

Abstract Abstract
Lake Kastoria is one of the potentially “ancient” Balkan lakes that has a great environmental importance and ecological value, attracts high touristic interest and is under various anthropogenic pressures. It belongs to a Natura 2000 Special Protection Area and a Site of Community Interest. The city of Kastoria is located at the western part of the lake and just next to it, towards the centre of the lake, is a peninsula, a habitat island. In the framework of research concerning the flora of lake islands of Greece, one of the main objectives of the present study is to fill a gap concerning plant species richness of the habitat island within the protected Lake Kastoria, which is surrounded by the lake except for its north-western part where the border of the city of Kastoria is located. Floristic analysis of the habitat island of Lake Kastoria is in large measure accounted, concerning chorology with emphasis on Balkan endemics (8.7%), life forms, by hemicryptophytes (36.1%), therophytes (33.2%), phanerophytes (16.4%) and geophytes (10.9%) and, for habitats, by taxa preferring agricultural and ruderal ones (53.3%). Another objective is to compare its floristic composition to the one of the island within the protected urban Lake Pamvotis - one of the very few lake islands in Greece - focusing on the influence of urbanisation. The α- and β- diversity are measured in order to reveal floristic differences. Beta diversity partitioning in turnover and nestedness showed that the β-diversity is mostly expressed as compositional turnover. The role of the society in combination with long-term programmes for the study of plant species richness, functional diversity and patterns of species assemblages over time are necessary for the effective management and protection of protected areas, including lake insular areas.
Keywords: protected area, Natura 2000, plant diversity, α-diversity, β-diversity, spatial turnover, monitoring
Introduction
The majority of ancient or putatively ancient European lakes is restricted to the Balkan area and is thought to be restricted within a cluster of lakes, about 300 km around Lakes Ohrid and Prespa (Touka et al. 2018), including lakes such as the lakes Skutari (Montenegro, Albania), Mikri Prespa (Greece, Albania), Vegoritis, Kastoria or Orestias, Pamvotis and Trichonis (Greece). Lake Kastoria or Orestias is one of the potentially “ancient” lakes of the Balkans, meaning long-lived modern or palaeo-lakes (Frogley et al. 2001, Frogley et al. 2002, Frogley and Preece 2004). Lake Pamvotis is an ancient lake of Pleistocenic biota, characterised as a Quaternary refugium (Frogley et al. 2001, Tzedakis 2002).
Lake Kastoria, belongs to the Natura 2000 Special Protection Area GR1320003 and Site of Community Importance GR1320001 (Fig. 1) and it falls under the responsibility of the Protected Areas of Western Macedonia Management Body. Lake Pamvotis belongs to the Natura 2000 Special Protection Area and Site of Community Importance GR2130005 and it falls under the responsibility of the Lake Pamvotis Management Body (see Sarika et al. 2019). Kastoria and Pamvotis lakes, both urban lakes, are rather heavily modified (Latinopoulos et al. 2018). Considering the fact that Lake Kastoria is an urban lake with a population of 17,000 inhabitants living on the shoreline and 35,000 in the entire catchment, it is easy to understand that this lake is affected by human activity. This is also the case for Lake Pamvotis, near which Ioannina city is located with a population of more than 120,000 inhabitants (Kati et al. 2006, Sarika et al. 2019).
Figure 1.
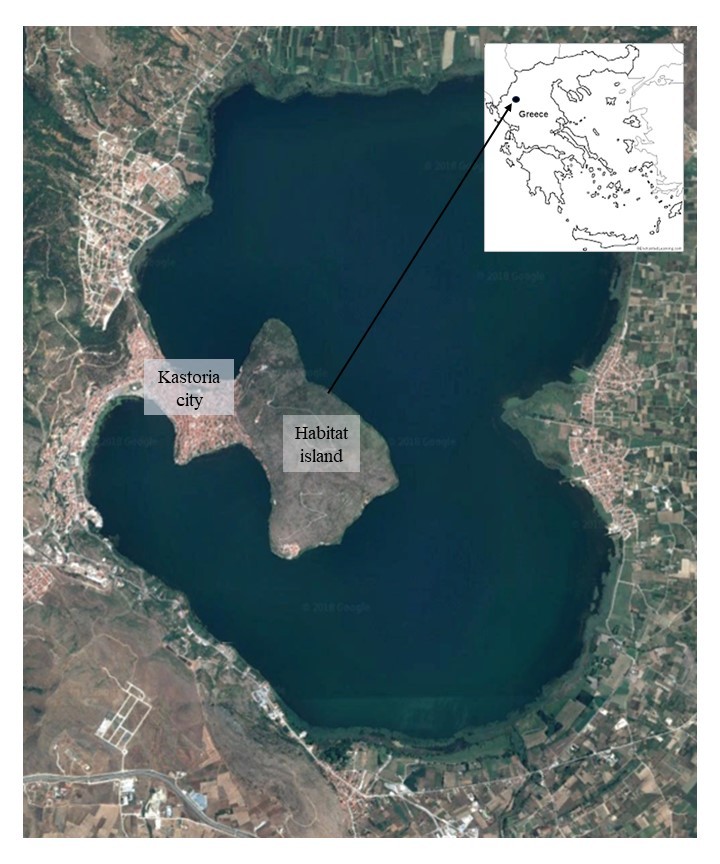
Map of the habitat island within Lake Kastoria.
Both lakes have attracted research interests as sedimentary archives on long term environmental and climate history and as a hotspot for European biodiversity (Touka et al. 2018). Many studies have also been published, concerning the hydrology and hydromorphology, the assessment of the typology and the trophic status, the plankton, the hydrophyte vegetation of North-western Greece's lakes, focusing on the lakes of Kastoria and Pamvotis (amongst others Koussouris et al. 1987, Latinopoulos et al. 2016, Latinopoulos et al. 2018, Moustaka-Gouni et al. 2006, Papastergiadou et al. 2009, Stefanidis and Papastergiadou 2007, Stefanidis and Papastergiadou 2010, Katsiapi et al. 2016, Kagalou and Psilovikos 2014, Moustaka-Gouni et al. 2007).
Lakes are “negative islands”, that is, they are more or less isolated freshwater areas surrounded by a hostile land matrix, behaving as islands in many biogeographical and ecological respects (Whittaker and Fernandez-Palacios 2007). Since lakes behave like that, any type of island (true or habitat) within them is a lake island in a negative island. Habitat islands are essentially all forms of insular systems that do not qualify as being “real islands”. They are discrete patches of distinct terrestrial habitat surrounded by strongly contrasting habitats (Whittaker and Fernandez-Palacios 2007). Within both studied lakes, there are islands. A true island can be found within Lake Pamvotis (Sarika et al. 2019) and a habitat island, a peninsula, within Lake Kastoria. This habitat island, neighbouring the city of Kastoria, is still floristically unexplored and is surrounded by the lake apart from its north-western part where the border of the city is located (Fig. 1). Human interference is obvious both in the protected lakes as well as in the two lake islands. Thus, it should be noted that the need for monitoring species diversity in protected areas is urgent and the assessment of species diversity is crucial (Chelli et al. 2019). In order to understand the patterns of species richness, they can be separated into different components: α-diversity that represents the species richness of a local habitat and β-diversity that is the component that represents the difference in species composition between sample units or between habitats and can be driven by nestedness that reflects a non-random process of disaggregation of assemblages and turnover that is a process of substitution of species by environmental selection or historical and spatial restriction (Whittaker 1965, Coelho et al. 2018, Baselga 2010).
In the framework of this study concerning plant species richness of lake islands of North-Western Greece, the main objectives are a) to fill the gap in the floristic information available concerning the study of the floristic composition of the habitat island within the protected Lake Kastoria and b) to depict the differences in species richness and composition with those of the island within the protected Pamvotis lake (Sarika et al. 2019), measuring α-diversity and estimating β-diversity through the contribution of spatial turnover and nestedness and focusing on the influence of urbanisation, since both are within urban lakes.
Material and methods
Survey area
Lake Kastoria has a surface area of 27.9 km2 at an altitude of 630 m in the Kastoria Prefecture, North-Western Greece. It has an average depth of 4.4 m and a maximum depth of 9.1 m. The city of Kastoria (> 47,160 inhabitants) is located on the western part of the lake and discharges from there have resulted in increased eutrophication (Moustaka-Gouni et al. 2007). Lake Kastoria is a very fragile shallow aquatic ecosystem, long challenged by the various rural (logging, agricultural wastes, stockbreeding etc.), craft (tanneries, fur/leather production) and urban (e.g. sewer discharges, rubble depositions and extensive littering) activities of the area. The area is a significant touristic destination, especially the caves that have been formed (Dragon Cave), (Kagalou and Psilovikos 2014). The eastern border of the city of Kastoria forms the north-western border of the habitat island studied (KaHI), the peninsula, that is surrounded by the lake at all its other parts (Fig. 1). Human intervention is evident with urban development dominating in Lake Kastoria (Kagalou and Psilovikos 2014, Kati et al. 2006).
Lake Pamvotis has a surface area of 22 km2 at an average altitude of 470 m. It is a shallow lake (mean depth of 4.3 m) and the city of Ioannina (>120,000 inhabitants) is located at its south-western part. Although ecologically important, the ecosystem of the lake is seriously affected by organic (Kagalou et al. 2003) and heavy metal (Papagiannis et al. 2004) pollution, riparian habitat urbanisation and hydrological regime disruption. For the purpose of this research, the temporal turnover of plant species richness of the island within Lake Pamvotis (PaI) that attracts >170,000 visitors every year, has been studied (Sarika et al. 2019).
Data Collection and Floristic analysis
The present study concerns plant species richness and composition of the habitat island within Lake Kastoria (KaHI) and comparison with those of the true island within the protected Pamvotis lake (PaI). It is based on our team's (a) fieldwork at KaHI, from April 2018 to October 2019 and (b) published research concerning plant species diversity of PaI, the island within Lake Pamvotis (Sarika et al. 2019, see sampling period Β), from May 2018 to April 2019. Sample collections on KaHI and PaI during 2018-2019 were carried out by Α. Papanikolaou and Μ. Panitsa and plant specimens are deposited in UPA Herbarium. The sampling scheme used included fieldwork (plant specimen collections and observations) all over KaHI during all seasons of the year and this was also the case for PaI. For the determination of the plant material, Tutin et al. (1964), Tutin et al. (1972), Tutin et al. (1976), Tutin et al. (1980), Tutin et al. (1981), Strid and Tan (1997) and Strid and Tan (2002) were used. The families, genera, species and subspecies are listed, within the major taxonomic groups in alphabetical order. Nomenclature, life form, chorology, status categories and habitat preferences of plant taxa follow Dimopoulos et al. (2013). The six main life forms have been used for the floristic analysis as were given by Dimopoulos et al. (2013) and are summarised as follows: phanerophytes (P), chamaephytes (Ch), hemicryptophytes (H), geophytes (G), therophytes (T) and aquatics (Aq). Chorological categories follow the system of Dimopoulos et al. (2013) and for the analysis, they have been grouped to 5 main chorological elements: Widespread, Mediterranean, Balkan, Greek endemics and Alien.
In the framework of the authors' research, a list of all the information recorded has been created, including the plant taxa registered on KaHI and those of PaI (Sarika et al. 2019), their biological and chorological types and their habitat preferences according to the data provided by the “Vascular Plants of Greece" (Dimopoulos et al. 2013; see also "Flora of Greece web" 2017+, http://portal.cybertaxonomy.org/flora-greece-intro).
The α-diversity is one of the components of species richness, measured as the number of species occurring in a sample unit. In this study, the sample unit is the habitat island (surface area of 27.9 km2) and its α-diversity is compared with that of the island within Pamvotis lake (surface area of 22 km2). To allow floristic comparisons between KaHI and PaI, a table presenting floristic similarities was prepared. The chi-square test, which is a well-suited statistical tool for these purposes, was used to compare biological and chorological spectra between KaHI and PaI. In order to test β-diversity between them, we checked its two components: spatial species turnover using Sørensen dissimilarity index (bsor) and nestedness of assemblages using nestedness-resultant dissimilarity (bnes) index (Baselga 2010). The Sørensen dissimilarity index (bsor) is one of the most used measures due to its dependence on the proportion of species shared between two communities, that incorporates both true spatial turnover and differences in richness (Koleff et al. 2003). The Sørensen dissimilarity index (bsor) is formulated as: 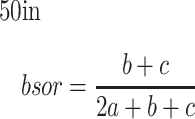 where a is the number of species common to both sites, b is the number of species that occur in the first site, but not in the second and c is the number of species that occur in the second site, but not in the first. The nestedness-resultant dissimilarity (bnes) index is a measure of the dissimilarity of communities due to the effect of nestedness patterns (Baselga 2010) and is formulated as:
where a is the number of species common to both sites, b is the number of species that occur in the first site, but not in the second and c is the number of species that occur in the second site, but not in the first. The nestedness-resultant dissimilarity (bnes) index is a measure of the dissimilarity of communities due to the effect of nestedness patterns (Baselga 2010) and is formulated as: 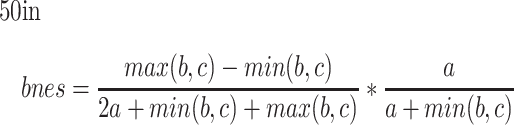
Results
There is no previous documented information concerning plant species richness of the habitat island within the protected lake of Kastoria studied. Field investigations on the island revealed a total number of 274 plant taxa recorded, out of which 2 are Pteridophyta, 4 Gymnospermae and 268 Angiospermae, belonging to 196 genera and 74 families (Table 1).
Table 1.
List of plant taxa recorded on the habitat island within Lake Kastoria. Abbreviations: Bk = Balkan chorological element, X = alien, xenophyte, non-native plant taxa including cultigens, permanently established in at least one floristic region of Greece, r = range-restricted plant taxon characterised by a restricted distribution, by populations occurring along a linear distance not exceeding 500 km and R = ruderal taxon
| Plant Families | Plant taxa | Bk | Status | R |
| Pteridophytes | ||||
| Aspleniaceae | Asplenium ceterach L. | |||
| Dennstaedticeae | Pteridium aquilinum (L.) Kuhn | |||
| Gymnosperms | ||||
| Cupressaceae | Cupressus sempervirens L. | |||
| Juniperus oxycedrus subsp. deltoides (R.P. Adams) N.G.Passal. | ||||
| Ephedraceae | Ephedra foeminea Forskål | |||
| Pinaceae | Pinus brutia Ten. (cultivated) | |||
| Angiosperms | ||||
| Acanthaceae | Acanthus hungaricus (Borbás) Baen. | * | ||
| Aceraceae | Acer platanoides L. | |||
| Acer pseudoplatanus L. | ||||
| Alliaceae | Allium amethystinum Tausch | 1 | ||
| Allium bornmuelleri Hayek | * | r | ||
| Allium cyrilli Ten. | 1 | |||
| Allium guttatum Steven | ||||
| Allium rhodopeum Velen. | 1 | |||
| Allium sphaerocephalon L. | 1 | |||
| Amaranthaceae | Amaranthus quitensis Kunth | X | 1 | |
| Amaryllidaceae | Sternbergia lutea (L.) Sprend | |||
| Anacardiaceae | Pistacia terebinthus L. | |||
| Apiaceae | Anthriscus sylvestris subsp. nemorosus (M.Bieb.) Koso-Pol. | 1 | ||
| Daucus carota L. | 1 | |||
| Eryngium campestre L. | 1 | |||
| Foeniculum vulgare Mill. | 1 | |||
| Malabaila involucrata Boiss. & Spruner | * | 1 | ||
| Oenanthe aquatica (L.) Poir. | ||||
| Orlaya daucoides (L.) Greuter | 1 | |||
| Pastinaca hirsuta Pančić | * | 1 | ||
| Scandix australis subsp. grandiflora (L.) Thell. | 1 | |||
| Tordylium apulum L. | 1 | |||
| Apocynaceae | Nerium oleander L. subsp. oleander (obs.) | |||
| Araceae | Arum italicum Mill. subsp. italicum | |||
| Arum maculatum L. | ||||
| Araliaceae | Hedera helix L. | |||
| Aristolochiaceae | Aristolochia clematitis L. | 1 | ||
| Asclepiadaceae | Cionura erecta (L.) Griseb. | 1 | ||
| Asparagaceae | Asparagus acutifolius L. | |||
| Asphodelaceae | Asphodeline lutea (L.) Rchb. | |||
| Asteraceae | Achillea clypeolata Sm. | |||
| Achillea millefolium L. | ||||
| Anthemis arvensis L. | 1 | |||
| Artemisia vulgaris L. | 1 | |||
| Calendula arvensis (Vaill.) L. | 1 | |||
| Carduus nutans subsp. leiophyllus (Petrović) Stoj. & Stef. | 1 | |||
| Carduus pycnocephalus L. | 1 | |||
| Centaurea graeca Griseb. | * | r | ||
| Cichorium intybus L. | 1 | |||
| Crepis neglecta L. | ||||
| Echinops sphaerocephalus L. | 1 | |||
| Erigeron canadensis L. | X | 1 | ||
| Lactuca muralis (L.) Gaertn. | ||||
| Lactuca viminea (L.) J. Presl & C. Presl subsp. viminea | ||||
| Leontodon hispidus L. subsp. hispidus | ||||
| Matricaria recutita L. | 1 | |||
| Podospermum canum C.A.Mey. | ||||
| Senecio vernalis Waldst. & Kit. | 1 | |||
| Senecio vulgaris L. | 1 | |||
| Silybum marianum (L.) Gaertn. | 1 | |||
| Sonchus oleraceus L. | 1 | |||
| Taraxacum sect. Fontana Soest | ||||
| Taraxacum sect. Ruderalia Kirschner & al. | 1 | |||
| Tragopogon porrifolius subsp. eriospermus (Ten.) Greuter | 1 | |||
| Xanthium strumarium L. | 1 | |||
| Boraginaceae | Anchusa officinalis subsp. intacta (Griseb.) Selvi & Bigazzi | 1 | ||
| Cynoglossum officinale L. | 1 | |||
| Echium italicum subsp. biebersteinii (Lacaita) Greuter & Burdet | 1 | |||
| Heliotropium europaeum L. | 1 | |||
| Lithospermum officinale L. | ||||
| Myosotis ramosissima Rochel | 1 | |||
| Myosotis sicula Guss. | ||||
| Brassicaceae | Alliaria petiolata (M.Bieb.) Cavara & Grande | 1 | ||
| Alyssum chalcidicum Janka | * | r | 1 | |
| Arabidopsis thaliana (L.) Heynh. | 1 | |||
| Arabis hirsuta (L.) Scop. | ||||
| Aubrieta deltoidea (L.) DC. | ||||
| Aurinia saxatilis (L.) Desv. | ||||
| Calepina irregularis (Asso) Thell. | 1 | |||
| Capsella bursa-pastoris (L.) Medik. | 1 | |||
| Cardamine hirsuta L. | 1 | |||
| Clypeola jonthlaspi L. | 1 | |||
| Diplotaxis tenuifolia (L.) DC. | 1 | |||
| Draba verna L. | ||||
| Erysimum crassistylum C. Presl | ||||
| Hesperis laciniata All. subsp. laciniata | ||||
| Hirschfeldia incana (L.) Lagr.-Foss. | 1 | |||
| Lepidium draba L. | 1 | |||
| Lunaria annua subsp. pachyrhiza (Borbás) Maire & Petitm. | 1 | |||
| Microthlaspi perfoliatum (L.) F.K.Mey. | 1 | |||
| Rapistrum rugosum (L.) All. | 1 | |||
| Sisymbrium officinale (L.) Scop. | 1 | |||
| Teesdalia coronopifolia (J.P.Bergeret) Thell. | ||||
| Caesalpiniaceae | Cercis siliquastrum L. | |||
| Campanulaceae | Asyneuma limonifolium (L.) Janch. | |||
| Campanula sparsa Friv. | * | |||
| Campanula versicolor Andrews | ||||
| Caprifoliaceae | Sambucus nigra L. | 1 | ||
| Caryophyllaceae | Cerastium glomeratum Thuill. | 1 | ||
| Holosteum umbellatum L. subsp. umbellatum | 1 | |||
| Petrorhagia dubia (Raf.) G.López & Romo | 1 | |||
| Petrorhagia prolifera (L.) P.W.Ball & Heywood | ||||
| Petrorhagia saxifraga (L.) Link | ||||
| Silene graeca Boiss. & Spruner | * | r? | ||
| Silene latifolia Poir. | 1 | |||
| Silene ungeri Fenzl | * | r | ||
| Silene vulgaris (Moench) Garcke | 1 | |||
| Stellaria media (L.) Vill. | 1 | |||
| Celastraceae | Euonymus verrucosus Scop. | |||
| Chenopodiaceae | Chenopodium album L. | 1 | ||
| Convolvulaceae | Calystegia sepium (L.) R.Br. | |||
| Convolvulus althaeoides L. | 1 | |||
| Convolvulus arvensis L. | 1 | |||
| Convolvulus cantabrica L. | ||||
| Cuscuta campestris Yunck. | X | 1 | ||
| Crassulaceae | Sedum acre L. | |||
| Sedum album L. | ||||
| Sedum annuum L. | ||||
| Umbilicus horizontalis (Guss.) DC. | ||||
| Umbilicus rupestris (Salisb.) Dandy | ||||
| Dipsacaceae | Pterocephalus plumosus (L.) Coult. | |||
| Scabiosa tenuis Boiss. | * | r? | ||
| Euphorbiaceae | Euphorbia cyparissias L. | 1 | ||
| Euphorbia helioscopia L. | 1 | |||
| Euphorbia myrsinites L. | ||||
| Euphorbia platyphyllos L. subsp. platyphyllos | 1 | |||
| Mercurialis annua L. | 1 | |||
| Mercurialis ovata Sternb. & Hoppe | ||||
| Fabaceae | Anthyllis vulneraria subsp. bulgarica (Sagorski) Cullen | * | ||
| Colutea arborescens L. subsp. arborescens | ||||
| Galega officinalis L. | 1 | |||
| Hippocrepis emerus subsp. emeroides (Boiss. & Spruner) Lassen | ||||
| Lathyrus setifolius L. | 1 | |||
| Lotus pedunculatus Cav. | ||||
| Medicago arabica (L.) Huds. | 1 | |||
| Medicago minima (L.) Bartal. | 1 | |||
| Medicago polymorpha L. | 1 | |||
| Medicago sativa L. subsp. sativa | 1 | |||
| Medicago sativa subsp. falcata (L.) Arcang. | 1 | |||
| Melilotus albus Medik. | 1 | |||
| Melilotus officinalis (L.) Lam. | 1 | |||
| Oxytropis pilosa (L.) DC. | ||||
| Robinia pseudoacacia L. | X | 1 | ||
| Spartium junceum L. | 1 | |||
| Trifolium campestre Schreb. | ||||
| Trifolium dubium Sibth. | ||||
| Trifolium pratense L. | ||||
| Trifolium repens L. | 1 | |||
| Vicia sativa L. | 1 | |||
| Vicia villosa subsp. microphylla (d'Urv.) P.W.Ball | 1 | |||
| Vicia villosa subsp. varia (Host) Corb. | 1 | |||
| Fagaceae | Quercus frainetto Ten. | |||
| Fumariaceae | Fumaria rostellata Knaf | 1 | ||
| Geraniaceae | Erodium ciconium (L.) L'Hér. | 1 | ||
| Geranium lucidum L. | 1 | |||
| Geranium molle L. | 1 | |||
| Geranium purpureum Vill. | 1 | |||
| Geranium rotundifolium L. | 1 | |||
| Hippocastanaceae | Aesculus hippocastanum L. | * | ||
| Hyacinthaceae | Muscari comosum (L.) Mill. | 1 | ||
| Muscari neglectum Ten. | ||||
| Ornithogalum sibthorpii Greuter | ||||
| Prospero autumnale (L.) Speta | ||||
| Hypericaceae | Hypericum perforatum L. | 1 | ||
| Hypericum rumeliacum Boiss. | * | |||
| Iridaceae | Crocus cancellatus Herb. subsp. maziaricus (Herb.) B.Mathew | |||
| Iris attica Boiss. & Heldr. | ||||
| Juglandaceae | Juglans regia L. | |||
| Lamiaceae | Ballota nigra subsp. sericea (Vandas) Patzak | * | r | 1 |
| Lamium amplexicaule L. | 1 | |||
| Lamium garganicum L. subsp. garganicum | ||||
| Marrubium peregrinum L. | 1 | |||
| Mentha aquatica L. | ||||
| Salvia candidissima Vahl | ||||
| Sideritis montana L. subsp. montana | 1 | |||
| Stachys annua (L.) L. | 1 | |||
| Stachys plumosa Griseb. | * | |||
| Teucrium capitatum L. | ||||
| Thymus longicaulis C. Presl | ||||
| Thymus teucrioides Boiss. & Spruner | * | r | ||
| Lauraceae | Laurus nobilis L. | |||
| Liliaceae | Lilium candidum L. | |||
| Lythraceae | Lythrum salicaria L. | |||
| Malvaceae | Alcea biennis subsp. cretica (Weinm.) Valdés | * | 1 | |
| Malva sylvestris L. | 1 | |||
| Moraceae | Broussonetia papyrifera (L.) Vent. | X | 1 | |
| Ficus carica L. | ||||
| Morus alba L. | X | 1 | ||
| Oleaceae | Jasminum fruticans L. | |||
| Phillyrea latifolia L. | ||||
| Orchidaceae | Ophrys helenae Renz | * | r | |
| Ophrys sphegodes Mill. | ||||
| Orobanchaceae | Orobanche sp | |||
| Papaveraceae | Chelidonium majus L. | 1 | ||
| Papaver dubium L. | 1 | |||
| Papaver rhoeas L. | 1 | |||
| Plantaginaceae | Plantago lanceolata L. | 1 | ||
| Plantago major L. subsp. major | 1 | |||
| Platanaceae | Platanus orientalis L. | |||
| Poaceae | Aegilops neglecta Bertol. | 1 | ||
| Avena barbata Link | 1 | |||
| Avena sterilis L. | 1 | |||
| Bothriochloa ischaemum (L.) Keng | ||||
| Bromus diandrus Roth | 1 | |||
| Bromus sterilis L. | 1 | |||
| Cynodon dactylon (L.) Pers. | 1 | |||
| Dactylis glomerata L. | 1 | |||
| Dasypyrum villosum (L.) P.Candargy | 1 | |||
| Hordeum marinum Huds. | ||||
| Hordeum murinum subsp. leporinum (Link) Arcang. | 1 | |||
| Lolium perenne L. | 1 | |||
| Lolium rigidum Gaudin | 1 | |||
| Melica ciliata L. | ||||
| Melica transsilvanica subsp. klokovii Tzvelev | ||||
| Milium vernale M. Bieb. | 1 | |||
| Phleum subulatum (Savi) Asch. & Graebn. | ||||
| Piptatherum holciforme (M. Bieb.) Roem. & Schult. | ||||
| Poa annua L. | 1 | |||
| Poa bulbosa L. | ||||
| Poa nemoralis L. | ||||
| Poa trivialis L. | ||||
| Setaria pumila (Poir.) Roem. & Schult. | 1 | |||
| Setaria viridis (L.) P. Beauv. subsp. viridis | 1 | |||
| Stipa thessala Hausskn. | * | r | ||
| Polygonaceae | Rumex acetosella subsp. acetoselloides (Balansa) Nijs | 1 | ||
| Rumex conglomeratus Murray | ||||
| Rumex crispus L. | 1 | |||
| Portulacaceae | Portulaca oleracea L. s.l. | 1 | ||
| Primulaceae | Cyclamen hederifolium Aiton | |||
| Lysimachia vulgaris L. | ||||
| Ranunculaceae | Anemone pavonina Lam. | |||
| Clematis flammula L. | ||||
| Delphinium balcanicum Pawł. | * | r | 1 | |
| Delphinium peregrinum L. | 1 | |||
| Nigella damascena L. | 1 | |||
| Ranunculus psilostachys Griseb. | * | |||
| Rhamnaceae | Paliurus spina-christi Mill. | |||
| Rosaceae | Crataegus monogyna Jacq. | |||
| Prunus avium (L.) L. | ||||
| Prunus domestica subsp. insititia (L.) Bonnier & Layens | 1 | |||
| Prunus dulcis (Mill.) D.A. Webb | X | 1 | ||
| Prunus spinosa subsp. dasyphylla (Schur) Domin | ||||
| Prunus webbii (Spach) Vierh. | ||||
| Pyracantha coccinea M. Roem. | ||||
| Rosa canina L. | ||||
| Rubus canescens DC. | ||||
| Rubus sanctus Schreb. | 1 | |||
| Sanguisorba minor Scop. | ||||
| Rubiaceae | Galium aparine L. | 1 | ||
| Galium setaceum subsp. decaisnei (Boiss.) Ehrend. | ||||
| Galium spurium L. | 1 | |||
| Galium verum L. subsp. verum | ||||
| Ruscaceae | Ruscus aculeatus L. | |||
| Rutaceae | Dictamnus albus L. | |||
| Salicaceae | Populus alba L. | |||
| Populus nigra L. | ||||
| Saxifragaceae | Saxifraga tridactylites L. | 1 | ||
| Scrophulariaceae | Linaria dalmatica (L.) Mill. | 1 | ||
| Linaria genistifolia (L.) Mill. subsp. genistifolia | 1 | |||
| Linaria peloponnesiaca Boiss. & Heldr. | * | 1 | ||
| Scrophularia heterophylla Willd. | ||||
| Verbascum graecum Boiss. | * | r? | 1 | |
| Verbascum undulatum Lam. | * | 1 | ||
| Simaroubaceae | Ailanthus altissima (Mill.) Swingle | X | 1 | |
| Smilacaceae | Smilax excelsa L. | |||
| Tiliaceae | Tilia rubra DC. | |||
| Ulmaceae | Celtis australis L. | |||
| Ulmus minor Mill. | ||||
| Urticaceae | Parietaria cretica L. | |||
| Parietaria officinalis L. | ||||
| Urtica dioica L. | 1 | |||
| Valerianaceae | Valeriana italica Lam. | |||
| Valerianella echinata (L.) DC. | 1 | |||
| Valerianella locusta (L.) Laterr. | 1 | |||
| Veronicaceae | Veronica hederifolia L. | 1 | ||
| Veronica polita Fr. | 1 | |||
| Veronica triloba (Opiz) Opiz | 1 | |||
| Vervenaceae | Verbena officinalis L. | 1 |
Asteraceae (25 taxa), Poaceae (25 taxa), Fabaceae (23 taxa), Brassicaceae (21 taxa), Lamiaceae (12 taxa), Rosaceae (11), Caryophyllaceae (10) and Apiaceae (10 taxa) are the richest in taxa families in the total of plant taxa recorded. Taxa belonging to these families represent 50% of the total flora of the habitat island (Fig. 2). The richest in taxa genera are: Allium (6 taxa), Prunus and Medicago (5 taxa), Euphorbia, Galium, Geranium, Silene, Trifolium and Poa (4 taxa each), (Fig. 3).
Figure 2.

Richest in taxa families of the habitat island within Lake Kastoria.
Figure 3.

Richest in taxa genera of the habitat island within Lake Kastoria.
Regarding the chorological origin of the taxa, widespread taxa dominate (55.1%), followed by Mediterranean elements (33.3%), Balkan taxa (8.7%) and alien taxa, xenophytes (2.9%). Fig. 4 shows proportions of different chorological elements recorded. The presence of Balkan elements is significant, since 24 taxa have been registered (Table 1). Eight naturalised xenophytes have been registered, belonging to 7 families, namely Ailanthus altissima, Amaranthus quitensis, Broussonetia papyrifera, Cuscuta campestris, Erigeron canadensis, Morus alba, Prunus dulcis and Robinia pseudoacacia.
Figure 4.

Proportion of chorological categories of the plant taxa diversity of the habitat island within Lake Kastoria.
According to the IUCN Red List (IUCN 2019), Allium bornmuelleri is a range-restricted Balkan endemic, characterised as Data Deficient as also are Allium cyrilli and Prunus webbii, while Allium amethystinum, A. sphaerocephalon, Prunus avium, P. spinosa and Dictamnus albus are characterised as of Least Concern. Lilium candidum, has been assessed as Near Threatened in the national red list (Phitos et al. 2009).
On the life form spectrum, Hemicryptophytes are the most dominant (36.1%), followed by Therophytes (33.2%), Phanerophytes (16.4%) and Geophytes (10.9%) (Fig. 5). The evaluation of habitat preferences of plant taxa reveals that plants (exclusive or non-exclusive) of agricultural and ruderal habitats (52%) are the most common, followed by plants of grasslands (17.8%), of woodland and scrub (21.1%), exclusively of cliffs and rocks (2.5%) and of freshwater habitats (4%) (Fig. 6).
Figure 5.

Life form spectrum of the plant diversity of the habitat island within Lake Kastoria.
Figure 6.

Proportion of different habitat categories for taxa recorded in the habitat island within lake Kastoria: Freshwater habitats (A), Cliffs and rocks (C), temperate and submediterranean grasslands (G), High mountain vegetation (H), agricultural and ruderal habitats (R) and Woodlands and scrub (W). x = generalist taxa.
The α-diversity for KaHI is 9.9 per km2 and for PaI 10.5 per km2. Comparing their floristic composition, 105 taxa were common (25.2%), 169 (40.5%) were registered only on KaHI and 143 (34.3%) only on PaI (Sarika et al. 2019) during their second sampling period from May 2018 to April 2019 (Fig. 7). Using the chi-square test, the observed data of biological and chorological spectra of KaHI and PaI were compared to a null hypothesis that distributes the data according to the expectation that they are due to chance. The observed data do not fit the model, since for biological types: df = 4, x2 = 0.457 and p = 0.977 and for chorological types: df = 3, x2 = 0.002 and p = 0.999, thus proving the null hypothesis correct and the relation of the data not statistically significant.
Figure 7.

Contribution of different species richness variables of the habitat island within Lake Kastoria (KaHI) and of the island of Lake Pamvotis (PaI), on the beta diversity components: spatial turnover (bsor) and nestedness (bnes). Abbreviations: R = ruderal taxa, Th = Therophytes, Hm = Hemicryptophytes, Ph = Phanerophytes, Ge = Geophytes, Bk = Balkan taxa.
Concerning beta diversity, the values of its components, bsor and bnes are presented in Fig. 7. Values of bnes are lower than the values of bsor for all species richness variables. The values of bsor are higher than 0.5 on different functional categories as life forms (therophytes, hemicryptophytes, geophytes), chorological elements (Balkan taxa) and habitat preferences (ruderal taxa, ruderal therophytes). Of the common taxa, 61.9% were ruderal, 3.8% were alien (Robinia pseudoacacia, Erigeron canadensis, Ailanthus altissima, Cuscuta campestris) and 5.7% were Balkan and range restricted (Centaurea graeca, Alyssum chalcidicum, Silene ungeri, Ballotanigra subsp. sericea, Ophrys helenae and Scabiosa tenuis).
Discussion
Lake Kastoria, one of the potentially “ancient” Balkan lakes, is a protected lake of a great environmental importance and ecological value. Lake Kastoria’s habitat island and Lake Pamvotis’ island (both in NW Greece) are two of the very few lake islands occurring in Greece – together with the small islands of Agios Achilleios and Vidronisi within Prespa Lake (N Greece) - all belonging to protected lake areas.
There has been a gap so far concerning plant species richness of the habitat island studied, which is included in the protected area of Lake Kastoria, (GR1320001 and GR1320003). During field investigations conducted on the island, 274 plant taxa were registered in total, belonging to 74 families and 196 genera and presenting a rather high α-diversity. Of the taxa recorded, in total, 38.7% belong to the families Asteraceae, Poaceae, Lamiaceae, Fabaceae and Brassicaceae, as is also the case with the island within Lake Pamvotis (Sarika et al. 2019).
The combination of the high percentage of Mediterranean taxa and the high percentage of therophytes, reflects the Mediterranean character of the flora of the studied habitat island as well as of the Lake Pamvotis island. The proportion of alien taxa is about 2.9%, less than the one recorded for the Greek flora as a whole (5% according to Arianoutsou et al. 2010; 3.8% according to Dimopoulos et al. 2013), but in a restricted area.
The α-diversity, the diversity at a local scale of the two lake islands, was rather similar. Panitsa and Tzanoudakis (2010) stated that the number of species per surface unit (alpha-diversity) is an important parameter that highlights the role of small islands in the conservation of the diversity. The Echinades islet group (Ionian area, Western Greece), hosts a high number of taxa per surface unit (Iliadou et al. 2014), as is also the case in the East Aegean islet groups of Arki and Lipsi (Panitsa and Tzanoudakis 2001). Concerning island biogeography, species richness on different islands is likely to be particularly affected by species richness on the mainland and the degree of inter-island dispersal (Bunnefeld and Phillimore 2012).
The two lake islands studied present remarkable similarities as well as significant differences in their floristic composition. Both islands are located within urban lakes in NW Greece, have a small surface area, are of strong continental character and they also share the same rather strong human strains. Cody (2006) showed that plant species turnover is a highly variable phenomenon, depending on the functional traits of each species, the local geomorphology and the ecological conditions of the studied areas. Urban development fragments, isolates and degrades natural habitats; it also simplifies and homogenises species composition, causing a more linear decline of richness with increasing urbanisation in regions where anthropogenic impact outside cities is more limited (Luck and Smallbone 2010, Norton et al. 2016, Oliveira Hagen et al. 2017). The restructuring of biotic communities in urban areas is non-random and strongly associated with the loss of species, with limited tolerance to urban development and increased abundances of more tolerant species, with urban environments filtering species according to their ecological and life-history traits (amongst others Fischer et al. 2015, Oliveira Hagen et al. 2017). This filtering process occurs across all three stages of the biotic urbanisation process, i.e. arrival, adaptation and spread (Evans et al. 2010). Urbanisation changes seed dispersal functions which promote invasive plant species over closely-related native species (Caughlin et al. 2016).
Ruderal plant taxa are typically occurring and prevailing in disturbed areas, in agricultural and ruderal habitats and especially in sites with pronounced direct or indirect human activity, rural and urban sites, roadsides, excessively grazed and trampled sites, as well as in naturally nutrient-rich and frequently disturbed pioneer habitats (Dimopoulos et al. 2013). On the contrary, species that do not make much use anthropogenic resources, but are largely reliant on the natural habitat lying within a matrix of urban development, face the challenge of maintaining movement through an increasingly fractured landscape to access remnant patches of suitable habitat and have to adapt to declining habitat quality and encroachment of weeds, as well as to the presence of introduced feral and domestic predators within these habitat patches (Bryant et al. 2017). The high percentage of common ruderal taxa between the two lake islands reveals their contribution to their plant species composition, mainly due to the influence of the nearby urban ecosystems and due to human activities in the studied area, in general. Human activities are amongst the factors enhancing turnover (Panitsa et al. 2008) and ruderal flora and vegetation generally represent one of the most dynamic floristic-vegetation complexes (Pyšek et al. 2017).
Beta diversity partitioning in turnover and nestedness showed that the β-diversity of the studied two, habitat and true, islands was mostly expressed as compositional turnover since the nestedness component was lower than the turnover one. This result was mostly based on changes in turnover rates (such as changes in floristic composition), rather than a change due to nestedness. Changes in community composition and spatial structure (spatial beta diversity) need a lot of work to be quantified (Dornelas et al. 2014, McGill et al. 2015) and focusing on the composition of plant species richness of the two lake islands showed that human interference is continuous and the values of the spatial turnover component of β-diversity are significant. A high percentage of common taxa between the two islands are ruderal therophytes and hemicryptophytes but they still conserve floristic dissimilarity concerning Balkan endemic taxa. That means that functional diversity indices allow the quantification and comparison of functional diversity amongst communities much more than simple measurements of biodiversity (Pla et al. 2012).
Both protected lakes and the islands within them suffer because of various anthropogenic pressures affecting their plant species and functional diversity (Latinopoulos et al. 2016, Latinopoulos et al. 2018, Papastergiadou et al. 2009, Stefanidis and Papastergiadou 2007, Stefanidis and Papastergiadou 2010). Since they are urban lakes, urbanisation significantly influences their ecosystems’ functioning and the services they provide to humans (Alberti 2016, Alberti et al. 2003). Consequently, the role of society on effective management and conservation of these urban affected islands will be crucial. Additionally, long-term programmes for the study of plant species richness, functional diversity and patterns of species assemblages over time in protected areas, as in the studied area, is more than necessary (Chelli et al. 2019) for their effective management and protection.
Conflicts of interest
There are no conflicts of interest.
Conflicts of interest
There are no conflicts of interest.
References
- Alberti MARINA, Marzluff JOHN M., Shulenberger ERIC, Bradley GORDON, Ryan CLARE, Zumbrunnen CRAIG. Integrating humans into ecology: Opportunities and challenges for studying urban ecosystems. BioScience. 2003;53(12):1169–1179. doi: 10.1641/0006-3568(2003)053[1169:ihieoa]2.0.co;2. [DOI] [Google Scholar]
- Alberti Marina. The effects of urban patterns on ecosystem function. International Regional Science Review. 2016;28(2):168–192. doi: 10.1177/0160017605275160. [DOI] [Google Scholar]
- Arianoutsou Margarita, Bazos Ioannis, Delipetrou Pinelopi, Kokkoris Yannis. The alien flora of Greece: taxonomy, life traits and habitat preferences. Biological Invasions. 2010;12(10):3525–3549. doi: 10.1007/s10530-010-9749-0. [DOI] [Google Scholar]
- Baselga Andrés. Partitioning the turnover and nestedness components of beta diversity. Global Ecology and Biogeography. 2010;19(1):134–143. doi: 10.1111/j.1466-8238.2009.00490.x. [DOI] [Google Scholar]
- Bryant Gillian L., Kobryn Halina T., Hardy Giles E. StJ., Fleming Patricia A. Habitat islands in a sea of urbanisation. Urban Forestry & Urban Greening. 2017;28:131–137. doi: 10.1016/j.ufug.2017.10.016. [DOI] [Google Scholar]
- Bunnefeld Nils, Phillimore Albert B. Island, archipelago and taxon effects: mixed models as a means of dealing with the imperfect design of nature's experiments. Ecography. 2012;35(1):15–22. doi: 10.1111/j.1600-0587.2011.07078.x. [DOI] [Google Scholar]
- Caughlin Trevor, Wheeler Jessica H., Jankowski Jill, Lichstein Jeremy W. Urbanized landscapes favored by fig-eating birds increase invasive but not native juvenile strangler fig abundance. Figshare. 2016 doi: 10.6084/M9.FIGSHARE.C.3304794. [DOI] [PubMed]
- Chelli Stefano, Simonetti Enrico, Campetella Giandiego, Chiarucci Alessandro, Cervellini Marco, Tardella Federico Maria, Tomasella Michela, Canullo Roberto. Plant diversity changes in a nature reserve: a probabilistic sampling method for quantitative assessments. Nature Conservation. 2019;34:145–161. doi: 10.3897/natureconservation.34.30043. [DOI] [Google Scholar]
- Cody M. Plants on islands: Diversity and dynamics on a continental archipelago. University of California Press, Berkeley; 2006. [DOI] [Google Scholar]
- Coelho Marcel Serra, Carneiro Marco Antônio Alves, Branco Cristina Alves, Borges Rafael Augusto Xavier, Fernandes Geraldo Wilson. Species turnover drives β-diversity patterns across multiple spatial scales of plant-galling interactions in mountaintop grasslands. PLOS One. 2018;13(5):e0195565.. doi: 10.1371/journal.pone.0195565. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dimopoulos P, Raus T, Bergmeier E, Constantinidis T, Iatrou G, Kokkini S, Strid A, Tzanoudakis D. Vascular plants of Greece: An annotated checklist. Englera. 2013;31:1–372. [Google Scholar]
- Dornelas M., Gotelli N. J., McGill B., Shimadzu H., Moyes F., Sievers C., Magurran A. E. Assemblage time series reveal biodiversity change but not systematic loss. Science. 2014;344(6181):296–299. doi: 10.1126/science.1248484. [DOI] [PubMed] [Google Scholar]
- Evans Karl L., Hatchwell Ben J., Parnell Mark, Gaston Kevin J. A conceptual framework for the colonisation of urban areas: the blackbird Turdus merula as a case study. Biological Reviews. 2010;85:643–667. doi: 10.1111/j.1469-185x.2010.00121.x. [DOI] [PubMed] [Google Scholar]
- Fischer Jason D., Schneider Sarah C., Ahlers Adam A., Miller James R. Categorizing wildlife responses to urbanization and conservation implications of terminology. Conservation Biology. 2015;29(4):1246–1248. doi: 10.1111/cobi.12451. [DOI] [PubMed] [Google Scholar]
- Frogley M. R., Griffiths H. I., Heaton T. H. E. Historical biogeography and Late Quaternary environmental change of Lake Pamvotis, Ioannina (north-western Greece): evidence from ostracods. Journal of Biogeography. 2001;28(6):745–756. doi: 10.1046/j.1365-2699.2001.00582.x. [DOI] [Google Scholar]
- Frogley M. R., Griffiths H. I., Martens K. Modern and fossil ostracods from ancient lakes. Holmes J. A., Chivas A. R., editors. 167-184The Ostracoda: Applications in Quaternary research. 2002 doi: 10.1029/131GM09. [DOI]
- Frogley M. R., Preece R. C. A faunistic review of the modern and fossil molluscan fauna from Lake Pamvotis, Ioannina, an Ancient Lake in NWw Greece: Implications for endemism in the Balkans. In: Griffiths H. I., Kryštufek B., Reed J. M., editors. Balkan Biodiversity. Springer; Dordrecht: 2004. 243-260. [DOI] [Google Scholar]
- Iliadou Eleni, Panitsa Maria, Raus Thomas, Dimopoulos Panayotis. Flora and factors affecting species diversity in protected “Natura 2000” sites of the Ionian area: the Echinades islet group (Greece) Willdenowia. 2014;44(1):121–136. doi: 10.3372/wi.44.44113. [DOI] [Google Scholar]
- IUCN The IUCN Red List of threatened species. Version 2019-2. http://www.iucnredlist.org
- Kagalou I., Papastergiadou E., Tsimarakis G., Petridis D. Evaluation of the trophic state of Lake Pamvotis Greece, a shallow urban lake. Hydrobiologia. 2003:745–752. doi: 10.1023/b:hydr.0000008603.69847.9e. [DOI]
- Kagalou Ifigenia, Psilovikos Aris. Assessment of the typology and the trophic status of two Mediterranean lake ecosystems in Northwestern Greece. Water Resources. 2014;41(3):335–343. doi: 10.1134/s0097807814030105. [DOI] [Google Scholar]
- Kati Vassiliki, Mani Paraskevi, von Helversen Otto, Willemse Fer, Elsner Norbert, Dimopoulos Panayotis. Human land use threatens endemic wetland species: The case of Chorthippus lacustris (La Greca and Messina 1975) (Orthoptera: Acrididae) in Epirus, Greece. Journal of Insect Conservation. 2006;10(1):65–74. doi: 10.1007/s10841-005-2642-y. [DOI] [Google Scholar]
- Katsiapi Matina, Moustaka-Gouni Maria, Sommer Ulrich. Assessing ecological water quality of freshwaters: PhyCoI—a new phytoplankton community Index. Ecological Informatics. 2016;31:22–29. doi: 10.1016/j.ecoinf.2015.11.004. [DOI] [Google Scholar]
- Koleff Patricia, Gaston Kevin J., Lennon Jack J. Measuring beta diversity for presence-absence data. Journal of Animal Ecology. 2003;72(3):367–382. doi: 10.1046/j.1365-2656.2003.00710.x. [DOI] [Google Scholar]
- Koussouris Th., Diapoulis A., Balopoulos E. Limnological situations in two shallow Greek Lakes (Kastoria and Mikri Prespa lakes) GeoJournal. 1987;14(3) doi: 10.1007/bf00208214. [DOI] [Google Scholar]
- Latinopoulos Dionissis, Ntislidou Chrysoula, Kagalou Ifigeneia. Multipurpose plans for the sustainability of the Greek Lakes: Emphasis on multiple stressors. Environmental Processes. 2016;3(3):589–602. doi: 10.1007/s40710-016-0152-4. [DOI] [Google Scholar]
- Latinopoulos Dionissis, Ntislidou Chrysoula, Kagalou Ifigenia. A multi-approach lake habitat survey method for impact assessment in two heavily modified lakes: a case of two Northern Greek lakes. Environmental Monitoring and Assessment. 2018;190:658. doi: 10.1007/s10661-018-7045-0. [DOI] [PubMed] [Google Scholar]
- Luck G. W., Smallbone L. T. Species diversity and urbanisation: patterns, drivers and implications. In: Gaston K. J., editor. Urban Ecology. Cambridge University Press; 2010. 88-119. [DOI] [Google Scholar]
- McGill Brian J., Dornelas Maria, Gotelli Nicholas J., Magurran Anne E. Fifteen forms of biodiversity trend in the Anthropocene. Trends in Ecology & Evolution. 2015;30(2):104–113. doi: 10.1016/j.tree.2014.11.006. [DOI] [PubMed] [Google Scholar]
- Moustaka-Gouni Maria, Vardaka Elisabeth, Michaloudi Evangelia, Kormas Konstantinos Ar., Tryfon Eleni, Mihalatou Helen, Gkelis Spyros, Lanaras Tom. Plankton food web structure in a eutrophic polymictic lake with a history of toxic cyanobacterial blooms. Limnology and Oceanography. 2006;51:715–727. doi: 10.4319/lo.2006.51.1_part_2.0715. [DOI] [Google Scholar]
- Moustaka-Gouni Maria, Vardaka Elisabeth, Tryfon Eleni. Phytoplankton species succession in a shallow Mediterranean lake (L. Kastoria, Greece): steady-state dominance of Limnothrix redekei, Microcystis aeruginosa and Cylindrospermopsis raciborskii. Hydrobiologia. 2007;575(1):129–140. doi: 10.1007/s10750-006-0360-4. [DOI] [Google Scholar]
- Norton Briony A., Evans Karl L., Warren Philip H. Urban biodiversity and landscape ecology: Patterns, processes and planning. Current Landscape Ecology Reports. 2016;1(4):178–192. doi: 10.1007/s40823-016-0018-5. [DOI] [Google Scholar]
- Oliveira Hagen Emily, Hagen Oskar, Ibáñez-Álamo Juan D., Petchey Owen L., Evans Karl L. Impacts of urban areas and their characteristics on avian functional diversity. Frontiers in Ecology and Evolution. 2017;5 doi: 10.3389/fevo.2017.00084. [DOI] [Google Scholar]
- Panitsa Maria, Tzanoudakis Dimitris. A floristic investigation of the islet groups Arki and Lipsi (East Aegean Area, Greece) Folia Geobotanica. 2001;36(3):265–279. doi: 10.1007/bf02803180. [DOI] [Google Scholar]
- Panitsa Maria, Tzanoudakis Dimitrios, Sfenthourakis Spyros. Turnover of plants on small islets of the eastern Aegean Sea within two decades. Journal of Biogeography. 2008;35(6):1049–1061. doi: 10.1111/j.1365-2699.2007.01846.x. [DOI] [Google Scholar]
- Panitsa Maria, Tzanoudakis Dimitris. Floristic diversity on small islands and islets: Leros islets’ group (East Aegean area, Greece) Phytologia Balcanica. 2010;16(2):271–284. [Google Scholar]
- Papagiannis I., Kagalou I., Leonardos J., Petridis D., Kalfakakou V. Copper and zinc in four freshwater fish species from Lake Pamvotis (Greece) Environment International. 2004;30(3):357–362. doi: 10.1016/j.envint.2003.08.002. [DOI] [PubMed] [Google Scholar]
- Papastergiadou E., Kagalou I., Stefanidis K., Retalis A., Leonardos I. Effects of anthropogenic influences on the trophic state, land uses and aquatic vegetation in a shallow Mediterranean lake: Implications for estoration. Water Resources Management. 2009;24(3):415–435. doi: 10.1007/s11269-009-9453-y. [DOI] [Google Scholar]
- Phitos D, Constantinidis T, Kamari G. The Red Data Book of rare and threatened plants of Greece. Vols. I, II. Hellenic Βotanical Society; Patras: 2009. Greek. [Google Scholar]
- Pla Laura, Casanoves Fernando, Di Rienzo Julio. Quantifying functional biodiversity. Springer Briefs in Environmental Science; 2012. [DOI] [Google Scholar]
- Pyšek Petr, Pergl Jan, Essl Franz, Lenzner Bernd, Dawson Wayne, Kreft Holger, Weigelt Patrick, Winter Marten, Kartesz John, Nishino Misako, Antonova Liubov A., Barcelona Julie F., Cabesaz Francisco J., Cárdenas Dairon, Cárdenas-Toro Juliana, Castaño Nicolás, Chacón Eduardo, Chatelain Cyrille, Dullinger Stefan, Ebel Aleksandr L., Figueiredo Estrela, Fuentes Nicol, Genovesi Piero, Groom Quentin J., Henderson Lesley, Inderjit, Kupriyanov Andrey, Masciadri Silvana, Maurel Noëlie, Meerman Jan, Morozova Olga, Moser Dietmar, Nickrent Daniel, Nowak Pauline M., Pagad Shyama, Patzelt Annette, Pelser Pieter B., Seebens Hanno, Shu Wen-sheng, Thomas Jacob, Velayos Mauricio, Weber Ewald, Wieringa Jan J., Baptiste María P., Kleunen Mark van. Naturalized alien flora of the world. Preslia. 2017;89(3):203–274. doi: 10.23855/preslia.2017.203. [DOI] [Google Scholar]
- Sarika Maria, Papanikolaou Alexandros, Yannitsaros Artemios, Chitos Theodoros, Panitsa Maria. Temporal turnover of the flora of lake islands: The island of Lake Pamvotis (Epirus, Greece) Biodiversity Data Journal. 2019;7:e37023. doi: 10.3897/bdj.7.e37023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stefanidis K, Papastergiadou E. Aquatic vegetation and related abiotic environment in a shallow urban lake of Greece. Belgian Journal of Botany. 2007;140(1):25–38. [Google Scholar]
- Stefanidis K., Papastergiadou E. Influence of hydrophyte abundance on the spatial distribution of zooplankton in selected lakes in Greece. Hydrobiologia. 2010;656(1):55–65. doi: 10.1007/s10750-010-0435-0. [DOI] [Google Scholar]
- Strid A, Tan K. Flora Hellenica Vol.1. Koeltz Scientific Books; 1997. [Google Scholar]
- Strid A, Tan K. Flora Hellenica Vol.2. Koeltz Scientific Books; 2002. [Google Scholar]
- Touka Anastasia, Vareli Katerina, Igglezou Maria, Monokrousos Nikolaos, Alivertis Dimitrios, Halley John M., Hadjikakou Sotiris, Frillingos Stathis, Sainis Ioannis. Ancient European lakes: Reservoirs of hidden microbial diversity? The case of Lake Pamvotis (NW Greece) Open Journal of Ecology. 2018;08(10):537–578. doi: 10.4236/oje.2018.810033. [DOI] [Google Scholar]
- Tutin T. G., Heywood V. H., Burges N. A., Moore D. M., Valentine D. H., Walters S. M., Webb D. A. Flora Europaea. Volume 1. Lycopodiaceae To Platanaceae. Zenodo. 1964 doi: 10.5281/ZENODO.302862. [DOI]
- Tutin T. G., Heywood V. H., Burges N. A., Moore D. M., Valentine D. H., Walters S. M., Webb D. A. Flora Europaea. Volume 3. Diapensiacea to Myoporaceae. Zenodo. 1972 doi: 10.5281/ZENODO.305475. [DOI]
- Tutin T. G., Heywood V. H., Burges N. A., Moore D. M., Valentine D. H., Walters S. M., Webb D. A. Flora Europaea. Volume 4. Plantaginaceae to Compositae (and Rubiaceae) Zenodo. 1976 doi: 10.5281/ZENODO.293764. [DOI]
- Tutin T. G., Heywood V. H., Burges N. A., Moore D. M., Valentine D. H., Walters S. M., Webb D. A. Flora Europaea. Volume 5. Alismataceae to Orchidaceae (Monocotyledones) Zenodo. 1980 doi: 10.5281/ZENODO.293845. [DOI]
- Tutin T. G., Heywood V. H., Burges N. A., Moore D. M., Valentine D. H., Walters S. M., Webb D. A. Flora Europaea. Volume 2. Rosaceae to Umbelliferae (3rd Edition) Zenodo. 1981 doi: 10.5281/ZENODO.293200. [DOI]
- Tzedakis P. C. Buffered tree population changes in a Quaternary eefugium: Evolutionary implications. Science. 2002;297(5589):2044–2047. doi: 10.1126/science.1073083. [DOI] [PubMed] [Google Scholar]
- Whittaker R. H. Dominance and diversity in land plant communities: Numerical relations of species express the importance of competition in community function and evolution. Science. 1965;147(3655):250–260. doi: 10.1126/science.147.3655.250. [DOI] [PubMed] [Google Scholar]
- Whittaker R J, Fernandez-Palacios J M. Island biogeography. Ecology, evolution and conservation. 2nd Edition. Oxford and New York: Oxford University Press; 2007. [Google Scholar]