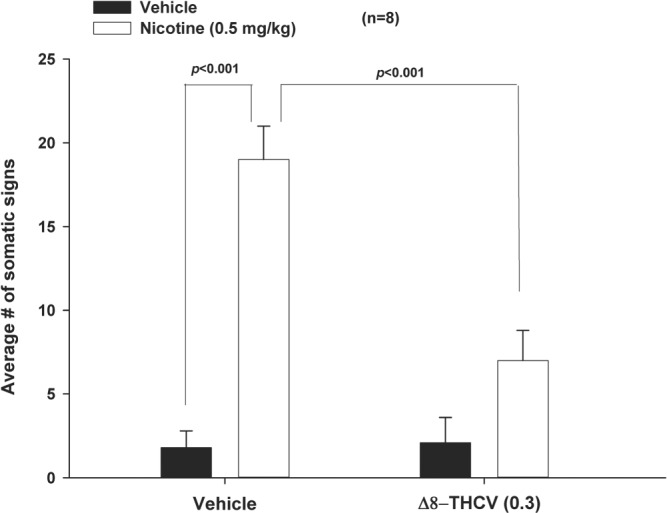
Effect of subcutaneous administration of Δ8‐tetrahydrocannabivarin (Δ8‐THCV) on the characteristic overt somatic signs of nicotine withdrawal. Animals in acute nicotine withdrawal showed high levels of overt somatic withdrawal signs, which were rated by an observer blind as to the treatments administered to the animals. These overt somatic withdrawal signs were then averaged for each animal, and a mean overall somatic withdrawal score compiled for each animal. A mean somatic withdrawal score was then computed for each group of animals. This mean somatic withdrawal score is indicated on the y axis of the figure as Average # of somatic signs. As can be seen, these averaged signs of somatic withdrawal from nicotine were significantly and robustly ameliorated by 0.3 mg·kg−1 (s.c.) of Δ8‐THCV. * P < .05, significantly different as indicated. Sample size n = 8
Table 3.
Effects of Δ8‐THCV on characteristic somatic signs of nicotine withdrawal.
| Signs | Individual withdrawal signs under vehicle, nicotine, and/or Δ8‐THCV | |||
|---|---|---|---|---|
| Veh/Veh | Veh/Δ8‐THCV | Nic/Veh | Nic/Δ8‐THCV | |
| Paw tremors | 1.3 ± 0.5 | 0.7 ± 0.4 | 10.9 ± 1.2 | 4.5 ± 1.2 |
| Head shakes | 0.3 ± 0.2 | 0 ± 0 | 3.9 ± 1.2 | 0.8 ± 0.3 |
| Backing | 0 ± 0 | 0 ± 0 | 1.3 ± 0.6 | 0.8 ± 0.4 |
| Body tremors | 0 ± 0 | 0 ± 0 | 2 ± 0.4 | 0.5 ± 0.3 |
| Others | 0 ± 0 | 0.8 ± 0.2 | 1 ± 0 | 0.8 ± 0.2 |
Note. Data are expressed as means ± SEM for n = 8 per group.
Abbreviation: Δ8‐THCV, Δ8‐tetrahydrocannabivarin.
3.8. Δ8‐THCV reverses nicotine withdrawal‐induced hyperalgesia
Nicotine withdrawal produced profound hyperalgesia—measured as time to painful response during a 20‐s test period, using the hot plate test (Figure 7). As shown, Δ8‐THCV totally reversed nicotine withdrawal‐induced hyperalgesia. Two‐way ANOVA followed by Newman–Keuls multiple comparisons indicated that nicotine‐withdrawal‐induced hyperalgesia was virtually abolished in animals treated with 0.3 mg·kg−1 Δ8‐THCV. Δ8‐THCV did not alter behaviour in animals that were treated with saline instead of nicotine.
Figure 7.
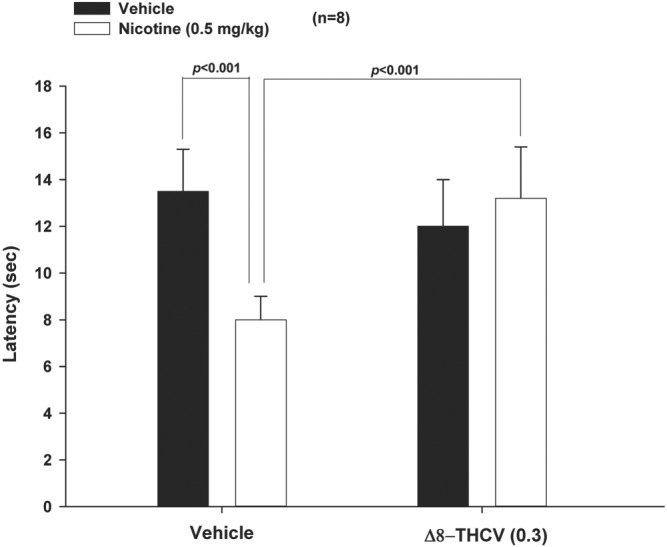
Effect of subcutaneous (s.c.) administration of Δ8‐tetrahydrocannabivarin (Δ8‐THCV) on nicotine‐withdrawal‐induced hyperalgesia, as measured by withdrawal latency (primarily paw‐licking) using a hot plate device. Animals undergoing nicotine withdrawal showed high levels of hyperalgesia in the hot plate test, that is, significantly shorter response latency to the pain induced by the hot plate. This nicotine‐withdrawal‐induced hyperalgesia was virtually abolished by 0.3 mg·kg−1 (s.c.) Δ8‐THCV. * P < .05, significantly different as indicated. Sample size n = 8
4. DISCUSSION
Δ8‐THCV and Δ9‐THCV are propyl homologues of Δ9‐tetrahydrocannabinol (Δ9‐THC), the principal psychoactive constituent of cannabis. This chemical change gives the THCVs different pharmacological profiles from that of Δ9‐THC. More specifically, like Δ9‐THC, Δ8‐THCV and Δ9‐THCV are effective displacers of the high‐potency CB1 receptor agonist https://www.guidetopharmacology.org/GRAC/LigandDisplayForward?ligandId=730 but, unlike Δ9‐THC, act as CB1 receptor antagonists in vitro as indicated by their ability to antagonize CP55940 in the GTPγS binding assay and the high‐potency CB1 receptor agonist https://www.guidetopharmacology.org/GRAC/LigandDisplayForward?ligandId=733 in the mouse vas deferens assay (Pertwee et al., 2007). Δ8‐THCV and Δ9‐THCV can also produce signs of CB1 receptor antagonism in vivo, as indicated by the ability of both of them to attenuate Δ9‐THC‐induced anti‐nociception and hypothermia at doses of 0.3, 1, and/or 3 mg·kg−1 i.v. and by the ability of Δ8‐THCV, although not Δ9‐THCV, to attenuate Δ9‐THC‐induced ring immobility at 0.3 and 3 mg·kg−1 i.v. (Pertwee et al., 2007).
Δ9‐THCV is found naturally in cannabis, sometimes in amounts exceeding 50% of total cannabinoids in some strains of cannabis from southern Africa, India, Nepal, and eastern Asia (ElSohly, Radwan, Gul, Chandra, & Galal, 2017; Hillig & Mahlberg, 2004; Turner, Hadley, & Fetterman, 1973). Currently, the THCVs are not scheduled as controlled or addictive substances by the U.S. Federal government nor the United Nations Convention on Psychotropic Substances and appear to be exceptionally safe in human use (Englund et al., 2016; Jadoon et al., 2016). As noted above, we and others have reported that CB1 receptor antagonists have remarkable anti‐addiction efficacy against a wide range of addictive drugs in a large number of preclinical animal models. As also noted above, we and others have reported that CB2 receptor agonists show similar anti‐addiction efficacy in a wide array of preclinical animal models (Jordan & Xi, 2019). Given that Δ8‐THCV and Δ9‐THCV combine CB1 antagonist action with CB2 agonist action, we have suggested that the THCVs may constitute a safe and non‐psychoactive class of potential anti‐addiction, anti‐craving, and anti‐relapse pharmacotherapies (Gardner, 2014). The present experiments appear to confirm that suggestion, at least for nicotine.
As noted above, CB1 receptor antagonists have anti‐addiction efficacy against a broad range of addictive substances in animal models (Cohen et al., 2005; De Vries et al., 2001; Lupica et al., 2004; Maldonado et al., 2006; Tanda & Goldberg, 2003; Xi et al., 2006; Xi et al., 2008), as well as potential anti‐obesity effects (Van Gaal et al., 2005). On the latter grounds, the CB1 receptor antagonist https://www.guidetopharmacology.org/GRAC/LigandDisplayForward?ligandId=743 (rimonabant) was approved by the European Commission in 2006 for treatment of overeating and obesity—especially in patients with associated risk factors such as type 2 diabetes or dyslipidaemia—and became available for prescription use in the United Kingdom in July 2006. By 2008, SR141716 was available in 56 countries. Although intended to control overeating and obesity, SR141716 was soon recognized to produce a highly significant (50%) increased rate of abstinence from smoking when compared to placebo in Phase III human clinical trials (Cahill & Ussher, 2011; Elrashidi & Ebbert, 2014). Unfortunately, SR141716 was also found to produce significant anxiety, depression, and suicidality in humans (Christensen, Kristensen, Bartels, Bliddal, & Astrup, 2007; Sam, Salem, & Ghatei, 2011). In October 2008, the European Medicines Agency recommended suspension of clinical use of SR141716 after concluding that its risks outweighed its benefits, and its approval was withdrawn by the European Commission in January 2009. This effectively terminated all anti‐addiction medication development based solely on CB1 receptor antagonism. However, the underlying mechanism(s) by which SR141716 produces anxiety, depression, and suicidality in humans has never been explored. The assumption has been that these effects result from CB1 receptor inverse agonism, but this has not been proven. In animal models, SR141716 by itself produces an anhedonic‐like effect—as assessed by electrical brain‐stimulation reward (He et al., 2019; Xi et al., 2008), in vivo brain microdialysis (Gardner, Gamaleddin, Manzanares Robles, & Rodrígues de Fonseca, 2013), and conditioned place aversion (Gardner et al., 2013). It also produces anxiety‐like effects and depressive‐like effects as shown in the elevated plus maze and forced swim test (Gueye et al., 2016). In the present experiments, Δ8‐THCV alone (i.e., when administered to animals given vehicle) did not produce pro‐anxiety‐like effects, anti‐anxiety‐like effects, CPP, or CPA.
Recent published studies indicate that CB2 receptor agonism produces potent anti‐addictive effects not dissimilar to those seen with CB1 receptor antagonism (Jordan & Xi, 2019; Manzanares et al., 2018). As noted above, activation of CB2 receptors by https://www.guidetopharmacology.org/GRAC/LigandDisplayForward?ligandId=747 inhibits cocaine self‐administration (Xi et al., 2011; Zhang et al., 2017; but see Adamczyk et al., 2012), cocaine‐induced CPP (Canseco‐Alba et al., 2018; Delis et al., 2017; Ignatowska‐Jankowska, Muldoon, Lichtman, & Damaj, 2013), and cocaine‐enhanced nucleus accumbens dopamine and locomotion in rodents (Delis et al., 2017; Xi et al., 2011). In contrast to these findings with cocaine, genetic deletion or pharmacological blockade (by https://www.guidetopharmacology.org/GRAC/LigandDisplayForward?ligandId=750 or https://www.guidetopharmacology.org/GRAC/LigandDisplayForward?ligandId=751) of CB2 receptors has been reported to attenuate nicotine‐induced CPP (Canseco‐Alba et al., 2018; Ignatowska‐Jankowska et al., 2013; Navarrete et al., 2013), nicotine self‐administration (Navarrete et al., 2013), and nicotine withdrawal symptoms (Navarrete et al., 2013; but see Ignatowska‐Jankowska et al., 2013). However, findings with various differing CB2 receptor agonists are conflicting. An early study indicated that the CB2 receptor agonist https://www.guidetopharmacology.org/GRAC/LigandDisplayForward?ligandId=3316, at low doses (1, 10 mg·kg−1, i.p.), failed to alter nicotine self‐administration or reinstatement of nicotine‐seeking behaviour (Gamaleddin, Zvonok, Makriyannis, Goldberg, & Le Foll, 2012), while another CB2 receptor agonist, O‐1966, when given in combination with a subthreshold dose of nicotine, elicited a CPP (Ignatowska‐Jankowska et al., 2013). In contrast, pretreatment with JWH133 blocked nicotine‐induced CPP (Canseco‐Alba et al., 2018) and Xie2‐64, a CB2 receptor inverse agonist, dose‐dependently inhibited nicotine‐enhanced optogenetic brain‐stimulation reward in rats and nicotine self‐administration in both rats and wild‐type mice, but not in CB2 receptor‐KO mice (Jordan et al., unpublished results). Among the possible reasons underlying such conflicting findings, one possibility is that CB2 receptors may play a different role in cocaine versus nicotine reward. More studies are required to understand the underlying mechanisms by which CB2 receptor agonism produces robust anti‐cocaine effects (e.g., Xi et al., 2011) and by which combined CB1 receptor antagonism/ CB2 receptor agonism produces the robust anti‐nicotine effects observed in the present study. Moreover, it will be important to seek pharmacological action(s) other than CB2 receptor agonism that may contribute to the anti‐nicotine effects produced by Δ8‐THCV in the present experiments. Further to the issue of differences between the present findings and those of Gamaleddin et al. (2012), we suggest that species (and possibly strain) differences may—in part—play a role, as species differences in splicing, expression, and brain distribution of CB2 genes and receptors have been found to alter the rewarding effects produced by drugs of abuse (see McPartland, Glass, & Pertwee, 2007; Liu et al., 2009; Zhang et al., 2015).
In summary, in the present work, we found strong (and in most instances, robust) anti‐nicotine‐dependence effects induced by Δ8‐THCV, in laboratory rodents using seven different preclinical animal models of nicotine dependence. We therefore suggest that the THCVs constitute a novel and possibly highly effective new class of anti‐addiction medications. We note that cannabis strains with very high levels of THCVs are currently available from cannabis dispensaries in California, Colorado, and the United Kingdom. We urge that (a) follow‐on experiments be carried out with Δ9‐THCV to confirm that Δ8‐THCV's remarkable anti‐addiction effects are also present in the phytocannabinoid analogue; (b) follow‐on experiments be carried out to determine what other addictive substances (opioids, alcohol, psychostimulants, etc.) may have their addictive properties altered by the THCVs; (c) experiments be undertaken to identify pharmacological actions of Δ8‐THCV and Δ9‐THCV other than CB1 receptor antagonism and CB2 receptor agonism that may contribute to the anti‐nicotine effects herein reported; (d) experiments be undertaken to rigorously determine whether Δ8‐THCV or Δ9‐THCV produce rimonabant‐like adverse effects such as depression, anxiety, or suicidality; (e) follow‐on studies be undertaken to determine whether the robust anti‐nicotine effects seen in the present experiments are replicated in both male and female animal subjects or at the human level; and (f) naturalistic field studies be carried out to determine whether regular users of high THCV‐containing strains of cannabis report decreased use and/or craving for such dependence‐producing drugs as nicotine, alcohol, or opioids.
CONFLICT OF INTEREST
The authors declare no conflicts of interest. Co‐authors Xi, Bi, Wang, and Gardner disclose that—during this research—they were salaried employees of the Intramural Research Program, National Institute on Drug Abuse, U.S. Public Health Service. Co‐author Pertwee discloses that, in addition to being Emeritus Professor of Neuropharmacology at the Institute of Medical Sciences of the University of Aberdeen, he is also Director of Pharmacology for GW Pharmaceuticals.
AUTHOR CONTRIBUTIONS
Z.‐X.X., R.G.P., M.I.D., A.H.L., and E.L.G. were responsible for the study concept and design. Z.‐X.X., P.M., X.‐F.W., G.‐H.B., M.I.D., and A.H.L. designed the animal experiments and carried out the experiments. Z.‐X.X., P.M., M.I.D., A.H.L., and E.L.G. analysed and interpreted the data. Z.‐X.X., M.I.D., and E.L.G. drafted the manuscript. E.L.G. extensively revised the manuscript into its final form, with valuable additional input from R.G.P. and M.I.D. All authors critically reviewed the manuscript content and approved the final version for publication.
DECLARATION OF TRANSPARENCY AND SCIENTIFIC RIGOUR
This Declaration acknowledges that this paper adheres to the principles for transparent reporting and scientific rigour of preclinical research as stated in the BJP guidelines for https://bpspubs.onlinelibrary.wiley.com/doi/abs/10.1111/bph.14207, and https://bpspubs.onlinelibrary.wiley.com/doi/full/10.1111/bph.14206, and as recommended by funding agencies, publishers and other organisations engaged with supporting research.
ACKNOWLEDGEMENTS
This research was supported by funds from the Intramural Research Program of the U.S. National Institute on Drug Abuse (Z.‐X.X., G.‐H.B., X.‐F.W., and E.L.G.) and by the National Institute on Drug Abuse research grant R01‐DA005274 (M.I.D.).
Xi Z‐X, Muldoon P, Wang X‐F, et al. Δ8‐Tetrahydrocannabivarin has potent anti‐nicotine effects in several rodent models of nicotine dependence. Br J Pharmacol. 2019;176:4773–4784. 10.1111/bph.14844
This article has been contributed to by US Government employees and their work is in the public domain in the USA.
REFERENCES
- Adamczyk, P. , Miszkiel, J. , McCreary, A. C. , Filip, M. , Papp, M. , & Przegaliński, E. (2012). The effects of cannabinoid CB1, CB2 and vanilloid TRPV1 receptor antagonists on cocaine addictive behavior in rats. Brain Research, 1444, 45–54. 10.1016/j.brainres.2012.01.030 [DOI] [PubMed] [Google Scholar]
- Alexander, S. P. H. , Christopoulos, A. , Davenport, A. P. , Kelly, E. , Marrion, N. V. , Peters, J. A. , … CGTP Collaborators (2017). The concise guide to pharmacology 2017/18: G protein‐coupled receptors. British Journal of Pharmacology, 174, S17–S129. 10.1111/bph.13878 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bátkai, S. , Mukhopadhyay, P. , Horváth, B. , Rajesh, M. , Gao, R. Y. , Mahadevan, A. , … Pacher, P. (2012). Δ8‐Tetrahydrocannabivarin prevents hepatic ischaemia/reperfusion injury by decreasing oxidative stress and inflammatory responses through cannabinoid CB2 receptors. British Journal of Pharmacology, 165, 2450–2461. 10.1111/j.1476-5381.2011.01410.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bolognini, D. , Costa, B. , Maione, S. , Comelli, F. , Marini, P. , Di Marzo, V. , … Pertwee, R. G. (2010). The plant cannabinoid Δ9‐tetrahydrocannabivarin can decrease signs of inflammation and inflammatory pain in mice. British Journal of Pharmacology, 160, 677–687. 10.1111/j.1476-5381.2010.00756.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cahill, K. , & Ussher, M. H. (2011). Cannabinoid type 1 receptor antagonists for smoking cessation. Cochrane Database of Systematic Reviews, 2011(3), CD005353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Campos, A. C. , Fogaça, M. V. , Aguiar, D. C. , & Guimarães, F. S. (2013). Animal models of anxiety disorders and stress. Revista Brasileira de Psiquiatria, 35(Suppl 2), S101–S111. 10.1590/1516-4446-2013-1139 [DOI] [PubMed] [Google Scholar]
- Canseco‐Alba, A. , Schanz, N. , Sanabria, B. , Zhao, J. , Lin, Z. , Liu, Q.‐R. , & Onaivi, E. S. (2018). Behavioral effects of psychostimulants in mutant mice with cell‐type specific deletion of CB2 cannabinoid receptors in dopamine neurons. Behavioural Brain Research, 360, 286–297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Christensen, R. , Kristensen, P. K. , Bartels, E. M. , Bliddal, H. , & Astrup, A. (2007). Efficacy and safety of the weight‐loss drug rimonabant: A meta‐analysis of randomised trials. Lancet, 370, 1706–1713. 10.1016/S0140-6736(07)61721-8 [DOI] [PubMed] [Google Scholar]
- Cohen, C. , Kodas, E. , & Griebel, G. (2005). CB1 receptor antagonists for the treatment of nicotine addiction. Pharmacology, Biochemistry, and Behavior, 81, 387–395. 10.1016/j.pbb.2005.01.024 [DOI] [PubMed] [Google Scholar]
- Damaj, M. I. , Kao, W. , & Martin, B. R. (2003). Characterization of spontaneous and precipitated nicotine withdrawal in the mouse. The Journal of Pharmacology and Experimental Therapeutics, 307, 526–534. 10.1124/jpet.103.054908 [DOI] [PubMed] [Google Scholar]
- De Vries, T. J. , Shaham, Y. , Homberg, J. R. , Crombag, H. , Schuurman, K. , Dieben, J. , … Schoffelmeer, A. N. (2001). A cannabinoid mechanism in relapse to cocaine seeking. Nature Medicine, 7, 1151–1154. 10.1038/nm1001-1151 [DOI] [PubMed] [Google Scholar]
- Delis, F. , Polissidis, A. , Poulia, N. , Justinova, Z. , Nomikos, G. G. , Goldberg, S. R. , & Antoniou, K. (2017). Attenuation of cocaine‐induced conditioned place preference and motor activity via cannabinoid CB2 receptor agonism and CB1 receptor antagonism in rats. The International Journal of Neuropsychopharmacology, 20, 269–278. 10.1093/ijnp/pyw102 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elrashidi, M. Y. , & Ebbert, J. O. (2014). Emerging drugs for the treatment of tobacco dependence: 2014 update. Expert Opinion on Emerging Drugs, 19, 243–260. 10.1517/14728214.2014.899580 [DOI] [PubMed] [Google Scholar]
- ElSohly, M. A. , Radwan, M. M. , Gul, W. , Chandra, S. , & Galal, A. (2017). Phytochemistry of Cannabis sativa L. In Kinghorn A., Falk H., Gibbons S., & Kobayashi J. (Eds.), Phytocannabinoids: Unraveling the complex chemistry and pharmacology of Cannabis sativa. Progress in the chemistry of organic natural products (Vol. 103) (pp. 1–36). Cham, Switzerland: Springer International Publishing Switzerland. [DOI] [PubMed] [Google Scholar]
- Englund, A. , Atakan, Z. , Kralj, A. , Tunstall, N. , Murray, R. , & Morrison, P. (2016). The effect of five day dosing with THCV on THC‐induced cognitive, psychological and physiological effects in healthy male human volunteers: A placebo‐controlled, double‐blind, crossover pilot trial. Journal of Psychopharmacology, 30, 140–151. 10.1177/0269881115615104 [DOI] [PubMed] [Google Scholar]
- Gamaleddin, I. , Zvonok, A. , Makriyannis, A. , Goldberg, S. R. , & Le Foll, B. (2012). Effects of a selective cannabinoid CB2 agonist and antagonist on intravenous nicotine self‐administration and reinstatement of nicotine seeking. PLoS ONE, 7(1), e29900 10.1371/journal.pone.0029900 Epub 2012 Jan 26 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gamaleddin, I. H. , Trigo, J. M. , Gueye, A. B. , Zvonok, A. , Makriyannis, A. , Goldberg, S. R. , & Le Foll, B. (2015). Role of the endogenous cannabinoid system in nicotine addiction: Novel insights. Frontiers in Psychiatry, 6(Article 41), 1–12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gardner, E. L. (2002). Addictive potential of cannabinoids: The underlying neurobiology. Chemistry and Physics of Lipids, 121, 267–290. 10.1016/S0009-3084(02)00162-7 [DOI] [PubMed] [Google Scholar]
- Gardner, E. L. (2005). Endocannabinoid signaling system and brain reward: Emphasis on dopamine. Pharmacology, Biochemistry, and Behavior, 81, 263–284. 10.1016/j.pbb.2005.01.032 [DOI] [PubMed] [Google Scholar]
- Gardner, E. L. (2014). Cannabinoids and addiction In Pertwee R. G. (Ed.), Handbook of cannabis (pp. 173–188). Oxford, UK: Oxford Univ Press; 10.1093/acprof:oso/9780199662685.003.0009 [DOI] [Google Scholar]
- Gardner, E. L. , Gamaleddin, I. , Manzanares Robles, J. , & Rodrígues de Fonseca, F. (2013). The endocannabinoid system: Useful targets for anti‐addiction treatments? Substance Abuse, 34, 324–325. [Google Scholar]
- Gill, E. W. , Paton, W. D. M. , & Pertwee, R. G. (1970). Preliminary experiments on the chemistry and pharmacology of cannabis. Nature, 228, 134–136. 10.1038/228134a0 [DOI] [PubMed] [Google Scholar]
- Government of Canada/Gouvernement du Canada (2018). Cannabis in Canada: Get the facts. Available at https://www.canada.ca/en/services/health/campaigns/cannabis/canadians.html ().
- Grotenhermen F., & Russo E. (Eds.) (2002). Cannabis and cannabinoids: Pharmacology, toxicology, and therapeutic potential. Binghamton, NY: Haworth Press. [Google Scholar]
- Gueye, A. B. , Pryslawsky, Y. , Trigo, J. M. , Poulia, N. , Delis, F. , Antoniou, K. , … Le Foll, B. (2016). The CB1 neutral antagonist AM4113 retains the therapeutic efficacy of the inverse agonist rimonabant for nicotine dependence and weight loss with better psychiatric tolerability. The International Journal of Neuropsychopharmacology, 19(12), 1–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harding, S. D. , Sharman, J. L. , Faccenda, E. , Southan, C. , Pawson, A. J. , Ireland, S. , … NC‐IUPHAR (2018). The IUPHAR/BPS guide to pharmacology in 2018: Updates and expansion to encompass the new guide to immunopharmacology. Nucleic Acids Research, 46, D1091–D1106. 10.1093/nar/gkx1121 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harmey, D. , Griffin, P. R. , & Kenny, P. J. (2012). Development of novel pharmacotherapeutics for tobacco dependence: Progress and future directions. Nicotine & Tobacco Research, 14, 1300–1318. 10.1093/ntr/nts201 [DOI] [PMC free article] [PubMed] [Google Scholar]
- He, X.‐H. , Jordan, C. J. , Vemuri, K. , Bi, G.‐H. , Zhan, J. , Gardner, E. L. , … Xi, Z. X. (2019). Cannabinoid CB1 receptor neutral antagonist AM4113 inhibits heroin self‐administration without depressive side effects in rats. Acta Pharmacologica Sinica, 40, 365–373. 10.1038/s41401-018-0059-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hillig, K. W. , & Mahlberg, P. G. (2004). A chemotaxonomic analysis of variation in Cannabis (Cannabaceae). American Journal of Botany, 91, 966–975. 10.3732/ajb.91.6.966 [DOI] [PubMed] [Google Scholar]
- Howlett, A. C. , Barth, F. , Bonner, T. I. , Cabral, G. , Casellas, P. , Devane, W. A. , … Pertwee, R. G. (2002). International Union of Pharmacology. XXVII. Classification of cannabinoid receptors. Pharmacological Reviews, 54, 161–202. [DOI] [PubMed] [Google Scholar]
- Hughes, J. R. , Peters, E. N. , & Naud, S. (2008). Relapse to smoking after 1 year of abstinence: A meta‐analysis. Addictive Behaviors, 33, 1516–1520. 10.1016/j.addbeh.2008.05.012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ignatowska‐Jankowska, B. M. , Muldoon, P. P. , Lichtman, A. H. , & Damaj, M. I. (2013). The cannabinoid CB2 receptor is necessary for nicotine‐conditioned place preference, but not other behavioral effects of nicotine in mice. Psychopharmacology, 229, 591–601. 10.1007/s00213-013-3117-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jackson, K. J. , Martin, B. R. , Changeux, J.‐P. , & Damaj, M. I. (2008). Differential role of nicotinic acetylcholine receptor subunits in physical and affective nicotine withdrawal signs. The Journal of Pharmacology and Experimental Therapeutics, 325, 302–312. 10.1124/jpet.107.132977 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jadoon, K. A. , Ratcliffe, S. H. , Barrett, D. A. , Thomas, E. L. , Stott, C. , Bell, J. D. , … Tan, G. D. (2016). Efficacy and safety of cannabidiol and tetrahydrocannabivarin on glycemic and lipid parameters in patients with type 2 diabetes: A randomized, double‐blind, placebo‐controlled, parallel group pilot study. Diabetes Care, 39, 1777–1786. 10.2337/dc16-0650 [DOI] [PubMed] [Google Scholar]
- Jordan, C. J. , & Xi, Z.‐X. (2019). Progress in brain cannabinoid CB2 receptor research: From genes to behavior. Neuroscience and Biobehavioral Reviews, 98, 208–220. 10.1016/j.neubiorev.2018.12.026 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kilkenny, C. , Browne, W. , Cuthill, I. C. , Emerson, M. , & Altman, D. G. (2010). Animal research: Reporting in vivo experiments: The ARRIVE guidelines. British Journal of Pharmacology, 160, 1577–1579. 10.1111/j.1476-5381.2010.00872.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kirk, R. E. (1982). Experimental design (2nd ed.). Belmont, CA: Brooks/Cole. [Google Scholar]
- Kota, D. , Martin, B. R. , Robinson, S. E. , & Damaj, M. I. (2007). Nicotine dependence and reward differ between adolescent and adult male mice. The Journal of Pharmacology and Experimental Therapeutics, 322, 399–407. 10.1124/jpet.107.121616 [DOI] [PubMed] [Google Scholar]
- Kwilasz, A. J. , Harris, L. S. , & Vann, R. E. (2009). Removal of continuous nicotine infusion produces somatic but not behavioral signs of withdrawal in mice. Pharmacology, Biochemistry, and Behavior, 94, 114–118. 10.1016/j.pbb.2009.07.015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu, Q.‐R. , Pan, C.‐H. , Hishimoto, A. , Li, C.‐Y. , Xi, Z.‐X. , Llorente‐Berzal, A. , … Uhl, G. R. (2009). Species differences in cannabinoid receptor 2 (CNR2 gene): Identification of novel human and rodent CB2 isoforms, differential tissue expression and regulation by cannabinoid receptor ligands. Genes, Brain, and Behavior, 8, 519–530. 10.1111/j.1601-183X.2009.00498.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lumeng, L. , Hawkins, T. D. , & Li, T.‐K. (1977). New strains of rats with alcohol preference and non‐preference In Thurman R. G., Williamson J. R., Drott H., & Chance B. (Eds.), Alcohol and aldehyde metabolizing systems (Vol. 3) (pp. 537–544). New York, NY: Academic Press; 10.1016/B978-0-12-691403-0.50056-2 [DOI] [Google Scholar]
- Lupica, C. R. , Riegel, A. C. , & Hoffman, A. F. (2004). Marijuana and cannabinoid regulation of brain reward circuits. British Journal of Pharmacology, 143, 227–234. 10.1038/sj.bjp.0705931 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maldonado, R. , Valverde, O. , & Berrendero, F. (2006). Involvement of the endocannabinoid system in drug addiction. Trends in Neurosciences, 29, 225–232. 10.1016/j.tins.2006.01.008 [DOI] [PubMed] [Google Scholar]
- Manzanares, J. , Cabañero, D. , Puente, N. , García‐Gutiérrez, M. S. , Grandes, P. , & Maldonado, R. (2018). Role of the endocannabinoid system in drug addiction. Biochemical Pharmacology, 157, 108–121. [DOI] [PubMed] [Google Scholar]
- Matsuda, L. A. (1997). Molecular aspects of cannabinoid receptors. Critical Reviews in Neurobiology, 11, 143–166. 10.1615/CritRevNeurobiol.v11.i2-3.30 [DOI] [PubMed] [Google Scholar]
- Matsuda, L. A. , Lolait, S. J. , Brownstein, M. J. , Young, A. C. , & Bonner, T. I. (1990). Structure of a cannabinoid receptor and functional expression of the cloned cDNA. Nature, 346, 561–564. 10.1038/346561a0 [DOI] [PubMed] [Google Scholar]
- McPartland, J. M. , Duncan, M. , Di Marzo, V. , & Pertwee, R. G. (2015). Are cannabidiol and Δ9‐tetrahydrocannabivarin negative modulators of the endocannabinoid system? A systematic review. British Journal of Pharmacology, 172, 737–753. 10.1111/bph.12944 [DOI] [PMC free article] [PubMed] [Google Scholar]
- McPartland, J. M. , Glass, M. , & Pertwee, R. G. (2007). Meta‐analysis of cannabinoid ligand binding affinity and receptor distribution: Interspecies differences. British Journal of Pharmacology, 152, 583–593. 10.1038/sj.bjp.0707399 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Munro, S. , Thomas, K. L. , & Abu‐Shaar, M. (1993). Molecular characterization of a peripheral receptor for cannabinoids. Nature, 365, 61–65. 10.1038/365061a0 [DOI] [PubMed] [Google Scholar]
- National Conference of State Legislatures (2018). State medical marijuana laws. Available at http://www.ncsl.org/research/health/state-medical-marijuana-laws.aspx ().
- National Research Council (2011). Guide for the care and use of laboratory animals (8th ed.). Washington, DC: National Academies Press. [Google Scholar]
- Navarrete, F. , García‐Gutiérrez, M. S. , & Manzanares, J. (2018). Pharmacological regulation of cannabinoid CB2 receptor modulates the reinforcing and motivational actions of ethanol. Biochemical Pharmacology, 157, 227–234. 10.1016/j.bcp.2018.07.041 [DOI] [PubMed] [Google Scholar]
- Navarrete, F. , Rodríguez‐Arias, M. , Martín‐García, E. , Navarro, D. , García‐Gutiérrez, M. S. , Aguilar, M. A. , … Manzanares, J. (2013). Role of CB2 cannabinoid receptors in the rewarding, reinforcing, and physical effects of nicotine. Neuropsychopharmacology, 38, 2515–2524. 10.1038/npp.2013.157 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pertwee, R. G. (2008). The diverse CB1 and CB2 receptor pharmacology of three plant cannabinoids: Δ9‐Tetrahydrocannabinol, cannabidiol and Δ9‐tetrahydrocannabivarin. British Journal of Pharmacology, 153, 199–215. 10.1038/sj.bjp.0707442 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pertwee, R. G. , Thomas, A. , Stevenson, L. A. , Ross, R. A. , Varvel, S. A. , Lichtman, A. H. , … Razdan, R. K. (2007). The psychoactive plant cannabinoid, Δ9‐tetrahydrocannabinol, is antagonized by Δ8‐ and Δ9‐tetrahydrocannabivarin in mice in vivo. British Journal of Pharmacology, 150, 586–594. 10.1038/sj.bjp.0707124 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rose, J. E. (2009). New findings on nicotine addiction and treatment. Nebraska Symposium on Motivation, 55, 131–141. [DOI] [PubMed] [Google Scholar]
- Sam, A. H. , Salem, V. , & Ghatei, M. A. (2011). Rimonabant: From RIO to ban. Journal of Obesity, 2011, 432607 10.1155/2011/432607 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schmidt, B. L. , Tambeli, C. H. , Gear, R. W. , & Levine, J. D. (2001). Nicotine withdrawal hyperalgesia and opioid‐mediated analgesia depend on nicotine receptors in nucleus accumbens. Neuroscience, 106, 129–136. 10.1016/S0306-4522(01)00264-0 [DOI] [PubMed] [Google Scholar]
- Stolerman, I. P. , & Jarvis, M. J. (1995). The scientific case that nicotine is addictive. Psychopharmacology, 117, 2–10; discussion 14‐20. 10.1007/BF02245088 [DOI] [PubMed] [Google Scholar]
- Tanda, G. , & Goldberg, S. R. (2003). Cannabinoids: Reward, dependence, and underlying neurochemical mechanisms—Review of recent preclinical data. Psychopharmacology, 169, 115–134. 10.1007/s00213-003-1485-z [DOI] [PubMed] [Google Scholar]
- Turner, C. E. , Hadley, K. , & Fetterman, P. S. (1973). Constituents of Cannabis sativa L. VI: Propyl homologs in samples of known geographical origin. Journal of Pharmaceutical Sciences, 62, 1739–1741. 10.1002/jps.2600621045 [DOI] [PubMed] [Google Scholar]
- U.S. Department of Health and Human Services (2010). How tobacco smoke causes disease: The biology and behavioral basis for smoking‐attributable disease: A report of the surgeon general. Atlanta, GA: U.S. Department of Health and Human Services, Centers for Disease Control and Prevention, National Center for Chronic Disease Prevention and Health Promotion, Office on Smoking and Health. [PubMed] [Google Scholar]
- Van Gaal, L. F. , Rissanen, A. M. , Scheen, A. J. , Ziegler, O. , Rössner, S. , & for the RIO‐Europe Study Group (2005). Effects of the cannabinoid‐1 receptor blocker rimonabant on weight reduction and cardiovascular risk factors in overweight patients: 1‐year experience from the RIO‐Europe study. Lancet, 365, 1389–1397. [DOI] [PubMed] [Google Scholar]
- Van Sickle, M. D. , Duncan, M. , Kingsley, P. J. , Mouihate, A. , Urbani, P. , Mackie, K. , … Sharkey, K. A. (2005). Identification and functional characterization of brainstem cannabinoid CB2 receptors. Science, 310, 329–332. 10.1126/science.1115740 [DOI] [PubMed] [Google Scholar]
- Venniro, M. , Caprioli, D. , & Shaham, Y. (2016). Animal models of drug relapse and craving: From drug priming‐induced reinstatement to incubation of craving after voluntary abstinence. Progress in Brain Research, 224, 25–52. 10.1016/bs.pbr.2015.08.004 [DOI] [PubMed] [Google Scholar]
- Wang, X.‐F. , Bi, G.‐H. , He, Y. , Yang, H.‐J. , Gao, J.‐T. , Okunola‐Bakare, O. M. , … Newman, A. H. (2015). R‐modafinil attenuates nicotine‐taking and nicotine‐seeking behavior in alcohol‐preferring rats. Neuropsychopharmacology, 40, 1762–1771. 10.1038/npp.2015.24 [DOI] [PMC free article] [PubMed] [Google Scholar]
- World Health Organization (2013). WHO report on the global tobacco epidemic, 2013. Geneva, Switzerland: WHO Press, World Health Organization. [Google Scholar]
- Xi, Z.‐X. , Gilbert, J. G. , Peng, X.‐Q. , Pak, A. C. , Li, X. , & Gardner, E. L. (2006). Cannabinoid CB1 receptor antagonist AM 251 inhibits cocaine‐primed relapse in rats: Role of glutamate in the nucleus accumbens. The Journal of Neuroscience, 26, 8531–8536. 10.1523/JNEUROSCI.0726-06.2006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xi, Z.‐X. , Peng, X.‐Q. , Li, X. , Song, R. , Zhang, H.‐Y. , Liu, Q.‐R. , … Gardner, E. L. (2011). Brain cannabinoid CB2 receptors modulate cocaine's actions in mice. Nature Neuroscience, 14, 1160–1166. 10.1038/nn.2874 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xi, Z.‐X. , Spiller, K. , Pak, A. C. , Gilbert, J. , Dillon, C. , Li, X. , … Gardner, E. L. (2008). Cannabinoid CB1 receptor antagonists attenuate cocaine's rewarding effects: Experiments with self‐administration and brain‐stimulation reward in rats. Neuropsychopharmacology, 33, 1735–1745. 10.1038/sj.npp.1301552 [DOI] [PubMed] [Google Scholar]
- Zhang, H.‐Y. , Bi, G.‐H. , Li, X. , Li, J. , Qu, H. , Zhang, S.‐J. , … Liu, Q. R. (2015). Species differences in cannabinoid receptor 2 and receptor responses to cocaine self‐administration in mice and rats. Neuropsychopharmacology, 40, 1037–1051. 10.1038/npp.2014.297 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang, H.‐Y. , Gao, M. , Liu, Q.‐R. , Bi, G.‐H. , Li, X. , Yang, H.‐J. , … Xi, Z. X. (2014). Cannabinoid CB2 receptors modulate midbrain dopamine neuronal activity and dopamine‐related behavior in mice. Proceedings of the National Academy of Sciences of the United States of America, 111, E5007–E5015. 10.1073/pnas.1413210111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang, H.‐Y. , Gao, M. , Shen, H. , Bi, G.‐H. , Yang, H.‐J. , Liu, Q.‐R. , … Xi, Z. X. (2017). Expression of functional cannabinoid CB2 receptor in VTA dopamine neurons in rats. Addiction Biology, 22, 752–765. 10.1111/adb.12367 [DOI] [PMC free article] [PubMed] [Google Scholar]