

Abstract
The global population of individuals over the age of 65 is growing at an unprecedented rate and is expected to reach 1.6 billion by 2050. Most older individuals are affected by multiple chronic diseases, leading to complex drug treatments and increased risk of physical and cognitive disability. Improving or preserving the health and quality of life of these individuals is challenging due to a lack of well‐established clinical guidelines. Physicians are often forced to engage in cycles of “trial and error” that are centered on palliative treatment of symptoms rather than the root cause, often resulting in dubious outcomes. Recently, geroscience challenged this view, proposing that the underlying biological mechanisms of aging are central to the global increase in susceptibility to disease and disability that occurs with aging. In fact, strong correlations have recently been revealed between health dimensions and phenotypes that are typical of aging, especially with autophagy, mitochondrial function, cellular senescence, and DNA methylation. Current research focuses on measuring the pace of aging to identify individuals who are “aging faster” to test and develop interventions that could prevent or delay the progression of multimorbidity and disability with aging. Understanding how the underlying biological mechanisms of aging connect to and impact longitudinal changes in health trajectories offers a unique opportunity to identify resilience mechanisms, their dynamic changes, and their impact on stress responses. Harnessing how to evoke and control resilience mechanisms in individuals with successful aging could lead to writing a new chapter in human medicine.
Keywords: aging, biological aging, hallmarks of aging, inflammation, multimorbidity, resilience, senescence
Finding a reference metric for the rate of biological aging is key to understanding the molecular nature of the aging process. Defining and validating this metric in humans opens the door to a new kind of medicine that will overcome the limitation of current disease definitions. We will then be able to approach health in a global perspective and bring life course preventative measures to the center of attention.
1. INTRODUCTION
In its most profound essence, resilience is at the core of life and is interpreted as the harmonic assemblage of the biochemical processes that are aimed at maintaining the identity, integrity, and autonomy of individual organisms against the perturbations induced by both internal and external environments. Developmental changes that occur during fetal growth and postnatal development are fast, massive, tightly predetermined, and stereotyped, probably because they are driven by a redundant and self‐correcting genetic “software.” This period of development is followed by a time of relative stability, where changes in physical and cognitive function are small and only detectable by very sensitive tools or challenging tests. During this “middle” period, most individuals in the population are free of diseases (Blekhman et al., 2008; Olshansky, 2016). However, underneath this apparent stability there are several compensatory and homeostatic mechanisms hidden that constantly operate to preserve biochemical balance and prevent phenotypic derangements, as well as functional decline. Early in life, these mechanisms are highly effective and provide a robust homeostasis, but begin to fade later in life, and unrepaired damage accumulates beyond the functional threshold (Figure 1). The extreme variability by which these mechanisms maintain a stable homeostasis explains why the variance of aging phenotypes expands over time, even at extreme old age despite the leveling force of selective mortality. Understanding the nature of these “resilience mechanisms” (homeostatic mechanisms, in green in Figure 1) and “accumulated damages” (entropic forces, in red in Figure 1), as well as finding methods to assess them in humans, is a very active area of investigation. For example, while the condition of “frailty” in older persons is often defined as a “reduction of physiological compensation,” almost all criteria currently proposed are based on measures of damage. Damage only emerges clinically when compensatory mechanisms are exhausted (Ferrucci & Fabbri, 2018). As shown in Figure 1, physical decline, cognitive decline, and frailty may result from two interrelated mechanisms, one inducing and the other preventing damage, which may act separately or jointly. We postulate that the interaction between damage and repair could explain why some individuals are aging “faster” and studying them jointly may point to the mechanisms of accelerated aging.
Figure 1.
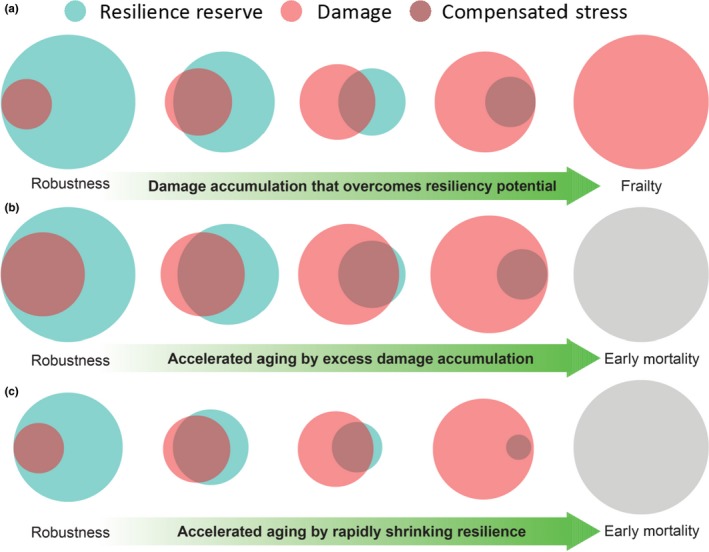
Normal aging (a) and different pathways to accelerated aging (b and c). A. Robust resilience at a young age fully compensates damage. Over time, damage accumulates that is not fully compensated by resilience. Toward the end of life, resiliency is overwhelmed, and new stresses cause fast, unopposed damage accumulation that leads to frailty and eventually to death. Accelerated aging may occur either because of faster rates of damage accumulation (b) or because of rapid shrinking and eventual collapse of resilience (c). Note that even in the state of robustness, damage can be already abnormally high (b) and resilience already abnormally low (c)
Studies in animal models have begun to reveal the nature of these mechanisms, and some assays for humans have been developed. Although many of these unique or composite measures generally track chronological age with a predictable schedule, the biology of their compensatory and homeostatic nature is only partially explained and their relevance for health is limited to observational studies (Hilmer & Le Couteur, 2016; Kirkland, Tchkonia, Zhu, Niedernhofer, & Robbins, 2017; Moreno‐Villanueva et al., 2015; Newman et al., 2016; Niedernhofer, Kirkland, & Ladiges, 2017; Richardson et al., 2015; Robbins & Niedernhofer, 2017).
In the next part of this manuscript, we will try to summarize what measures of aging biology are currently available. We are inspired by two recent articles that outlined the “hallmarks” and the “pillars” of aging (López‐Otín, Blasco, Partridge, Serrano, & Kroemer, 2013; Sierra, 2016), but we purposely limit this description to those measures that can be obtained in humans and we point to their validity, limitations, and potential for further development. For most biomarkers, whether they reflect damage, compensation, or a combination of the two remains unknown.
1.1. Genomic instability
The accumulation of DNA damage (somatic mutations) with age has been proposed as the primary cause of aging because of its effects on the fidelity of proteins and the regulation of gene expression. While mutational load plays a role in carcinogenesis, solid evidence that the accumulation of somatic mutations during normal aging is associated with the phenotypes of aging is lacking. Studies that compare single‐cell and multicellular DNA high‐fidelity sequences and studies that systematically screen for mutation in single cells that are clonally expanded are underway. Spontaneous somatic mutations accumulate in human B lymphocytes, and it has been suggested that they may contribute to functional decline of B lymphocytes in the elderly (Zhang et al., 2019). Similarly, a slight accumulation of DNA somatic mutations with aging has been demonstrated in skeletal muscle satellite cells from human biopsies (Franco et al., 2018), while Bae et al. sequenced DNA from single neurons and demonstrated that somatic mutations accumulate with aging from 4 months to 82 years of age (Bae et al., 2017; Lodato et al., 2017). The functional relevance of these mutations is unknown.
While several biochemical and cell‐based tests of DNA repair capacity have been developed and shown to be reasonably objective and reliable, quantification of DNA repair capacity in humans remains unsatisfactory (Berwick & Vineis, 2005; Trzeciak et al., 2008; Trzeciak, Barnes, & Evans, 2008). The few tests described in the literature have not been applied to large populations and lack independent validation (El‐Zein et al., 2010; Fang, Neutzner, Turtschi, Flammer, & Mozaffarieh, 2015; Hamann & Hartwig, 2014; Holton, Ebenstein, & Gassman, 2018; Nagel et al., 2014; Reddy et al., 2016). Moreover, there is no consensus on gold standard assays and most methods require large amounts of freshly collected pure cell types, and these only address repair capacity of a subset of specific lesions. For example, the comet assay, which quantifies alkaline‐labile sites and/or specific DNA strand breaks, has been used for years, but the reliability and validity of its results have been questioned, partly due to extreme sensitivity to experimental conditions (Collins, 2014; Saha et al., 2008). In addition, some assays require repair of an exogenous substrate, but the substrate design has been proven challenging (Latimer & Kelly, 2014; Reddy et al., 2016; Shen, Fox, Ahn, & Loeb, 2014). DNA somatic mutations accumulation and loss of efficiency of DNA repair mechanisms are likely important drivers of biological aging. However, reliable and valid assays for their quantification should be developed before they can be used in human research and in clinical applications.
1.2. Telomere length
Telomeres are tract of tandem repeats of the six‐nucleotide unit sequence (TTAGGG) that protect chromosome ends from eliciting a DNA damage response. During DNA replication, the DNA polymerases are unable to fully recreate the end of the telomeric DNA and telomeres shorten during each cell division, which ultimately leads to replicative senescence in vitro (Allsopp et al., 1992; Greider, 1998; Herbig, Jobling, Chen, Chen, & Sedivy, 2004). The enzyme telomerase can replenish the lost telomeric DNA, a mechanism that plays a fundamental role in cancer growth, but there is no evidence that telomerase is a resilience mechanism for aging. Telomeres have been proposed to serve as a “molecular clock,” and short telomeres have been hypothesized to contribute to the aging process (Greider, 2010; Saretzki, 2018; Vera, Bernardes de Jesus, Foronda, Flores, & Blasco, 2012; Whittemore, Vera, Martinez‐Nevado, Sanpera, & Blasco, 2019). A 13‐year prospective study in the Baltimore Longitudinal Study of Aging reported that indeed, average telomere length shortens with aging, but the direction and magnitude of change are different in different circulating cells and extremely heterogeneous across individuals, with a substantial percentage of individuals showing average lengthening. Interestingly, significant amounts of telomere shortening were explained by decreased telomerase activity in the cells that express this enzyme, suggesting that measuring telomerase activity in human cells may be informative (Lin et al., 2015). Several reports indicate that short telomeres may be associated with central obesity (García‐Calzón et al., 2013; Mundstock et al., 2015), lifetime accumulation of stress (Epel et al., 2004; Osler, Bendix, Rask, & Rod, 2016; Puterman et al., 2016), increased risk of cardiovascular events (Baragetti et al., 2016; Hammadah et al., 2017), reduced immune response to influenza vaccination (Najarro et al., 2015), mortality (Batsis et al., 2017; Goglin et al., 2016; Heidinger et al., 2012), and several adverse health outcomes (Lin et al., 2015; Lorenzi et al., 2018; Lustig et al., 2017; Sanders & Newman, 2013). Genetic mutations associated with short telomeres have been shown to cause dyskeratosis congenita, pulmonary fibrosis, and several other severe medical conditions that are grouped under the definition of “telomere syndrome” (El‐Chemaly et al., 2018; Ungar et al., 2018). Different methods are available to measure telomere length in circulating cells, including restriction fragment analysis and fluorescence in situ hybridization. Observational studies using these techniques have reported contrasting results, and longitudinal studies have revealed erratic changes over time, possibly due to large measurement error (Berglund et al., 2016; Bischoff et al., 2006; Eerola et al., 2010; Lin et al., 2016; Müezzinler, Zaineddin, & Brenner, 2013; Solomon et al., 2014). Work is underway to establish an optimal “gold standard” assay for epidemiological studies (Behrens et al., 2017; Montpetit et al., 2014). At this stage, there is not enough evidence in the literature to consider measuring telomere shortening as a biological mechanism of aging or telomere length as a biomarker of biological aging. In general, the clinical relevance of measuring telomere length is unclear.
1.3. Cellular senescence
Cellular senescence is a stress response mechanism characterized by replication arrest and complex changes in morphology, chromatin organization, secretome, and expression of typical protein biomarkers (Muñoz‐Espín & Serrano, 2014; Rodier & Campisi, 2011). Conditions that trigger senescence include genomic instability, extreme telomere shortening, metabolic and proteostatic stress, reactive oxidative species (ROS), oncogene activation, mitochondrial dysfunction, epigenetic changes, and other mechanisms that have not been fully clarified (Childs, Durik, Baker, & van Deursen, 2015; Childs et al., 2017; López‐Otín et al., 2013). In general, these conditions trigger a response that activates the tumor suppressor genes p53, p16Ink4a, and p21 that utilize different pathways to induce cell cycle arrest (Hall et al., 2017; Liu et al., 2009). Most studies indicate that senescence‐induced replication arrest acts as a tumor suppression mechanism, but other physiological roles are emerging, including fetal organ development, wound healing, and aging (Baker & Petersen, 2018; Pratsinis, Mavrogonatou, & Kletsas, 2018; Wiley & Campisi, 2016; Zhang, Chen, Liu, Chen, & Liu, 2014). Irrespective of the nature of the senescence trigger, senescent cells develop a “senescence‐associated secretory phenotype” (SASP) and secrete pro‐inflammatory cytokines and chemokines, growth factors, and matrix proteases (Andriani et al., 2016; Coppé, Desprez, Krtolica, & Campisi, 2010; Strzyz, 2016). Notably, senescent cells become resistant to apoptosis and may persist in tissues for many years unless they are removed by the immune system, therefore interfering with tissue repair and regeneration (Kirkland & Tchkonia, 2017). It has been proposed that the accumulation of senescent cells and the negative effects of SASP proteins on intercellular matrix and on progenitor cells cause tissue degeneration and dysfunction, which may be a primary cause of aging and specific age‐related degenerative diseases, such as osteoarthritis, pulmonary fibrosis, atherosclerosis, diabetes, and Alzheimer's disease (Baker & Petersen, 2018; Bhat et al., 2012; Boccardi, Pelini, Ercolani, Ruggiero, & Mecocci, 2015; Diekman et al., 2018; Palmer et al., 2015; Waters et al., 2018). A recent study demonstrated that the number of cells expressing p16Ink4a in biopsy specimens of intramuscular fat was independently correlated with lower muscle strength and worse walking performance (Justice et al., 2018). Although there is clear evidence that the burden of senescence increases with aging in human CD4+ lymphocytes, kidney epithelia, and skin, the quantification of senescence in vivo is complex because, in spite of the defined set of core features, heterogeneous forms of senescence develop according to different triggers and tissues (Koppelstaetter et al., 2008; Liu et al., 2009; Waaijer et al., 2012). Importantly, none of the characteristic biomarkers described above, including p53, p21, senescence‐associated β‐galactosidase, and SASP factors, are specific to senescence, and p16Ink4a is not always present (Biran et al., 2017; Haferkamp et al., 2009; Laberge et al., 2012; Noren Hooten & Evans, 2017; Rodier & Campisi, 2011). Attempts to quantify senescent cell accumulation in humans from blood biomarkers assume that the SASP proteins dispersed in tissues spill over into circulation and may be detected there. Although none of these proteins are specific, jointly they could potentially identify a unique pattern that tracks the global burden of senescence across tissues or perhaps even show some specificity for their tissues of origin (Tanaka et al., 2018). The quantification of senescence in biopsies from different human tissues is an active area of research. Overall, quantification of senescence burden in humans is informative toward assessing biological aging, and measures based on cellular senescence are likely to enter soon into clinical research and practice.
1.4. Epigenetics
The term epigenetics encompasses the ensemble of mechanisms that modulate gene expression programs that adapt to environmental cues and define stable phenotypic characteristics from differentiated cell types (e.g., an adipocyte rather than a neuron). The three major epigenetic operators are DNA methylation, histone modification, and noncoding RNA. Among these three, a growing body of literature emphasizes the role of DNA methylation in aging and age‐related chronic diseases in humans (Gensous et al., 2017; Levine et al., 2018). In part, this is because DNA methylation is easily assessed in circulating cells and is relatively stable over time. In contrast, measuring histone posttranslational modification and noncoding RNA in humans is expensive, time‐consuming, not fully standardized, and amenable to rapid changes over relatively short time periods. In addition, while studies have related histone modifications and microRNA to cell senescence and diseases in animal models, whether these epigenetic mechanisms are drivers of biological aging in humans is uncertain (Bu, Wedel, Cavinato, & Jansen‐Dürr, 2017; Neault, Couteau, Bonneau, De Guire, & Mallette, 2017; Panda, Abdelmohsen, & Gorospe, 2017; Sidler, Kovalchuk, & Kovalchuk, 2017). Biochemically, DNA methylation is the addition of a methyl group to the 5th carbon of the pyrimidine ring of a cytosine (C) base juxtaposed to guanine (G) through a phosphate (p) bond (CpG), thus forming a 5‐methylcytosine (5mC). The presence of 5mC, especially at a promoter site, is generally believed to suppress gene transcription by blocking transcription factors from binding to promoter sequences, but accumulating evidence suggests that many other mechanisms are at play, including the control of transcriptional splicing (Avin, Umbricht, & Zeiger, 2016; Lev Maor, Yearim, & Ast, 2015; Young et al., 2005). DNA methylation is easily assessed in blood cells and tissues using microarrays, pyrosequencing, and whole‐genome bisulfite sequencing methods. As each CpG site can be differentially methylated in different cells, the site‐specific percent methylation of each CpG across the genome can be quantified. The percentage of 5hC at specific CpG sites can be used to derive an “epigenetic clock” that tracks closely with chronological aging (Hannum et al., 2013; Horvath, 2013). Their discovery has been confirmed by many studies across tissues, individuals, and populations, in addition to examining gestational age, and in longitudinal analyses (Horvath, 2013; Knight et al., 2016; Maierhofer et al., 2017; Quach et al., 2017; Sehl, Henry, Storniolo, Ganz, & Horvath, 2017). These findings demonstrate that some of the biological changes that occur with aging are not purely stochastic, but rather follow a predefined pattern that is constant across individuals and populations. Theoretically, the discrepancy between chronological and epigenetic clocks identifies individuals who are biologically older or younger than their chronological age. Consistent with this notion, “epigenetically older” individuals have a higher risk of developing several age‐related diseases and premature mortality for all causes and cardiovascular diseases (Chen et al., 2016; Marioni et al., 2015). In some studies, older epigenetic age has been associated with biomarkers of inflammation, as well as physical and cognitive function (Degerman et al., 2017; Gale et al., 2018; Levine et al., 2018; Ligthart et al., 2016; Marioni et al., 2015; Quach et al., 2017; Spiegel, Sewal, & Rapp, 2014). Unsurprisingly, the effect size for these associations is relatively small. As the CpG methylation sites included in epigenetic clock were selected based on chronological age, “discarded” CpG sites that deviate from chronological age are probably relevant in identifying accelerated or decelerated aging. In addition, most of the selection process of the relevant CpG sites has been cross‐sectional, which could be profoundly biased by secular trends. More recently, a second generation of epigenetic clocks was developed that uses a “phenotypic age” (PhenoAge) index for reference and/or is tuned to cardiovascular risk factors, including smoking (GrimAge), and is strongly predictive of mortality and a cadre of age‐related adverse health outcomes, including disability and dementia (Levine et al., 2018; Lu et al., 2019).
A recent literature suggests that hydroxymethylcytosine (5hmC), an oxidized form of 5‐methylcytosine (5mC) produced by Fe‐dependent dioxygenases named TETs (ten–eleven translocation) during demethylation, is a novel DNA epigenetic modulator with biological roles different from 5mC (Tahiliani et al., 2009). This view has been reinforced by the discovery of proteins showing a binding preference for 5hmC rather than 5mC (Mellen, Ayata, Dewell, Kriaucionis, & Heintz, 2012). Traditional bisulfite‐based assays for DNA methylation cannot distinguish 5mC from 5hmC, but new methods were recently developed for the regional detection and quantification of 5hmC (Szwagierczak, Bultman, Schmidt, Spada, & Leonhardt 2010; Terragni, Bitinaite, Zheng, & Pradhan, 2012). Differently from 5mC, abundance of 5hmC is highly variable across tissues, from less than 0.5% in the blood (Godderis et al., 2015) to close to 13% in the brain (Wen et al., 2014) where it is particularly high in mature neurons. Although the role of 5hmC has not definitively been established, contrary to 5mC that is thought to inhibit gene expression, 5hmC is enriched in coding regions of actively transcribed genes and some studies have shown positive correlations with expression levels (Branco, Ficz, & Reik, 2012; Colquitt, Allen, Barnea, & Lomvardas, 2013; Yu et al., 2012). There is evidence that hydroxymethylation increases with aging in several brain regions, including the hippocampus, while declining in peripheral mononuclear cells (Szulwach et al., 2011, Valentini et al., 2016). Brain hydroxymethylation has also been associated with age‐related neurodegenerative diseases such as Alzheimer's disease (Zhao et al., 2017). Whether information on hydroxymethylation and TET proteins in circulating cells or other tissues provides information on biological aging is unknown and is an active area of research.
The development of epigenetic clocks is based on an agnostic statistical approach because biological mechanisms driving the clock are unknown. When these mechanisms are clarified, tools could be developed that would be even more useful for clinical applications. Also, the extent to which age‐related epigenetic changes can be considered evidence of damage or compensation remains unclear. Based on developmental theories, during the prenatal and early‐life periods, epigenetic mechanisms refine the genetic program to be optimally responsive to present and future environmental challenges. For example, massive epigenetic changes occur when food is scarce, and these changes may remain even when food becomes available later on, contributing to diabetes and metabolic syndrome (Bygren et al., 2014; Jiménez‐Chillarón et al., 2012; Jimenez‐Chillaron et al., 2006; Lorite Mingot, Gesteiro, Bastida, & Sánchez‐Muniz, 2017). The theory of developmental origins of health and disease hypothesizes that these early changes may be adaptive at the time they develop but may become maladaptive in later life, causing chronic diseases (Barker, Osmond, Winter, Margetts, & Simmonds, 1989; Ben‐Shlomo, Cooper, & Kuh, 2016; Pembrey, Saffery, & Bygren, 2014; Wadhwa, Buss, Entringer, & Swanson, 2009). The phasic approach to this theory can be extended to the continuum of the lifespan, and epigenetic changes may be considered as a cluster of predefined adaptive mechanisms that are implemented to counteract the effects of other typical biological changes that occur with aging. The essential elements of this theory are summarized in Figure 2. Research regarding the epigenetic clock clearly demonstrates that methylation in some specific CpG sites is reset at birth, as witnessed by the “zero” epigenetic age of cord blood (Knight et al., 2016). During aging, there is continuous epigenetic tuning of the predefined gene expression in response to environmental stress. This adaptive response, which likely occurs hundreds of times over the life course, may be fully adaptive or lead to negative consequences in subsequent years. Thus, in agreement with the theory of developmental origins of health and disease, over the life course physiological responses to stress are affected by all previous adaptations to stress already encountered, and the readout of this status is an epigenetic signature (Ben‐Shlomo et al., 2016). Thus, “epigenetic acceleration” would mark adaptive epigenetic changes that occur with aging earlier than average because of early imbalances between damaging and resiliency mechanisms. Interestingly, since methylation can be modified, interventions that “slow down” aging, thereby reducing the need for compensatory mechanisms, would also result in younger epigenetic age. Overall, DNA methylation is emerging as one of the most robust biomarkers of “biological aging” and represents a promising area for research that may be translated soon into clinical practice.
Figure 2.
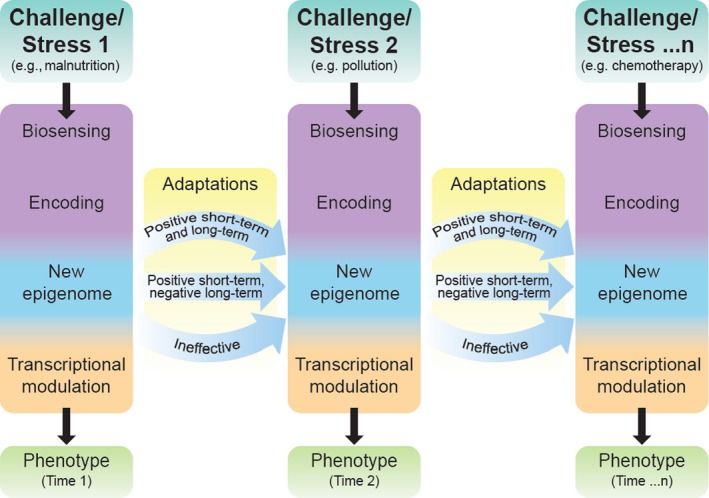
Epigenetic model of continuous transcriptional tuning leading to the aging phenotype. Long‐term adaptation within the lifespan requires epigenetic modulation of the transcriptional machinery. Environmental clues are read by specific biosensors and encoded into epigenetic changes that modulate transcriptional subroutines. The new epigenetic landscape is meant to be adaptive but may fail its purpose and become maladaptive in either the short or long term. Ineffective adaptation/compensation negatively impacts the rate of biological aging and, in turn, phenotypic and functional aging. In the scheme, we show only three cycles of epigenetic adaptation, at any point in time; the epigenetic landscape results from the sum of hundreds or even thousands of adaptive cycles that occur throughout life; and some more relevant than others. Importantly, very little is known about how environmental stresses are sensed and encoded into epigenetic changes
1.5. Mitochondrial function
Mitochondria are organelles found in all human cells, and their primary role is energy production through oxidative phosphorylation. They are also involved in signaling by producing ROS, as well as by regulating cellular metabolism, apoptosis‐programmed cell death, and other functions that are biologically important but cannot be reliably measured in vivo in humans (Gonzalez‐Freire et al., 2015). The mitochondrial theory of aging proposes that accumulation of damage to mitochondria and mitochondrial DNA (mtDNA) induces aging by reducing energy availability and increasing production of ROS that damage macromolecules (Harman, 1956, 1972, 2003). In humans, mitochondrial metabolic function is often studied in vitro in skeletal muscle by respirometry in permeabilized muscle fibers obtained through biopsies, as well as in vivo by phosphorous magnetic resonance spectroscopy (P31 MRS) (Lanza & Nair, 2010). Using both methods, it has been demonstrated that the degree of oxidative phosphorylation declines with aging in humans in the heart, skeletal muscle, and other tissues (Coen et al., 2012; Consolini, Ragone, Bonazzola, & Colareda, 2017; Fabbri et al., 2016; Gonzalez‐Freire et al., 2018; Holloway et al., 2018). Reduced mitochondrial function is associated with mobility decline in older persons, while the effect is mediated by a reduction of muscle strength (Zane et al., 2017). Currently, there are no measures of mitochondrial function in humans that are fully satisfactory. P31 MRS is noninvasive and reliable but is too expensive for large population studies, and this method only measures global skeletal muscle oxidative phosphorylation, which depends not only on the intrinsic mitochondrial function but also on the capacity of circulatory and microcirculatory system to deliver to mitochondria adequate amount of oxygen and oxidative substrates. Muscle biopsies are invasive but safe and allow for a variety of measurements—including direct mitochondrial respiration—as well as a wide range of biochemical assays that target different components of the energetic and biogenesis machinery, and the quantification of characterization of morphological changes using microscopy imaging techniques (Coggan, 1995; Hughes et al., 2015). Many of these indexes have been associated with aging and increased risk of chronic conditions (Consolini et al., 2017; Gonzalez‐Freire et al., 2018). Several assays are available for measuring ROS generation, antioxidant defense, and oxidative damage to macromolecules in blood, cells, and tissues (Dikalov & Harrison, 2014; Starkov, 2010). The significance of these oxidative stress biomarkers for aging is uncertain, as in many cases they have been studies in specific medical conditions and not in the context of aging studies (Al Shahrani, Heales, Hargreaves, & Orford, 2017; Hayashi & Cortopassi, 2015; Weber et al., 2017). Studies that use multiple mitochondrial biomarkers have revealed only a slight intercorrelation between the markers and aging, suggesting that they tap into different biological dimensions (Lara et al., 2015; Larsen et al., 2012; Marrocco, Altieri, & Peluso, 2017; Xia, Chen, McDermott, & Han, 2017). Recent data support the hypothesis that mtDNA copy number and degree of heteroplasmy—assessed in human blood cells and in tissue biopsies—provide information on mitochondrial physiology that is relevant for aging and age‐related diseases (McDermott et al., 2018; Moore et al., 2017; Zhang, Wang, Ye, Picard, & Gu, 2017). Both measurements can be utilized via PCR methods or, more recently, by derivation from genome sequencing data (Ding et al., 2015). High mtDNA copy number is considered to be a proxy measure of mitochondrial volume/function, and high mtDNA copy number in blood is associated with better health and survival among older persons, but the direction of this association may be reversed in certain conditions, such as diabetes (Mengel‐From et al., 2014; Moore et al., 2017). Humans have detectable levels of mtDNA‐acquired point mutations in circulating cells and whole blood and, notably, the burden of mutations increase with aging even when measured in inducible pluripotent stem cells (Kang et al., 2016; Qian et al., 2017). Many of these mutations are haploinsufficient or recessive and, when they reach a critical threshold of accumulation, can contribute to declining health in late life (Larsson, 2010; Wachsmuth, Hübner, Li, Madea, & Stoneking, 2016). Measures of mitochondrial physiology and function are powerful biomarkers of biological aging. However, they require extremely careful standardization. In particular, blood measurements may be affected by changes in circulating cells and high levels of mtDNA copy number can also indicate chronic tissue hypoxia (Eirin et al., 2016).
1.6. Proteostasis
The repair, recycling, and elimination of damaged macromolecules/organelles have emerged as key processes in maintaining cell integrity and function (Cuervo et al., 2005; Cuervo, Wong, & Martinez‐Vicente, 2010). These complex goals are accomplished through different mechanisms, such as chaperon‐dependent and chaperon‐independent autophagy, as well as protein biogenesis, folding, trafficking, and degradation (including proteasomal degradation; Kaushik & Cuervo, 2018; Morimoto & Cuervo, 2014; Wong & Cuervo, 2010). In animal models, autophagy and proteostasis become dysfunctional with aging. Rapamycin is an immunosuppressor that extends mammalian lifespans by inhibiting mTOR and stimulating autophagy. Genetic variants within core autophagy genes have been identified that contribute to human diseases, including hereditary spastic paraparesis, Parkinson's disease, and lysosomal storage disorders (Li et al., 2017; Settembre, Fraldi, Rubinsztein, & Ballabio, 2008; Wang et al., 2017). Beyond hereditary disease, evidence is emerging that autophagy becomes defective with aging and contributes to immunosenescence (Cuervo & Macian, 2014; Zhang, Puleston, & Simon, 2016). Accordingly, pretreatment with rapamycin analogs that inhibit TORC1 enhances immune function and reduces infections in the elderly (Mannick et al., 2014; Shavlakadze et al., 2018). Whether rapamycin or rapamycin analogs have potential for improving healthspan and lifespan in humans is unclear, and their potential side effects are of significant concern. Rapamycin analogs that selectively target TORC1, which should have less side effects, have been proposed for treatment of diseases of aging (Arriola Apelo & Lamming, 2016; Bjedov et al., 2010; Chi et al., 2015; El‐Chemaly et al., 2017; Miller et al., 2010).
Other compounds that modulate autophagy have shown anti‐aging properties, including the polyamine spermidine, the natural polyphenol resveratrol, and the gut bacterial product urolithin A. Tissue levels of spermidine decline with age in model organisms and in humans, although they are unusually high in healthy nona/centenarians (Eisenberg et al., 2009; Gupta et al., 2013; Pucciarelli et al., 2012). Spermidine administration increases lifespan and healthspan of multicellular model organisms, at least in part though TORC1 inhibition and enhancement of autophagy. Indeed, blockage of autophagy removes most positive effects of spermidine (Madeo, Eisenberg, Pietrocola, & Kroemer, 2018). Several lines of research suggest that resveratrol enhances autophagy and, through this mechanism, protects against multiple age‐related chronic diseases and increases longevity in mice on a high‐fat diet (Agarwal & Baur, 2011). Mechanisms by which resveratrol induces autophagy are still not fully elucidated but certainly involve both mTOR inhibition and histone deacetylation through the AMPK/SIRT1 signaling pathway (Lee et al., 2008). Interestingly, the combination of spermidine and resveratrol shows synergistic effects on autophagy induction (Morselli et al., 2011).
Urolithin A is a metabolite produced by gut microbiota from compounds found in many fruits and vegetables. Urolithin A has been shown to induce mitophagy in cell cultures, increase longevity in nematodes, and prevent age‐related muscle impairment in mouse models (Ryu et al., 2016). Administration of urolithin A in healthy, sedentary elderly individuals is followed by changes in muscle mitochondrial gene expression that are suggestive of improved mitochondrial and cellular health (Andreux et al., 2019).
Developing assays for autophagy in humans is challenging. Static measures of autophagosome accumulation based on quantification of LC3, an antigen that is only present in autophagosomes, are relatively simple, yet notoriously unreliable. In contrast, measures that track the dynamic flux of the autophagic process, by quantifying accumulation of autophagosome cargo upon inhibition of lysosomal proteolysis, are more reliable and the only suitable assay to discriminate whether increase abundance of autophagosomes is due to increase biogenesis or to reduced clearance (Klionsky, 2014; Yoshii & Mizushima, 2017). Recent studies suggest that adequate quantification of autophagy requires multiple approaches, most of which are expensive, labor‐intensive, and low‐throughput. Thus far, only a few studies provide evidence that autophagy becomes dysfunctional with aging, and a recent study shows that autophagy appears to be better maintained in members of families with extended longevity and positively correlates with improved T‐cell function (Raz et al., 2017). Similarly, no high‐throughput method is available for assessing proteostasis. Recently, a new measure has been developed that uses tetraphenylethene, a fluorescent dye, to label the free cysteine thiols that are normally hidden in the core of properly folded globular proteins and are uncovered by misfolding (Chen et al., 2017). Also, using prolonged starvation in human volunteers, Pietrocola et al. developed a method to assess autophagy in circulating leukocytes. They could detect enhanced autophagic flux in human neutrophils cultured in the presence or absence of leupeptin (Pietrocola et al., 2017). Although these methods are promising, further development and validation in human cells is needed before these assays can be used in clinical studies. Overall, mechanisms that handle repair, recycling, and eliminating damaged macromolecules/organelles could act as strong biomarkers of biological aging and would be extremely useful in clinical application, but better assessment methods need to be developed.
1.7. Stem cell exhaustion, deregulated nutrient sensing, and altered intercellular communication
These three “hallmarks of aging” have been combined in this section because their impact on age‐related diseases, healthspan, and longevity in humans has not been sufficiently characterized. Stem cell exhaustion has been postulated to play a primary role in aging as it interferes with self‐renewal of differentiated cells in tissues and organs, slowly curtailing function (Ren, Ocampo, Liu, & Izpisua Belmonte, 2017). Small cross‐sectional studies have provided some evidence that hematopoietic stem cells in humans accumulate DNA damage, possibly leading to reduced proliferative potential (Beerman, 2017; de Haan & Lazare, 2017). Studying the effect of aging on stem cells in humans is difficult. Hematopoietic stem cells, satellite cells, and epidermal stem cells represent the only easily accessible material, but their isolation is still problematic and only yields small quantities (Ahmadbeigi et al., 2013; Hinken & Billin, 2018; Lavker & Sun, 2000; Liu, Cheung, Charville, & Rando, 2015; Moestrup, Andersen, & Jensen, 2017; Rossi, Challen, Sirin, Lin, & Goodell, 2011). Overall, despite the great enthusiasm for using stem cells to treat many age‐related disease, data on changes with human aging in stem cell numbers, characteristics, and replication potential are still limited (Dexheimer, Mueller, Braatz, & Richter, 2011; Eichler et al., 2017; Fan et al., 2010; Golpanian et al., 2017, 2016; Hare et al., 2012; Jim et al., 2016; Li, Chen, Han, & Fu, 2010; Pang et al., 2011; Rigotti et al., 2016; Tompkins et al., 2017; Volarevic et al., 2011, 2018; Zhang et al., 2011). Understanding whether changes in stem cells biology are important for aging remains an important and promising question, and research in this field is warranted.
Nutrient sensing in humans is important for aging and longevity based on the extraordinary effectiveness of caloric restriction in increasing longevity and healthspan in animal models, including mammals (Anderson, Le Couteur, & de Cabo, 2017). Whether this concept can be transformed into empirical measures in humans remains to be elucidated. Similarly, the concept of “intercellular communication” is so generic as to encompass almost any known physiological mechanism. This concept will be revisited when discussing “inflammation,” which may be a special case of “intercellular communication” that is dysregulated with aging and predicts several adverse health outcomes in humans, as well as multimorbidity (Bektas, Schurman, Sen, & Ferrucci, 2018; Fabbri et al., 2014; Friedman, Christ, & Mroczek, 2015; Sanada et al., 2018).
2. CONNECTING THE BIOLOGY OF AGING WITH AGE‐ASSOCIATED MULTIMORBIDITY
Based on information in the section above, developing a proxy measure of biological aging for humans still requires work but is a very dynamic and promising area of investigation with strong potential for translation. Some of the measures described—namely mitochondrial function, DNA methylation, and, to a lesser extent, cellular senescence and autophagy—are ready to be implemented based on several epidemiological studies, although refinements are always possible (Capri et al., 2015; Choi et al., 2016; Cohen, Morissette‐Thomas, Ferrucci, & Fried, 2016; Jylhävä, Pedersen, & Hägg, 2017; Jylhävä et al., 2014; Kananen et al., 2016; Kent & Fitzgerald, 2016; Kim & Jazwinski, 2015; Levine et al., 2018; Li et al., 2018; Marioni et al., 2019; Marttila et al., 2015; Putin et al., 2017; Sillanpää et al., 2018). Measures of telomere length are hampered by noise and wide longitudinal variations that cannot be explained by health events and at this stage are not useful for measuring biological age (Arai et al., 2015; Jodczyk, Fergusson, Horwood, Pearson, & Kennedy, 2014; Tomaska & Nosek, 2009). New methods are being developed, some of which are focused on detecting the DNA damage response (a typical marker of critical telomere shortening) may yield better results (Choi, Kim, Kim, Kemp, & Sancar, 2015; Hewitt et al., 2012; Rossiello et al., 2017). Senescence has been studied successfully in T lymphocytes, skin, and intramuscular fat, and high‐throughput methods will be available soon (Evangelou et al., 2016; Lozano‐Torres et al., 2017). In addition, specific patterns of circulating proteins may exist that indirectly estimate the burden of senescence (Angelini et al., 2017; Hoffman, Lyu, Pletcher, & Promislow, 2017; Kadota et al., 2018; Menni et al., 2014; Tanaka et al., 2018; Yousefzadeh et al., 2017). Similarly, measures of autophagy are routinely used in mammalian studies and should be applicable to humans (Klionsky, 2014; Klionsky, Cuervo, & Seglen, 2007; Menzies, Moreau, Puri, Renna, & Rubinsztein, 2012). For the other hallmarks, the development of a reliable and valid test is less advanced and will take time.
Multiple lines of evidence suggest that the measures listed above are associated with the severity of multimorbidity but, except for the epigenetic clock, this association has not yet been clearly established. Logically, none of the measures described above represent an exhaustive measure of biological aging and, therefore, new aggregate measures are needed that leverage differences and complementarities of the various biomarkers. To accomplish these goals, the hallmarks of aging should be assessed in a group of individuals that is reasonably sized and enough dispersed across the lifespan to represent the variability of biological age in the general population. Initially, it will be important to evaluate the intercorrelation between these measures, as there is currently evidence that the hallmarks of aging are interconnected (Figure 3). Each numbered arrow in the left portion of Figure 3 refers to a piece of evidence that failure of a certain mechanism leads to impairment in others, a notion that is strengthened by emerging evidence in recent literature, although most is derived from animal studies (Acosta et al., 2013; Chang et al., 2015; Childs, Li, & van Deursen, 2018; García‐Prat et al., 2016; Gonzales‐Ebsen, Gregersen, & Olsen, 2017; Hall et al., 2017; Herranz & Gil, 2018; Kang et al., 2015; Ligthart et al., 2016; Mills, Kelly, & O'Neill, 2017; Moreno‐Blas, Gorostieta‐Salas, & Castro‐Obregón, 2018; Netea‐Maier, Plantinga, Veerdonk, Smit, & Netea, 2015; Wiley et al., 2016). Clarity is needed in determining if the hallmarks of aging are multifaceted expressions of the same core process or if they evolved independently, as interventions would either have to target each single mechanism or could address one mechanism with synergistic benefits on the others. A simple cross‐sectional correlation may not be optimal, as different manifestations of biological aging may occur according to different time schedules, some mechanism preceding others (Ferrucci, Levine, Kuo, & Simonsick, 2018). Thus, these measures needed to be examined using exploratory “lagged analysis” in a longitudinal perspective. Interestingly, all of the “hallmarks of aging” cited above directly or indirectly cause an inflammatory state, suggesting that the pro‐inflammatory state observed in many older persons may reflect the burden of biological aging (Ferrucci et al., 2005; Franceschi & Campisi, 2014; Fulop et al., 2018). Consistent with this hypothesis, inflammation measured by circulating levels of IL‐6 is the only known cross‐sectional and longitudinal predictor of multimorbidity and one of the strongest predictors of incident mobility loss and disability in activities of daily living (Fabbri et al., 2014; Ferrucci et al., 1999, 2002). Mobility loss, disability, and mortality could be used as reference outcomes to calibrate an index of biological aging as a weighted aggregated, predictive measure. However, while the “functional” outcomes are critical for quality of life in the elderly, they occur late in life and fail to capture the initial changes of biological aging at younger ages. Focusing on multimorbidity is a very promising approach, especially as the pace of biological aging and the development of subclinical pathologies are the primary forces behind increased susceptibility to disease (Fabbri et al., 2014). The rate of aging translates into different patterns of multimorbidity due to specific combinations of genetic susceptibility and environmental stress (Figure 3). Finally, as aging is a dynamic construct, the strength of any index of biological aging should be validated longitudinally by demonstrating that the accelerated progression of “biological aging” is paralleled by an accelerated deterioration in the phenotypic and functional dimensions of aging.
Figure 3.
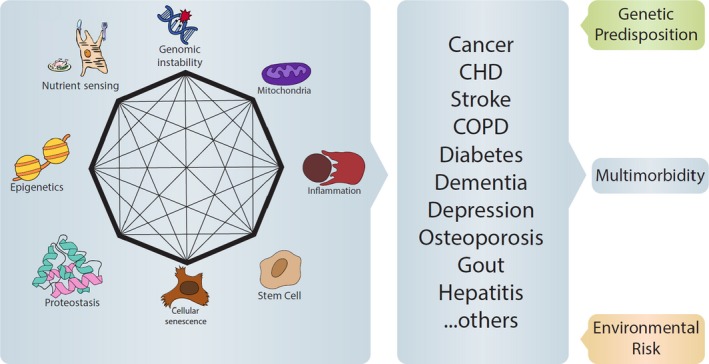
The hallmarks of aging are specific biological mechanisms that drive the rate of biological aging. Emerging research reveals that these different mechanisms are strongly interconnected and, therefore, impairment in one mechanism involves the others. In the figure, the octagon and lines within represent evidence for connections between the different mechanisms. The evidence reported is not exhaustive of the literature connecting the hallmarks. According to the geroscience hypothesis, failure in this network of homeostatic mechanisms affects the pace of aging and, in turn, causes a growing susceptibility to diseases. The specific combination of coexisting diseases that occur in each individual depends on their genetic background, as well as exposure to environmental and behavioral risk factors. The resulting multimorbidity is a major cause of disability. Notably, if the number of coexisting diseases is a major proxy biomarker of the pace of aging, it is unsurprising that the number of diseases rather than specific combination is the strongest risk factor for disability
3. THE TRANSLATIONAL VALUE OF ASSESSING BIOLOGICAL AGING
Substantial investment is necessary to develop an estimator of biological aging that is robust, precise, reliable, and sensitive to change. Thus, a fair question is whether such a titanic project is worth the effort and cost. The answer is YES, without hesitation. Developing an index of biological aging is perhaps the most critical milestone required to advance the field of aging research and, especially, to bring relieve from the burden of multimorbidity and disability in an expanding aging population. Ideally, these measures would be obtained by running tests using blood samples without performing a biopsy, preferably quickly and at low cost. An index of biological aging could be used to empirically address the geroscience hypothesis: “Is biological aging is the cause of the global susceptibility to disease with aging.” Data collected longitudinally—ideally in a life course epidemiological study—could then be used to test if individuals that accumulate coexisting diseases faster than in the general population also have accelerated biological aging. Similarly, these data could be used to test if individuals who are biologically “older,” independent of chronological age, are at a higher risk of developing different medical or functional conditions that do not share physiological mechanisms. Once validated, the fundamental basis of biological aging can be used to probe deeper into questions related to the mechanisms of aging, such as the following: Are there genetic traits that are associated with faster or slower biological aging? Are there “hallmarks” that are better at capturing biological aging at different stages of life?
These questions have immense relevance for geriatric medicine. Despite the rising emphasis on prevention, most current medical care is dedicated to diagnosing and managing diseases that are already symptomatic, which does not address the underlying issues related to geriatric health conditions. By understanding the intrinsic mechanisms of biological aging, including damage and resilience, medical professional will be able to best orient and prescribe therapeutic choices. These mechanisms are summarized in Table 1 according to the current state of knowledge. The first column lists measures of damage for each one of the hallmarks of aging, the second lists the compensatory measures that we would like to have available, and the third lists the compensatory measures that are currently available. Clearly, the current ability to measure biological compensations and resilience is very limited, although most are vital to human health. In fact, it has been proposed that chronic diseases, especially those that emerge in old age, may be cross‐classified based on their dependence on the force of the “noxa patogena” and the robustness of resilience.
Table 1.
Biomarkers of “damage” and “compensation” for the different hallmarks of aging
| Hallmark | Damage | Resilience (compensation) response | Measures |
|---|---|---|---|
| Genomic instability |
|
|
|
| Telomere shortening |
|
|
|
| Cellular senescence |
|
|
|
| Epigenetic changes |
|
|
|
| Mitochondrial dysfunction |
|
|
|
| Decreased autophagy, proteostasis |
|
|
|
| Stem cell exhaustion |
|
|
|
The second column lists measures of damage, some of which are already feasible in humans, while others are only theoretically feasible. The third column lists measures of resilience that would be theoretically desirable, while the fourth column lists measures that are currently feasible. Importantly, regarding many of the available measures, understanding if they reflect damage or compensation requires further investigation.
Abbreviations: NGS, new‐generation sequencing; SASP, senescence‐associated secretory phenotype; TEM, transmission electron microscopy.
The approach described above is not too farfetched from our experience. Hopefully, we all take good care of our cars before they break or malfunction; we make sure that the water an oil levels are ok, that the brake pads are not consumed, that the pressure in the tires is according to factory recommendations. We carefully follow maintenance schedule because we want to maximize the healthy life of our cars and avoid expensive repairs and replacements. Shouldn't we pay the same attention to our bodies? In the field of geriatrics, the situation is even more extreme and often patients come to the clinic when they are already affected by multiple diseases, have lost their autonomy, and have economic and social constrains. In other words, they come to observation when all the mechanisms of compensation and resilience are exhausted. Despite these odds, geriatricians sometime make miracles, but certainly not often enough. The possibility of measuring biological aging swaps this perspective and allows the assessment of health status at a time when our physiology is still resilient, there are still no symptoms, and interventions are more likely to be effective.
A robust biomarker of biological aging would have benefits beyond the early identification of persons who age “faster” than others. First, the genetic, environmental, and behavioral risk factors associated with accelerated aging could be identified. Then, longitudinal studies could be utilized to identify specific time points at which the trajectories of aging change and relate to those other health‐related triggers, such as the exposure to pollution associated with moving to a different city. As biological aging is the primary cause of resilience loss, measuring damage and compensation may help in determining between interventions with potentially serious side effects. Longitudinally, a marker of aging could be used to track if interventions with similar efficacy toward a specific target affect the “speed of aging” differently, which may impact accelerated declines in health. This approach could be used to both refine choices in alternative therapies and develop new medications in order to avoid damage accumulation or curtail compensatory mechanisms. Clinical trials then can be designed to specifically target the speed of aging, the underlying causes of multimorbidity, or both as the primary outcomes of interest. The list of interventions is almost limitless, even without considering the many other applications that are currently unknown and will only become evident as the field progresses.
4. MULTIMORBIDITY AND THE ART OF GERIATRICIANS
A primary focus in geriatric medical is the management of patients affected by multiple coexisting, chronic diseases, as well as physical and cognitive impairments. Indeed, geriatric patients typically have a long list of diagnoses, prescriptions, impairments, social problems, and financial constraints, often presenting a medical dilemma with no clear solution. Most clinical guidelines focus on one disease and, in only exceptional and recent cases, on diseases that belong to the same organ system (Jani et al., 2017; Moreno, Mangione, Kimbro, & Vaisberg, 2013; Spaak, 2017). This is in spite of the fact that co‐occurrence of two or more chronic diseases is the most prevalent medical condition in persons 65 or older (Cesari, Pérez‐Zepeda, & Marzetti, 2017; Fabbri et al., 2015; Guiding Principles for the Care of Older Adults with Multimorbidity: An Approach for Clinicians, 2012; Tisminetzky et al., 2017; Vetrano et al., 2017). Daily, geriatricians are faced with overwhelming complexity, requiring powerful tools: an exhaustive knowledge of medicine and physiology, the ability to evaluate from a list of diseases to choose from possible therapies, and a strong focus on quality of life and on patient preferences. Unfortunately, they are limited with little undersatnding of the biological basis for aging. If multimorbidity is a stochastic assemblage of separate pathologies, the resulting number of syndromes exceeds any serious attempt at classification, which is an essential prerequisite for tailored interventions. Thus, caring for older patients becomes a cyclic process, involving a sequence of trials and errors that are driven by a mixture of knowledge, experience, and intuition.
5. MULTIMORBIDITY AS AN EXPRESSION OF BIOLOGICAL AGING
The emerging field of geroscience presents a hopeful approach to multimorbidity, which aims to understand the relationship between biological aging and age‐related diseases at the molecular level. The traditional approach to studying of aging is rooted in a clear‐cut distinction between aging and diseases, while the geroscience paradigm intimately connects the molecular mechanisms of aging with the rising susceptibility to diseases. This may explain why the number of coexistent chronic diseases tends to increase geometrically with aging in both men and women (Fabbri et al., 2015; GBD, 2016 Disease, & Injury Incidence & Prevalence Collaborators, 2016; Guiding Principles for the Care of Older Adults with Multimorbidity: An Approach for Clinicians, 2012; He et al., 2018; Melis, Marengoni, Angleman, & Fratiglioni, 2014; Niccoli & Partridge, 2012; Rae et al., 2010; Rocca et al., 2014; St Sauver et al., 2015). This conceptual shift on the origin of age‐related multimorbidity opens new, previously unexplored opportunities for research and clinical care in older persons. Importantly, if the core mechanisms of aging can be identified, they could be targeted for interventions aimed at preventing multimorbidity and disability, while also improving the quality of life in old age.
To explain the development of this new science, the conceptual paradigm of geroscience needs to be fully explored. Time is the most “robust” and “precise” metric of aging; however, the chronological dimension presents intrinsic problems due to the magnitude of anatomical and physiological changes that occur with aging in a single time unit (e.g., one year), which can be quite heterogeneous.
6. CONCLUDING REMARKS
Progress in research is not linear. Periods characterized by rates of incremental knowledge are interlaced with “eureka” moments as milestone discoveries suddenly open new possibilities that thrust research and knowledge to a higher level. Galileo's use of the telescope to explore the stars, Kary Mullis's description of polymerase chain reaction, and Edwin Hubble's demonstration that the universe is expanding are just few examples of these moments. The field of aging research is living one of those magical moments. Finding a reference metric for the rate of biological aging is key to understanding the molecular nature of the aging process. Defining and validating this metric in humans opens the door to a new kind of medicine that will overcome the limitation of current disease definitions, approaching health in a global perspective and bringing life course preventative measures to the center of attention.
CONFLICT OF INTEREST
None.
ACKNOWLEDGMENTS
This work was supported by the National Institutes of Health (NIH) and the Intramural Research Program (IRP), National Institute on Aging (NIA). The authors would like to thank Drs. Ana Maria Cuervo, Judith Campisi (especially in the compilation of Table 1), Nan Ping Weng, David Wilson, Yi Liu, Myriam Gorospe, and Isabel Beerman for their comments and suggestions that substantially improved the quality of this manuscript.
Ferrucci L, Gonzalez‐Freire M, Fabbri E, et al. Measuring biological aging in humans: A quest. Aging Cell. 2020;19:e13080 10.1111/acel.13080
This article has been contributed to by US Government employees and their work is in the public domain in the USA.
REFERENCES
- Acosta, J. C. , Banito, A. , Wuestefeld, T. , Georgilis, A. , Janich, P. , Morton, J. P. , … Gil, J. (2013). A complex secretory program orchestrated by the inflammasome controls paracrine senescence. Nature Cell Biology, 15(8), 978–990. 10.1038/ncb2784 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Agarwal, B. , & Baur, J. A. (2011). Resveratrol and life extension. Annals of the New York Academy of Sciences, 1215, 138–143. 10.1111/j.1749-6632.2010.05850.x [DOI] [PubMed] [Google Scholar]
- Ahmadbeigi, N. , Soleimani, M. , Vasei, M. , Gheisari, Y. , Mortazavi, Y. , Azadmanesh, K. , … Nardi, N. B. (2013). Isolation, characterization, and transplantation of bone marrow‐derived cell components with hematopoietic stem cell niche properties. Stem Cells and Development, 22(23), 3052–3061. 10.1089/scd.2013.0005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Al Shahrani, M. , Heales, S. , Hargreaves, I. , & Orford, M. (2017). Oxidative stress: Mechanistic insights into inherited mitochondrial disorders and Parkinson’s disease. Journal of Clinical Medicine, 6(11), 100 10.3390/jcm6110100 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Allsopp, R. C. , Vaziri, H. , Patterson, C. , Goldstein, S. , Younglai, E. V. , Futcher, A. B. , … Harley, C. B. (1992). Telomere length predicts replicative capacity of human fibroblasts. Proceedings of the National Academy of Sciences, 89(21), 10114–10118. 10.1073/pnas.89.21.10114 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anderson, R. M. , Le Couteur, D. G. , & de Cabo, R. (2017). Caloric restriction research: New perspectives on the biology of aging. Journals of Gerontology. Series A, Biological Sciences and Medical Sciences, 73(1), 1–3. 10.1093/gerona/glx212 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Andreux, P. A. , Blanco‐Bose, W. , Ryu, D. , Burdet, F. , Ibberson, M. , Aebischer, P. , … Rinsch, C. (2019). The mitophagy activator urolithin A is safe and induces a molecular signature of improved mitochondrial and cellular health in humans. Nature Metabolism, 1(6), 595–603. 10.1038/s42255-019-0073-4 [DOI] [PubMed] [Google Scholar]
- Andriani, G. A. , Almeida, V. P. , Faggioli, F. , Mauro, M. , Tsai, W. L. , Santambrogio, L. , … Montagna, C. (2016). Whole chromosome instability induces senescence and promotes SASP. Scientific Reports, 6(1), 35218 10.1038/srep35218 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Angelini, F. , Pagano, F. , Bordin, A. , Picchio, V. , De Falco, E. , & Chimenti, I. (2017). Getting old through the blood: Circulating molecules in aging and senescence of cardiovascular regenerative cells. Frontiers in Cardiovascular Medicine, 4, 62 10.3389/fcvm.2017.00062 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arai, Y. , Martin‐Ruiz, C. M. , Takayama, M. , Abe, Y. , Takebayashi, T. , Koyasu, S. , … von Zglinicki, T. (2015). Inflammation, but not telomere length, predicts successful ageing at extreme old age: A longitudinal study of semi‐supercentenarians. EBioMedicine, 2(10), 1549–1558. 10.1016/j.ebiom.2015.07.029 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arriola Apelo, S. I. , & Lamming, D. W. (2016). Rapamycin: An InhibiTOR of aging emerges from the soil of Easter Island. Journals of Gerontology. Series A, Biological Sciences and Medical Sciences, 71(7), 841–849. 10.1093/gerona/glw090 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Avin, B. A. , Umbricht, C. B. , & Zeiger, M. A. (2016). Human telomerase reverse transcriptase regulation by DNA methylation, transcription factor binding and alternative splicing (Review). International Journal of Oncology, 49(6), 2199–2205. 10.3892/ijo.2016.3743 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bae, T. , Tomasini, L. , Mariani, J. , Zhou, B. , Roychowdhury, T. , Franjic, D. , … Vaccarino, F. M. (2017). Different mutational rates and mechanisms in human cells at pregastrulation and neurogenesis. Science, 359(6375), 550–555. 10.1126/science.aan8690 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baker, D. J. , & Petersen, R. C. (2018). Cellular senescence in brain aging and neurodegenerative diseases: Evidence and perspectives. Journal of Clinical Investigation, 128(4), 1208–1216. 10.1172/jci95145 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baragetti, A. , Palmen, J. , Garlaschelli, K. , Grigore, L. , Humphries, S. , Catapano, A. L. , … Giuseppe Danilo, N. (2016). Leukocyte telomere length, genetically determined, is causally associated with the progression of carotid Intima‐Media Thickness and incidence of cardiovascular events. Atherosclerosis, 252, e252 10.1016/j.atherosclerosis.2016.07.064 [DOI] [Google Scholar]
- Barker, D. J. P. , Osmond, C. , Winter, P. D. , Margetts, B. , & Simmonds, S. J. (1989). Weight in infancy and death from ischaemic heart disease. The Lancet, 334(8663), 577–580. 10.1016/s0140-6736(89)90710-1 [DOI] [PubMed] [Google Scholar]
- Batsis, J. A. , Mackenzie, T. A. , Vasquez, E. , Germain, C. M. , Emeny, R. T. , Rippberger, P. , … Bartels, S. J. (2017). Association of adiposity, telomere length and mortality: Data from the NHANES 1999–2002. International Journal of Obesity, 42(2), 198–204. 10.1038/ijo.2017.202 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beerman, I. (2017). Accumulation of DNA damage in the aged hematopoietic stem cell compartment. Seminars in Hematology, 54(1), 12–18. 10.1053/j.seminhematol.2016.11.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Behrens, Y. L. , Thomay, K. , Hagedorn, M. , Ebersold, J. , Henrich, L. , Nustede, R. , … Göhring, G. (2017). Comparison of different methods for telomere length measurement in whole blood and blood cell subsets: Recommendations for telomere length measurement in hematological diseases. Genes, Chromosomes and Cancer, 56(9), 700–708. 10.1002/gcc.22475 [DOI] [PubMed] [Google Scholar]
- Bektas, A. , Schurman, S. H. , Sen, R. , & Ferrucci, L. (2018). Aging, inflammation and the environment. Experimental Gerontology, 105, 10–18. 10.1016/j.exger.2017.12.015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ben‐Shlomo, Y. , Cooper, R. , & Kuh, D. (2016). The last two decades of life course epidemiology, and its relevance for research on ageing. International Journal of Epidemiology, 45(4), 973–988. 10.1093/ije/dyw096 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berglund, K. , Reynolds, C. A. , Ploner, A. , Gerritsen, L. , Hovatta, I. , Pedersen, N. L. , & Hägg, S. (2016). Longitudinal decline of leukocyte telomere length in old age and the association with sex and genetic risk. Aging, 8(7), 1398–1415. 10.18632/aging.100995 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berwick, M. , & Vineis, P. (2005). Measuring DNA repair capacity: Small steps. Journal of the National Cancer Institute, 97(2), 84–85. 10.1093/jnci/dji038 [DOI] [PubMed] [Google Scholar]
- Bhat, R. , Crowe, E. P. , Bitto, A. , Moh, M. , Katsetos, C. D. , Garcia, F. U. , … Torres, C. (2012). Astrocyte senescence as a component of Alzheimer’s disease. PLoS ONE, 7(9), e45069 10.1371/journal.pone.0045069 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Biran, A. , Zada, L. , Abou Karam, P. , Vadai, E. , Roitman, L. , Ovadya, Y. , … Krizhanovsky, V. (2017). Quantitative identification of senescent cells in aging and disease. Aging Cell, 16(4), 661–671. 10.1111/acel.12592 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bischoff, C. , Petersen, H. C. , Graakjaer, J. , Andersen‐Ranberg, K. , Vaupel, J. W. , Bohr, V. A. , … Christensen, K. (2006). No association between telomere length and survival among the elderly and oldest old. Epidemiology, 17(2), 190–194. 10.1097/01.ede.0000199436.55248.10 [DOI] [PubMed] [Google Scholar]
- Bjedov, I. , Toivonen, J. M. , Kerr, F. , Slack, C. , Jacobson, J. , Foley, A. , & Partridge, L. (2010). Mechanisms of life span extension by rapamycin in the fruit fly Drosophila melanogaster . Cell Metabolism, 11(1), 35–46. 10.1016/j.cmet.2009.11.010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blekhman, R. , Man, O. , Herrmann, L. , Boyko, A. R. , Indap, A. , Kosiol, C. , … Przeworski, M. (2008). Natural selection on genes that underlie human disease susceptibility. Current Biology, 18(12), 883–889. 10.1016/j.cub.2008.04.074 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boccardi, V. , Pelini, L. , Ercolani, S. , Ruggiero, C. , & Mecocci, P. (2015). From cellular senescence to Alzheimer’s disease: The role of telomere shortening. Ageing Research Reviews, 22, 1–8. 10.1016/j.arr.2015.04.003 [DOI] [PubMed] [Google Scholar]
- Branco, M. R. , Ficz, G. , & Reik, W. (2012). Uncovering the role of 5-hydroxymethylcytosine in the epigenome. Nature Reviews Genetics, 13(1), 7–13. 10.1038/nrg3080 [DOI] [PubMed] [Google Scholar]
- Bu, H. , Wedel, S. , Cavinato, M. , & Jansen‐Dürr, P. (2017). MicroRNA regulation of oxidative stress‐induced cellular senescence. Oxidative Medicine and Cellular Longevity, 2017, 1–12. 10.1155/2017/2398696 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bygren, L. , Tinghög, P. , Carstensen, J. , Edvinsson, S. , Kaati, G. , Pembrey, M. E. , & Sjöström, M. (2014). Change in paternal grandmothers' early food supply influenced cardiovascular mortality of the female grandchildren. BMC Genetics, 15(1), 12 10.1186/1471-2156-15-12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Capri, M. , Moreno‐Villanueva, M. , Cevenini, E. , Pini, E. , Scurti, M. , Borelli, V. , … Franceschi, C. (2015). MARK‐AGE population: From the human model to new insights. Mechanisms of Ageing and Development, 151, 13–17. 10.1016/j.mad.2015.03.010 [DOI] [PubMed] [Google Scholar]
- Cesari, M. , Pérez‐Zepeda, M. U. , & Marzetti, E. (2017). Frailty and multimorbidity: Different ways of thinking about geriatrics. Journal of the American Medical Directors Association, 18(4), 361–364. 10.1016/j.jamda.2016.12.086 [DOI] [PubMed] [Google Scholar]
- Chang, J. , Wang, Y. , Shao, L. , Laberge, R.‐M. , Demaria, M. , Campisi, J. , … Zhou, D. (2015). Clearance of senescent cells by ABT263 rejuvenates aged hematopoietic stem cells in mice. Nature Medicine, 22(1), 78–83. 10.1038/nm.4010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen, B. H. , Marioni, R. E. , Colicino, E. , Peters, M. J. , Ward‐Caviness, C. K. , Tsai, P. C. , … Horvath, S. (2016). DNA methylation‐based measures of biological age: Meta‐analysis predicting time to death. Aging (Albany NY), 8(9), 1844–1865. 10.18632/aging.101020 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen, M. Z. , Moily, N. S. , Bridgford, J. L. , Wood, R. J. , Radwan, M. , Smith, T. A. , … Hatters, D. M. (2017). A thiol probe for measuring unfolded protein load and proteostasis in cells. Nature Communications, 8(1), 474 10.1038/s41467-017-00203-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chi, M.‐S. , Lee, C.‐Y. , Huang, S.‐C. , Yang, K.‐L. , Ko, H.‐L. , Chen, Y.‐K. , … Chi, K.‐H. (2015). Double autophagy modulators reduce 2‐deoxyglucose uptake in sarcoma patients. Oncotarget, 6(30), 29808–29817. 10.18632/oncotarget.5060 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Childs, B. G. , Durik, M. , Baker, D. J. , & van Deursen, J. M. (2015). Cellular senescence in aging and age‐related disease: From mechanisms to therapy. Nature Medicine, 21(12), 1424–1435. 10.1038/nm.4000 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Childs, B. G. , Gluscevic, M. , Baker, D. J. , Laberge, R.‐M. , Marquess, D. , Dananberg, J. , & van Deursen, J. M. (2017). Senescent cells: An emerging target for diseases of ageing. Nature Reviews Drug Discovery, 16(10), 718–735. 10.1038/nrd.2017.116 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Childs, B. G. , Li, H. , & van Deursen, J. M. (2018). Senescent cells: A therapeutic target for cardiovascular disease. Journal of Clinical Investigation, 128(4), 1217–1228. 10.1172/jci95146 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Choi, J.‐H. , Kim, S.‐Y. , Kim, S.‐K. , Kemp, M. G. , & Sancar, A. (2015). An integrated approach for analysis of the DNA damage response in mammalian cells. Journal of Biological Chemistry, 290(48), 28812–28821. 10.1074/jbc.m115.690354 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Choi, S. , Reiter, D. A. , Shardell, M. , Simonsick, E. M. , Studenski, S. , Spencer, R. G. , … Ferrucci, L. (2016). 31P Magnetic Resonance Spectroscopy assessment of muscle bioenergetics as a predictor of gait speed in the Baltimore longitudinal study of aging. The Journals of Gerontology Series A: Biological Sciences and Medical Sciences, 71(12), 1638–1645. 10.1093/gerona/glw059 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coen, P. M. , Jubrias, S. A. , Distefano, G. , Amati, F. , Mackey, D. C. , Glynn, N. W. , … Goodpaster, B. H. (2012). Skeletal muscle mitochondrial energetics are associated with maximal aerobic capacity and walking speed in older adults. The Journals of Gerontology: Series A, 68(4), 447–455. 10.1093/gerona/gls196 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coggan, A. R. (1995). Muscle biopsy as a tool in the study of aging. Journals of Gerontology. Series A, Biological Sciences and Medical Sciences, 50, 30–34. [DOI] [PubMed] [Google Scholar]
- Cohen, A. , Morissette‐Thomas, V. , Ferrucci, L. , & Fried, L. (2016). Deep biomarkers of aging are population‐dependent. Aging, 8(9), 2253–2255. 10.18632/aging.101034 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Collins, A. R. (2014). Measuring oxidative damage to DNA and its repair with the comet assay. Biochimica Et Biophysica Acta (BBA) ‐ General Subjects, 1840(2), 794–800. 10.1016/j.bbagen.2013.04.022 [DOI] [PubMed] [Google Scholar]
- Colquitt, B. M. , Allen, W. E. , Barnea, G. , & Lomvardas, S. (2013). Alteration of genic 5-hydroxymethylcytosine patterning in olfactory neurons correlates with changes in gene expression and cell identity. Proceedings of the National Academy of Sciences, 110(36), 14682–14687. 10.1073/pnas.1302759110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Consolini, A. E. , Ragone, M. I. , Bonazzola, P. , & Colareda, G. A. (2017). Mitochondrial Bioenergetics During Ischemia and Reperfusion. Adv Exp Med Biol, 982, 141–167. 10.1007/978-3-319-55330-6_8 [DOI] [PubMed] [Google Scholar]
- Coppé, J.‐P. , Desprez, P.‐Y. , Krtolica, A. , & Campisi, J. (2010). The senescence‐associated secretory phenotype: The dark side of tumor suppression. Annual Review of Pathology: Mechanisms of Disease, 5(1), 99–118. 10.1146/annurev-pathol-121808-102144 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cuervo, A. M. , Bergamini, E. , Brunk, U. T. , Dröge, W. , Ffrench, M. , & Terman, A. (2005). Autophagy and aging: The importance of maintaining "Clean" cells. Autophagy, 1(3), 131–140. 10.4161/auto.1.3.2017 [DOI] [PubMed] [Google Scholar]
- Cuervo, A. M. , & Macian, F. (2014). Autophagy and the immune function in aging. Current Opinion in Immunology, 29, 97–104. 10.1016/j.coi.2014.05.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cuervo, A. M. , Wong, E. S. P. , & Martinez‐Vicente, M. (2010). Protein degradation, aggregation, and misfolding. Movement Disorders, 25(S1), S49–S54. 10.1002/mds.22718 [DOI] [PubMed] [Google Scholar]
- de Haan, G. , & Lazare, S. S. (2017). Aging of hematopoietic stem cells. Blood, 131(5), 479–487. 10.1182/blood-2017-06-746412 [DOI] [PubMed] [Google Scholar]
- Degerman, S. , Josefsson, M. , Nordin Adolfsson, A. , Wennstedt, S. , Landfors, M. , Haider, Z. , … Adolfsson, R. (2017). Maintained memory in aging is associated with young epigenetic age. Neurobiology of Aging, 55, 167–171. 10.1016/j.neurobiolaging.2017.02.009 [DOI] [PubMed] [Google Scholar]
- Dexheimer, V. , Mueller, S. , Braatz, F. , & Richter, W. (2011). Reduced reactivation from dormancy but maintained lineage choice of human mesenchymal stem cells with donor age. PLoS ONE, 6(8), e22980 10.1371/journal.pone.0022980 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Diekman, B. O. , Sessions, G. A. , Collins, J. A. , Knecht, A. K. , Strum, S. L. , Mitin, N. K. , … Sharpless, N. E. (2018). Expression of p16INK 4a is a biomarker of chondrocyte aging but does not cause osteoarthritis. Aging Cell, 17(4), e12771 10.1111/acel.12771 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dikalov, S. I. , & Harrison, D. G. (2014). Methods for detection of mitochondrial and cellular reactive oxygen species. Antioxidants & Redox Signaling, 20(2), 372–382. 10.1089/ars.2012.4886 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ding, J. , Sidore, C. , Butler, T. J. , Wing, M. K. , Qian, Y. , Meirelles, O. , … Schlessinger, D. (2015). Assessing mitochondrial DNA variation and copy number in lymphocytes of ~2,000 Sardinians using tailored sequencing analysis tools. PLoS Genetics, 11(7), e1005306 10.1371/journal.pgen.1005306 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eerola, J. , Kananen, L. , Manninen, K. , Hellstrom, O. , Tienari, P. J. , & Hovatta, I. (2010). No evidence for shorter leukocyte telomere length in Parkinson's disease patients. The Journals of Gerontology Series A: Biological Sciences and Medical Sciences, 65A(11), 1181–1184. 10.1093/gerona/glq125 [DOI] [PubMed] [Google Scholar]
- Eichler, F. , Duncan, C. , Musolino, P. L. , Orchard, P. J. , De Oliveira, S. , Thrasher, A. J. , … Williams, D. A. (2017). Hematopoietic stem‐cell gene therapy for cerebral adrenoleukodystrophy. New England Journal of Medicine, 377(17), 1630–1638. 10.1056/nejmoa1700554 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eirin, A. , Saad, A. , Tang, H. , Herrmann, S. M. , Woollard, J. R. , Lerman, A. , … Lerman, L. O. (2016). Urinary mitochondrial DNA copy number identifies chronic renal injury in hypertensive patients. Hypertension, 68(2), 401–410. 10.1161/hypertensionaha.116.07849 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eisenberg, T. , Knauer, H. , Schauer, A. , Buttner, S. , Ruckenstuhl, C. , Carmona‐Gutierrez, D. , … Madeo, F. (2009). Induction of autophagy by spermidine promotes longevity. Nature Cell Biology, 11(11), 1305–1314. 10.1038/ncb1975 [DOI] [PubMed] [Google Scholar]
- El‐Chemaly, S. , Cheung, F. , Kotliarov, Y. , O’Brien, K. J. , Gahl, W. A. , Chen, J. , … Gochuico, B. R. (2018). The immunome in two inherited forms of pulmonary fibrosis. Frontiers in Immunology, 9, 76 10.3389/fimmu.2018.00076 [DOI] [PMC free article] [PubMed] [Google Scholar]
- El‐Chemaly, S. , Taveira‐Dasilva, A. , Goldberg, H. J. , Peters, E. , Haughey, M. , Bienfang, D. , … Henske, E. P. (2017). Sirolimus and autophagy inhibition in lymphangioleiomyomatosis. Chest, 151(6), 1302–1310. 10.1016/j.chest.2017.01.033 [DOI] [PMC free article] [PubMed] [Google Scholar]
- El‐Zein, R. A. , Monroy, C. M. , Cortes, A. , Spitz, M. R. , Greisinger, A. , & Etzel, C. J. (2010). Rapid method for determination of DNA repair capacity in human peripheral blood lymphocytes amongst smokers. BMC Cancer, 10(1), 439 10.1186/1471-2407-10-439 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Epel, E. S. , Blackburn, E. H. , Lin, J. , Dhabhar, F. S. , Adler, N. E. , Morrow, J. D. , & Cawthon, R. M. (2004). Accelerated telomere shortening in response to life stress. Proceedings of the National Academy of Sciences, 101(49), 17312–17315. 10.1073/pnas.0407162101 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Evangelou, K. , Lougiakis, N. , Rizou, S. V. , Kotsinas, A. , Kletsas, D. , Muñoz‐Espín, D. , … Gorgoulis, V. G. (2016). Robust, universal biomarker assay to detect senescent cells in biological specimens. Aging Cell, 16(1), 192–197. 10.1111/acel.12545 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fabbri, E. , An, Y. , Zoli, M. , Simonsick, E. M. , Guralnik, J. M. , Bandinelli, S. , … Ferrucci, L. (2014). Aging and the burden of multimorbidity: Associations with inflammatory and anabolic hormonal biomarkers. The Journals of Gerontology: Series A, 70(1), 63–70. 10.1093/gerona/glu127 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fabbri, E. , Chia, C. W. , Spencer, R. G. , Fishbein, K. W. , Reiter, D. A. , Cameron, D. , … Ferrucci, L. (2016). Insulin resistance is associated with reduced mitochondrial oxidative capacity measured by 31P‐magnetic resonance spectroscopy in participants without diabetes from the Baltimore longitudinal study of aging. Diabetes, 66(1), 170–176. 10.2337/db16-0754 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fabbri, E. , Zoli, M. , Gonzalez‐Freire, M. , Salive, M. E. , Studenski, S. A. , & Ferrucci, L. (2015). Aging and multimorbidity: New tasks, priorities, and frontiers for integrated gerontological and clinical research. Journal of the American Medical Directors Association, 16(8), 640–647. 10.1016/j.jamda.2015.03.013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fan, M. , Chen, W. , Liu, W. , Du, G.‐Q. , Jiang, S.‐L. , Tian, W.‐C. , … Tian, H. (2010). The effect of age on the efficacy of human mesenchymal stem cell transplantation after a myocardial infarction. Rejuvenation Research, 13(4), 429–438. 10.1089/rej.2009.0986 [DOI] [PubMed] [Google Scholar]
- Fang, L. , Neutzner, A. , Turtschi, S. , Flammer, J. , & Mozaffarieh, M. (2015). Comet Assay as an indirect measure of systemic oxidative stress. Journal of Visualized Experiments, 99, e52763 10.3791/52763 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ferrucci, L. , Corsi, A. , Lauretani, F. , Bandinelli, S. , Bartali, B. , Taub, D. D. , … Longo, D. L. (2005). The origins of age‐related proinflammatory state. Blood, 105(6), 2294–2299. 10.1182/blood-2004-07-2599 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ferrucci, L. , & Fabbri, E. (2018). Inflammageing: Chronic inflammation in ageing, cardiovascular disease, and frailty. Nature Reviews Cardiology, 15(9), 505–522. 10.1038/s41569-018-0064-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ferrucci, L. , Harris, T. B. , Guralnik, J. M. , Tracy, R. P. , Corti, M.‐C. , Cohen, H. J. , … Havlik, R. J. (1999). Serum IL‐6 level and the development of disability in older persons. Journal of the American Geriatrics Society, 47(6), 639–646. 10.1111/j.1532-5415.1999.tb01583.x [DOI] [PubMed] [Google Scholar]
- Ferrucci, L. , Levine, M. E. , Kuo, P. L. , & Simonsick, E. M. (2018). Time and the metrics of aging. Circulation Research, 123(7), 740–744. 10.1161/CIRCRESAHA.118.312816 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ferrucci, L. , Penninx, B. W. J. H. , Volpato, S. , Harris, T. B. , Bandeen‐Roche, K. , Balfour, J. , … Md, J. M. G. (2002). Change in muscle strength explains accelerated decline of physical function in older women with high interleukin‐6 serum levels. Journal of the American Geriatrics Society, 50(12), 1947–1954. 10.1046/j.1532-5415.2002.50605.x [DOI] [PubMed] [Google Scholar]
- Franceschi, C. , & Campisi, J. (2014). Chronic inflammation (Inflammaging) and its potential contribution to age‐associated diseases. The Journals of Gerontology Series A: Biological Sciences and Medical Sciences, 69(Suppl. 1), S4–S9. 10.1093/gerona/glu057 [DOI] [PubMed] [Google Scholar]
- Franco, I. , Johansson, A. , Olsson, K. , Vrtačnik, P. , Lundin, P. , Helgadottir, H. T. , … Eriksson, M. (2018). Somatic mutagenesis in satellite cells associates with human skeletal muscle aging. Nature Communications, 9(1), 800 10.1038/s41467-018-03244-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Friedman, E. M. , Christ, S. L. , & Mroczek, D. K. (2015). Inflammation partially mediates the association of multimorbidity and functional limitations in a national sample of middle‐aged and older adults. Journal of Aging and Health, 27(5), 843–863. 10.1177/0898264315569453 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fulop, T. , Larbi, A. , Dupuis, G. , Le Page, A. , Frost, E. H. , Cohen, A. A. , … Franceschi, C. (2018). Immunosenescence and inflamm‐aging as two sides of the same coin: Friends or foes? Frontiers in Immunology, 8, 1960 10.3389/fimmu.2017.01960 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gale, C. R. , Marioni, R. E. , Čukić, I. , Chastin, S. F. , Dall, P. M. , Dontje, M. L. , … Deary, I. J. (2018). The epigenetic clock and objectively measured sedentary and walking behavior in older adults: The Lothian Birth Cohort 1936. Clinical Epigenetics, 10(1), 4 10.1186/s13148-017-0438-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- García‐Calzón, S. , Gea, A. , Razquin, C. , Corella, D. , Lamuela‐Raventós, R. M. , Martínez, J. A. , … Marti, A. (2013). Longitudinal association of telomere length and obesity indices in an intervention study with a Mediterranean diet: The PREDIMED‐NAVARRA trial. International Journal of Obesity, 38(2), 177–182. 10.1038/ijo.2013.68 [DOI] [PubMed] [Google Scholar]
- García‐Prat, L. , Martínez‐Vicente, M. , Perdiguero, E. , Ortet, L. , Rodríguez‐Ubreva, J. , Rebollo, E. , … Muñoz‐Cánoves, P. (2016). Autophagy maintains stemness by preventing senescence. Nature, 529(7584), 37–42. 10.1038/nature16187 [DOI] [PubMed] [Google Scholar]
- Gensous, N. , Bacalini, M. G. , Pirazzini, C. , Marasco, E. , Giuliani, C. , Ravaioli, F. , … Garagnani, P. (2017). The epigenetic landscape of age‐related diseases: The geroscience perspective. Biogerontology, 18(4), 549–559. 10.1007/s10522-017-9695-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- GBD 2017 Disease and Injury Incidence and Prevalence Collaborators . (2016). Global, regional, and national incidence, prevalence, and years lived with disability for 310 diseases and injuries, 1990–2015: A systematic analysis for the Global Burden of Disease Study 2015. The Lancet, 388(10053), 1545–1602. 10.1016/s0140-6736(16)31678-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Godderis, L. , Schouteden, C. , Tabish, A. , Poels, K. , Hoet, P. , Baccarelli, A. A. , & Van Landuyt, K. (2015). Global methylation and hydroxymethylation in DNA from blood and saliva in healthy volunteers. BioMed Research International, 2015, 845041 10.1155/2015/845041 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goglin, S. E. , Farzaneh‐Far, R. , Epel, E. S. , Lin, J. , Blackburn, E. H. , & Whooley, M. A. (2016). Correction: Change in leukocyte telomere length predicts mortality in patients with stable coronary heart disease from the heart and soul study. PLoS ONE, 11(12), e0168868 10.1371/journal.pone.0168868 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Golpanian, S. , DiFede, D. L. , Khan, A. , Schulman, I. H. , Landin, A. M. , Tompkins, B. A. , … Hare, J. M. (2017). Allogeneic human mesenchymal stem cell infusions for aging frailty. The Journals of Gerontology: Series A, 72(11), 1505–1512. 10.1093/gerona/glx056 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Golpanian, S. , DiFede, D. L. , Pujol, M. V. , Lowery, M. H. , Levis‐Dusseau, S. , Goldstein, B. J. , … Hare, J. M. (2016). Rationale and design of the allogeneiC human mesenchymal stem cells (hMSC) in patients with aging fRAilTy via intravenoUS delivery (CRATUS) study: A phase I/II, randomized, blinded and placebo controlled trial to evaluate the safety and potential efficacy of allogeneic human mesenchymal stem cell infusion in patients with aging frailty. Oncotarget, 7(11), 11899–11912. 10.18632/oncotarget.7727 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gonzales‐Ebsen, A. C. , Gregersen, N. , & Olsen, R. K. (2017). Linking telomere loss and mitochondrial dysfunction in chronic disease. Frontiers in Bioscience (Landmark Ed), 22, 117–127. [DOI] [PubMed] [Google Scholar]
- Gonzalez‐Freire, M. , de Cabo, R. , Bernier, M. , Sollott, S. J. , Fabbri, E. , Navas, P. , & Ferrucci, L. (2015). Reconsidering the role of mitochondria in aging. Journals of Gerontology. Series A, Biological Sciences and Medical Sciences, 70, 1334–1342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gonzalez‐Freire, M. , Scalzo, P. , D'Agostino, J. , Moore, Z. A. , Diaz‐Ruiz, A. , Fabbri, E. , … Ferrucci, L. (2018). Skeletal muscle ex vivo mitochondrial respiration parallels decline in vivo oxidative capacity, cardiorespiratory fitness, and muscle strength: The Baltimore Longitudinal Study of Aging. Aging Cell, 17(2), e12725 10.1111/acel.12725 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Greider, C. W. (1998). Telomeres and senescence: The history, the experiment, the future. Current Biology, 8(5), R178–R181. 10.1016/s0960-9822(98)70105-8 [DOI] [PubMed] [Google Scholar]
- Greider, C. W. (2010). Telomerase discovery: The excitement of putting together pieces of the puzzle (nobel lecture). Angewandte Chemie International Edition, 49(41), 7422–7439. 10.1002/anie.201002408 [DOI] [PubMed] [Google Scholar]
- Guiding Principles for the Care of Older Adults with Multimorbidity: An Approach for Clinicians . (2012). Guiding principles for the care of older adults with multimorbidity: An approach for clinicians: American Geriatrics Society Expert Panel on the Care of Older Adults with Multimorbidity. Journal of the American Geriatrics Society, 60(10), E1–E25. 10.1111/j.1532-5415.2012.04188.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gupta, V. K. , Scheunemann, L. , Eisenberg, T. , Mertel, S. , Bhukel, A. , Koemans, T. S. , … Sigrist, S. J. (2013). Restoring polyamines protects from age‐induced memory impairment in an autophagy‐dependent manner. Nature Neuroscience, 16(10), 1453–1460. 10.1038/nn.3512 [DOI] [PubMed] [Google Scholar]
- Haferkamp, S. , Scurr, L. L. , Becker, T. M. , Frausto, M. , Kefford, R. F. , & Rizos, H. (2009). Oncogene‐induced senescence does not require the p16INK4a or p14ARF melanoma tumor suppressors. Journal of Investigative Dermatology, 129(8), 1983–1991. 10.1038/jid.2009.5 [DOI] [PubMed] [Google Scholar]
- Hall, B. M. , Balan, V. , Gleiberman, A. S. , Strom, E. , Krasnov, P. , Virtuoso, L. P. , … Gudkov, A. V. (2017). p16(Ink4a) and senescence‐associated β‐galactosidase can be induced in macrophages as part of a reversible response to physiological stimuli. Aging, 9(8), 1867–1884. 10.18632/aging.101268 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hamann, I. , & Hartwig, A. (2014). Quantification of DNA repair capacity towards oxidatively damaged DNA in subcellular and cellular systems by a nonradioactive cleavage assay. Methods Mol Biol, 1208, 73–84. 10.1007/978-1-4939-1441-8_6 [DOI] [PubMed] [Google Scholar]
- Hammadah, M. , Al Mheid, I. , Wilmot, K. , Ramadan, R. , Abdelhadi, N. , Alkhoder, A. , … Vaccarino, V. (2017). Telomere shortening, regenerative capacity, and cardiovascular outcomes. Circulation Research, 120(7), 1130–1138. 10.1161/circresaha.116.309421 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hannum, G. , Guinney, J. , Zhao, L. , Zhang, L. , Hughes, G. , Sadda, S. , … Zhang, K. (2013). Genome‐wide methylation profiles reveal quantitative views of human aging rates. Molecular Cell, 49(2), 359–367. 10.1016/j.molcel.2012.10.016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hare, J. M. , Fishman, J. E. , Gerstenblith, G. , DiFede Velazquez, D. L. , Zambrano, J. P. , Suncion, V. Y. , … Heldman, A. W. (2012). Comparison of allogeneic vs autologous bone marrow–derived mesenchymal stem cells delivered by transendocardial injection in patients with ischemic cardiomyopathy. JAMA, 308(22), 2369 10.1001/jama.2012.25321 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harman, D. (1956). Aging: A theory based on free radical and radiation chemistry. Journal of Gerontology, 11(3), 298–300. 10.1093/geronj/11.3.298 [DOI] [PubMed] [Google Scholar]
- Harman, D. (1972). The biologic clock: The mitochondria? Journal of the American Geriatrics Society, 20(4), 145–147. 10.1111/j.1532-5415.1972.tb00787.x [DOI] [PubMed] [Google Scholar]
- Harman, D. (2003). The free radical theory of aging. Antioxidants & Redox Signaling, 5(5), 557–561. 10.1089/152308603770310202 [DOI] [PubMed] [Google Scholar]
- Hayashi, G. , & Cortopassi, G. (2015). Oxidative stress in inherited mitochondrial diseases. Free Radical Biology and Medicine, 88, 10–17. 10.1016/j.freeradbiomed.2015.05.039 [DOI] [PMC free article] [PubMed] [Google Scholar]
- He, Z. , Bian, J. , Carretta, H. J. , Lee, J. , Hogan, W. R. , Shenkman, E. , & Charness, N. (2018). Prevalence of multiple chronic conditions among older adults in Florida and the United States: Comparative analysis of the OneFlorida Data trust and national inpatient sample. Journal of Medical Internet Research, 20(4), e137 10.2196/jmir.8961 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heidinger, B. J. , Blount, J. D. , Boner, W. , Griffiths, K. , Metcalfe, N. B. , & Monaghan, P. (2012). Telomere length in early life predicts life span. Obstetrical & Gynecological Survey, 67(5), 283–284. 10.1097/ogx.0b013e3182546dd0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herbig, U. , Jobling, W. A. , Chen, B. P. C. , Chen, D. J. , & Sedivy, J. M. (2004). Telomere shortening triggers senescence of human cells through a pathway involving ATM, p53, and p21CIP1, but not p16INK4a. Molecular Cell, 14(4), 501–513. 10.1016/s1097-2765(04)00256-4 [DOI] [PubMed] [Google Scholar]
- Herranz, N. , & Gil, J. (2018). Mechanisms and functions of cellular senescence. Journal of Clinical Investigation, 128(4), 1238–1246. 10.1172/jci95148 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hewitt, G. , Jurk, D. , Marques, F. D. M. , Correia‐Melo, C. , Hardy, T. , Gackowska, A. , … Passos, J. F. (2012). Telomeres are favoured targets of a persistent DNA damage response in ageing and stress‐induced senescence. Nature Communications, 3(1), 708 10.1038/ncomms1708 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hilmer, S. N. , & Le Couteur, D. G. (2016). Standardized, multidisciplinary approaches for the study of aging biology and for translation of aging interventions. The Journals of Gerontology Series A: Biological Sciences and Medical Sciences, 71(4), 425–426. 10.1093/gerona/glv310 [DOI] [PubMed] [Google Scholar]
- Hinken, A. C. , & Billin, A. N. (2018). Isolation of Skeletal Muscle Stem Cells for Phenotypic Screens for Modulators of Proliferation. Methods Mol Biol, 1787, 77–86. https://doi.org/0.1007/978-1-4939-7847-2_6 [DOI] [PubMed] [Google Scholar]
- Hoffman, J. M. , Lyu, Y. , Pletcher, S. D. , & Promislow, D. E. L. (2017). Proteomics and metabolomics in ageing research: From biomarkers to systems biology. Essays in Biochemistry, 61(3), 379–388. 10.1042/ebc20160083 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holloway, G. P. , Holwerda, A. M. , Miotto, P. M. , Dirks, M. L. , Verdijk, L. B. , & van Loon, L. J. C. (2018). Age‐associated impairments in mitochondrial ADP sensitivity contribute to redox stress in senescent human skeletal muscle. Cell Reports, 22(11), 2837–2848. 10.1016/j.celrep.2018.02.069 [DOI] [PubMed] [Google Scholar]
- Holton, N. W. , Ebenstein, Y. , & Gassman, N. R. (2018). Broad spectrum detection of DNA damage by Repair Assisted Damage Detection (RADD). DNA Repair, 66–67, 42–49. 10.1016/j.dnarep.2018.04.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Horvath, S. (2013). DNA methylation age of human tissues and cell types. Genome Biology, 14(10), R115 10.1186/gb-2013-14-10-r115 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hughes, M. C. , Ramos, S. V. , Turnbull, P. C. , Nejatbakhsh, A. , Baechler, B. L. , Tahmasebi, H. , … Perry, C. G. R. (2015). Mitochondrial bioenergetics and fiber type assessments in microbiopsy vs. bergstrom percutaneous sampling of human skeletal muscle. Frontiers in Physiology, 6, 360 10.3389/fphys.2015.00360 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jani, B. D. , Nicholl, B. I. , McQueenie, R. , Connelly, D. T. , Hanlon, P. , Gallacher, K. I. , … Mair, F. S. (2017). Multimorbidity and co‐morbidity in atrial fibrillation and effects on survival: Findings from UK Biobank cohort. EP Europace, 20(FI_3), f329–f336. 10.1093/europace/eux322 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jim, H. S. L. , Sutton, S. K. , Small, B. J. , Jacobsen, P. B. , Wood, W. A. , Knight, J. M. , … Lee, S. J. (2016). Trajectories of quality of life after hematopoietic cell transplantation: Secondary analysis of blood and marrow transplant clinical trials network 0902 data. Biology of Blood and Marrow Transplantation, 22(11), 2077–2083. 10.1016/j.bbmt.2016.08.012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jiménez‐Chillarón, J. C. , Díaz, R. , Martínez, D. , Pentinat, T. , Ramón‐Krauel, M. , Ribó, S. , & Plösch, T. (2012). The role of nutrition on epigenetic modifications and their implications on health. Biochimie, 94(11), 2242–2263. 10.1016/j.biochi.2012.06.012 [DOI] [PubMed] [Google Scholar]
- Jimenez‐Chillaron, J. C. , Hernandez‐Valencia, M. , Lightner, A. , Faucette, R. R. , Reamer, C. , Przybyla, R. , … Patti, M. E. (2006). Reductions in caloric intake and early postnatal growth prevent glucose intolerance and obesity associated with low birthweight. Diabetologia, 49(8), 1974–1984. 10.1007/s00125-006-0311-7 [DOI] [PubMed] [Google Scholar]
- Jodczyk, S. , Fergusson, D. M. , Horwood, L. J. , Pearson, J. F. , & Kennedy, M. A. (2014). No association between mean telomere length and life stress observed in a 30 year birth cohort. PLoS ONE, 9(5), e97102 10.1371/journal.pone.0097102 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Justice, J. N. , Gregory, H. , Tchkonia, T. , LeBrasseur, N. K. , Kirkland, J. L. , Kritchevsky, S. B. , & Nicklas, B. J. (2018). Cellular senescence biomarker p16INK4a+ cell burden in thigh adipose is associated with poor physical function in older women. Journals of Gerontology. Series A, Biological Sciences and Medical Sciences, 73(7), 939–945. 10.1093/gerona/glx134 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jylhävä, J. , Pedersen, N. L. , & Hägg, S. (2017). Biological age predictors. Ebiomedicine, 21, 29–36. 10.1016/j.ebiom.2017.03.046 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jylhävä, J. , Raitanen, J. , Marttila, S. , Hervonen, A. , Jylhä, M. , & Hurme, M. (2014). Identification of a prognostic signature for old‐age mortality by integrating genome‐wide transcriptomic data with the conventional predictors: The Vitality 90+ Study. BMC Medical Genomics, 7(1), 10.1186/1755-8794-7-54 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kadota, T. , Fujita, Y. , Yoshioka, Y. , Araya, J. , Kuwano, K. , & Ochiya, T. (2018). Emerging role of extracellular vesicles as a senescence‐associated secretory phenotype: Insights into the pathophysiology of lung diseases. Molecular Aspects of Medicine, 60, 92–103. 10.1016/j.mam.2017.11.005 [DOI] [PubMed] [Google Scholar]
- Kananen, L. , Marttila, S. , Nevalainen, T. , Kummola, L. , Junttila, I. , Mononen, N. , … Jylhävä, J. (2016). The trajectory of the blood DNA methylome ageing rate is largely set before adulthood: Evidence from two longitudinal studies. AGE, 38(3), 65 10.1007/s11357-016-9927-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kang, C. , Xu, Q. , Martin, T. D. , Li, M. Z. , Demaria, M. , Aron, L. , … Elledge, S. J. (2015). The DNA damage response induces inflammation and senescence by inhibiting autophagy of GATA4. Science, 349(6255), aaa5612–aaa5612. 10.1126/science.aaa5612 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kang, E. , Wang, X. , Tippner‐Hedges, R. , Ma, H. , Folmes, C. D. L. , Gutierrez, N. M. , … Mitalipov, S. (2016). Age‐related accumulation of somatic mitochondrial DNA mutations in adult‐derived human iPSCs. Cell Stem Cell, 18(5), 625–636. 10.1016/j.stem.2016.02.005 [DOI] [PubMed] [Google Scholar]
- Kaushik, S. , & Cuervo, A. M. (2018). The coming of age of chaperone‐mediated autophagy. Nature Reviews Molecular Cell Biology, 19(6), 365–381. 10.1038/s41580-018-0001-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kent, J. A. , & Fitzgerald, L. F. (2016). In vivo mitochondrial function in aging skeletal muscle: Capacity, flux, and patterns of use. Journal of Applied Physiology, 121(4), 996–1003. 10.1152/japplphysiol.00583.2016 [DOI] [PubMed] [Google Scholar]
- Kim, S. , & Jazwinski, S. M. (2015). Quantitative measures of healthy aging and biological age. Healthy Aging Research, 4, 10.12715/har.2015.4.26 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kirkland, J. L. , & Tchkonia, T. (2017). Cellular senescence: A translational perspective. Ebiomedicine, 21, 21–28. 10.1016/j.ebiom.2017.04.013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kirkland, J. L. , Tchkonia, T. , Zhu, Y. , Niedernhofer, L. J. , & Robbins, P. D. (2017). The clinical potential of senolytic drugs. Journal of the American Geriatrics Society, 65(10), 2297–2301. 10.1111/jgs.14969 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klionsky, D. J. (2014). Coming soon to a journal near you—the updated guidelines for the use and interpretation of assays for monitoring autophagy. Autophagy, 10(10), 1691–1691. 10.4161/auto.36187 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klionsky, D. J. , Cuervo, A. M. , & Seglen, P. O. (2007). Methods for monitoring autophagy from yeast to human. Autophagy, 3(3), 181–206. 10.4161/auto.3678 [DOI] [PubMed] [Google Scholar]
- Knight, A. K. , Craig, J. M. , Theda, C. , Baekvad‐Hansen, M. , Bybjerg‐Grauholm, J. , Hansen, C. S. , … Smith, A. K. (2016). An epigenetic clock for gestational age at birth based on blood methylation data. Genome Biology, 17(1), 206 10.1186/s13059-016-1068-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koppelstaetter, C. , Schratzberger, G. , Perco, P. , Hofer, J. , Mark, W. , Ollinger, R. , … Mayer, G. (2008). Markers of cellular senescence in zero hour biopsies predict outcome in renal transplantation. Aging Cell, 7(4), 491–497. 10.1111/j.1474-9726.2008.00398.x [DOI] [PubMed] [Google Scholar]
- Laberge, R.‐M. , Zhou, L. , Sarantos, M. R. , Rodier, F. , Freund, A. , de Keizer, P. L. J. , … Campisi, J. (2012). Glucocorticoids suppress selected components of the senescence‐associated secretory phenotype. Aging Cell, 11(4), 569–578. 10.1111/j.1474-9726.2012.00818.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lanza, I. R. , & Nair, K. S. (2010). Mitochondrial metabolic function assessed in vivo and in vitro. Current Opinion in Clinical Nutrition and Metabolic Care, 13(5), 511–517. 10.1097/mco.0b013e32833cc93d [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lara, J. , Cooper, R. , Nissan, J. , Ginty, A. T. , Khaw, K.‐T. , Deary, I. J. , … Mathers, J. C. (2015). A proposed panel of biomarkers of healthy ageing. BMC Medicine, 13(1), 222 10.1186/s12916-015-0470-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Larsen, S. , Nielsen, J. , Hansen, C. N. , Nielsen, L. B. , Wibrand, F. , Stride, N. , … Hey‐Mogensen, M. (2012). Biomarkers of mitochondrial content in skeletal muscle of healthy young human subjects. The Journal of Physiology, 590(14), 3349–3360. 10.1113/jphysiol.2012.230185 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Larsson, N.‐G. (2010). Somatic mitochondrial DNA mutations in mammalian aging. Annual Review of Biochemistry, 79(1), 683–706. 10.1146/annurev-biochem-060408-093701 [DOI] [PubMed] [Google Scholar]
- Latimer, J. J. , & Kelly, C. M. (2014). Unscheduled DNA synthesis: the clinical and functional assay for global genomic DNA nucleotide excision repair. Methods Mol Biol, 1105, 511–532. 10.1007/978-1-62703-739-6_36 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lavker, R. M. , & Sun, T. T. (2000). Epidermal stem cells: Properties, markers, and location. Proceedings of the National Academy of Sciences, 97(25), 13473–13475. 10.1073/pnas.250380097 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee, I. H. , Cao, L. , Mostoslavsky, R. , Lombard, D. B. , Liu, J. , Bruns, N. E. , … Finkel, T. (2008). A role for the NAD‐dependent deacetylase Sirt1 in the regulation of autophagy. Proceedings of the National Academy of Sciences of the United States of America, 105(9), 3374–3379. 10.1073/pnas.0712145105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lev Maor, G. , Yearim, A. , & Ast, G. (2015). The alternative role of DNA methylation in splicing regulation. Trends in Genetics, 31(5), 274–280. 10.1016/j.tig.2015.03.002 [DOI] [PubMed] [Google Scholar]
- Levine, M. E. , Lu, A. T. , Quach, A. , Chen, B. H. , Assimes, T. L. , Bandinelli, S. , … Horvath, S. (2018). An epigenetic biomarker of aging for lifespan and healthspan. Aging, 10(4), 573–591. 10.18632/aging.101414 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li, C. , Gao, W. , Gao, Y. , Yu, C. , Lv, J. , Lv, R. , … Li, L. (2018). Age prediction of children and adolescents aged 6–17 years: An epigenome‐wide analysis of DNA methylation. Aging, 10(5), 1015–1026. 10.18632/aging.101445 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li, M. , Chen, M. , Han, W. , & Fu, X. (2010). How far are induced pluripotent stem cells from the clinic? Ageing Research Reviews, 9(3), 257–264. 10.1016/j.arr.2010.03.001 [DOI] [PubMed] [Google Scholar]
- Li, Y. , Huang, J. , Pang, S. , Wang, H. , Zhang, A. , Hawley, R. G. , & Yan, B. (2017). Novel and functional ATG12 gene variants in sporadic Parkinson's disease. Neuroscience Letters, 643, 22–26. 10.1016/j.neulet.2017.02.028 [DOI] [PubMed] [Google Scholar]
- Ligthart, S. , Marzi, C. , Aslibekyan, S. , Mendelson, M. M. , Conneely, K. N. , Tanaka, T. , … Dehghan, A. (2016). DNA methylation signatures of chronic low‐grade inflammation are associated with complex diseases. Genome Biology, 17(1), 255 10.1186/s13059-016-1119-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin, J. , Cheon, J. , Brown, R. , Coccia, M. , Puterman, E. , Aschbacher, K. , … Blackburn, E. H. (2016). Systematic and cell type‐specific telomere length changes in subsets of lymphocytes. Journal of Immunology Research, 2016, 1–9. 10.1155/2016/5371050 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin, Y. , Damjanovic, A. , Metter, E. J. , Nguyen, H. , Truong, T. , Najarro, K. , … Weng, N.‐P. (2015). Age‐associated telomere attrition of lymphocytes in vivo is co‐ordinated with changes in telomerase activity, composition of lymphocyte subsets and health conditions. Clinical Science, 128(6), 367–377. 10.1042/cs20140481 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu, L. , Cheung, T. H. , Charville, G. W. , & Rando, T. A. (2015). Isolation of skeletal muscle stem cells by fluorescence‐activated cell sorting. Nature Protocols, 10(10), 1612–1624. 10.1038/nprot.2015.110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu, Y. , Sanoff, H. K. , Cho, H. , Burd, C. E. , Torrice, C. , Ibrahim, J. G. , … Sharpless, N. E. (2009). Expression of p16INK4a in peripheral blood T‐cells is a biomarker of human aging. Aging Cell, 8(4), 439–448. 10.1111/j.1474-9726.2009.00489.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lodato, M. A. , Rodin, R. E. , Bohrson, C. L. , Coulter, M. E. , Barton, A. R. , Kwon, M. , & Walsh, C. A. (2017). Aging and neurodegeneration are associated with increased mutations in single human neurons. Science, 359(6375), 555–559. 10.1126/science.aao4426 [DOI] [PMC free article] [PubMed] [Google Scholar]
- López‐Otín, C. , Blasco, M. A. , Partridge, L. , Serrano, M. , & Kroemer, G. (2013). The Hallmarks of aging. Cell, 153(6), 1194–1217. 10.1016/j.cell.2013.05.039 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lorenzi, M. , Bonassi, S. , Lorenzi, T. , Giovannini, S. , Bernabei, R. , & Onder, G. (2018). A review of telomere length in sarcopenia and frailty. Biogerontology, 19(3–4), 209–221. 10.1007/s10522-018-9749-5 [DOI] [PubMed] [Google Scholar]
- Lorite Mingot, D. , Gesteiro, E. , Bastida, S. , & Sánchez‐Muniz, F. J. (2017). Epigenetic effects of the pregnancy Mediterranean diet adherence on the offspring metabolic syndrome markers. Journal of Physiology and Biochemistry, 73(4), 495–510. 10.1007/s13105-017-0592-y [DOI] [PubMed] [Google Scholar]
- Lozano‐Torres, B. , Galiana, I. , Rovira, M. , Garrido, E. , Chaib, S. , Bernardos, A. , … Sancenón, F. (2017). An OFF–ON two‐photon fluorescent probe for tracking cell senescence in vivo. Journal of the American Chemical Society, 139(26), 8808–8811. 10.1021/jacs.7b04985 [DOI] [PubMed] [Google Scholar]
- Lu, A. T. , Quach, A. , Wilson, J. G. , Reiner, A. P. , Aviv, A. , Raj, K. , … Horvath, S. (2019). DNA methylation GrimAge strongly predicts lifespan and healthspan. Aging, 11(2), 303–327. 10.18632/aging.101684 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lustig, A. , Liu, H. B. , Metter, E. J. , An, Y. , Swaby, M. A. , Elango, P. , … Weng, N.‐P. (2017). Telomere shortening, inflammatory cytokines, and anti‐cytomegalovirus antibody follow distinct age‐associated trajectories in humans. Frontiers in Immunology, 8, 1027 10.3389/fimmu.2017.01027 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Madeo, F. , Eisenberg, T. , Pietrocola, F. , & Kroemer, G. (2018). Spermidine in health and disease. Science, 359(6374), 359–410. 10.1126/science.aan2788 [DOI] [PubMed] [Google Scholar]
- Maierhofer, A. , Flunkert, J. , Oshima, J. , Martin, G. M. , Haaf, T. , & Horvath, S. (2017). Accelerated epigenetic aging in Werner syndrome. Aging, 9(4), 1143–1152. 10.18632/aging.101217 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mannick, J. B. , Del Giudice, G. , Lattanzi, M. , Valiante, N. M. , Praestgaard, J. , Huang, B. , … Klickstein, L. B. (2014). mTOR inhibition improves immune function in the elderly. Science Translational Medicine, 6(268), 268ra179–268ra179. 10.1126/scitranslmed.3009892 [DOI] [PubMed] [Google Scholar]
- Marioni, R. E. , Shah, S. , McRae, A. F. , Chen, B. H. , Colicino, E. , Harris, S. E. , … Deary, I. J. (2015). DNA methylation age of blood predicts all‐cause mortality in later life. Genome Biology, 16, 25 10.1186/s13059-015-0584-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marioni, R. E. , Shah, S. , McRae, A. F. , Ritchie, S. J. , Muniz‐Terrera, G. , Harris, S. E. , … Deary, I. J. (2015). The epigenetic clock is correlated with physical and cognitive fitness in the Lothian Birth Cohort 1936. International Journal of Epidemiology, 44(4), 1388–1396. 10.1093/ije/dyu277 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marioni, R. E. , Suderman, M. , Chen, B. H. , Horvath, S. , Bandinelli, S. , Morris, T. , … Hagg, S. (2019). Tracking the epigenetic clock across the human life course: A meta‐analysis of longitudinal cohort data. Journals of Gerontology. Series A, Biological Sciences and Medical Sciences, 74(1), 57–61. 10.1093/gerona/gly060 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marrocco, I. , Altieri, F. , & Peluso, I. (2017). Measurement and clinical significance of biomarkers of oxidative stress in humans. Oxidative Medicine and Cellular Longevity, 2017, 1–32. 10.1155/2017/6501046 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marttila, S. , Kananen, L. , Häyrynen, S. , Jylhävä, J. , Nevalainen, T. , Hervonen, A. , … Hurme, M. (2015). Ageing‐associated changes in the human DNA methylome: Genomic locations and effects on gene expression. BMC Genomics, 16(1), 179–196. 10.1186/s12864-015-1381-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- McDermott, M. M. , Peterson, C. A. , Sufit, R. , Ferrucci, L. , Guralnik, J. M. , Kibbe, M. R. , … Leeuwenburgh, C. (2018). Peripheral artery disease, calf skeletal muscle mitochondrial DNA copy number, and functional performance. Vascular Medicine, 23(4), 340–348. 10.1177/1358863x18765667 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Melis, R. , Marengoni, A. , Angleman, S. , & Fratiglioni, L. (2014). Incidence and predictors of multimorbidity in the elderly: A population‐based longitudinal study. PLoS ONE, 9(7), e103120 10.1371/journal.pone.0103120 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mellen, M. , Ayata, P. , Dewell, S. , Kriaucionis, S. , & Heintz, N. (2012). MeCP2 binds to 5hmC enriched within active genes and accessible chromatin in the nervous system. Cell, 151(7), 1417–1430. 10.1016/j.cell.2012.11.022 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mengel‐From, J. , Thinggaard, M. , Dalgård, C. , Kyvik, K. O. , Christensen, K. , & Christiansen, L. (2014). Mitochondrial DNA copy number in peripheral blood cells declines with age and is associated with general health among elderly. Human Genetics, 133(9), 1149–1159. 10.1007/s00439-014-1458-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Menni, C. , Kiddle, S. J. , Mangino, M. , Viñuela, A. , Psatha, M. , Steves, C. , … Valdes, A. M. (2014). Circulating proteomic signatures of chronological age. The Journals of Gerontology: Series A, 70(7), 809–816. 10.1093/gerona/glu121 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Menzies, F. M. , Moreau, K. , Puri, C. , Renna, M. , & Rubinsztein, D. C. (2012). Measurement of autophagic activity in mammalian cells. Curr Protoc Cell Biol, 54(1), 15–16. 10.1002/0471143030.cb1516s54 [DOI] [PubMed] [Google Scholar]
- Miller, R. A. , Harrison, D. E. , Astle, C. M. , Baur, J. A. , Boyd, A. R. , de Cabo, R. , … Strong, R. (2010). Rapamycin, but not resveratrol or simvastatin, extends life span of genetically heterogeneous mice. The Journals of Gerontology: Series A, 66A(2), 191–201. 10.1093/gerona/glq178 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mills, E. L. , Kelly, B. , & O'Neill, L. A. J. (2017). Mitochondria are the powerhouses of immunity. Nature Immunology, 18(5), 488–498. 10.1038/ni.3704 [DOI] [PubMed] [Google Scholar]
- Moestrup, K. S. , Andersen, M. S. , & Jensen, K. B. (2017). Isolation and In Vitro Characterization of Epidermal Stem Cells. Methods Mol Biol, 1553, 67–83. 10.1007/978-1-4939-6756-8_6 [DOI] [PubMed] [Google Scholar]
- Montpetit, A. J. , Alhareeri, A. A. , Montpetit, M. , Starkweather, A. R. , Elmore, L. W. , Filler, K. , … Jackson‐Cook, C. K. (2014). Telomere length. Nursing Research, 63(4), 289–299. 10.1097/nnr.0000000000000037 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moore, A. Z. , Ding, J. , Tuke, M. A. , Wood, A. R. , Bandinelli, S. , Frayling, T. M. , & Ferrucci, L. (2017). Influence of cell distribution and diabetes status on the association between mitochondrial DNA copy number and aging phenotypes in the InCHIANTI study. Aging Cell, 17(1), e12683 10.1111/acel.12683 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moreno‐Blas, D. , Gorostieta‐Salas, E. , & Castro‐Obregón, S. (2018). Connecting chaperone‐mediated autophagy dysfunction to cellular senescence. Ageing Research Reviews, 41, 34–41. 10.1016/j.arr.2017.11.001 [DOI] [PubMed] [Google Scholar]
- Moreno‐Villanueva, M. , Capri, M. , Breusing, N. , Siepelmeyer, A. , Sevini, F. , Ghezzo, A. , … Bürkle, A. (2015). MARK‐AGE standard operating procedures (SOPs): A successful effort. Mechanisms of Ageing and Development, 151, 18–25. 10.1016/j.mad.2015.03.007 [DOI] [PubMed] [Google Scholar]
- Moreno, G. , Mangione, C. M. , Kimbro, L. , & Vaisberg, E. (2013). Guidelines abstracted from the American Geriatrics Society guidelines for improving the care of older adults with diabetes mellitus: 2013 update. Journal of the American Geriatrics Society, 61(11), 2020–2026. 10.1111/jgs.12514 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morimoto, R. I. , & Cuervo, A. M. (2014). Proteostasis and the aging proteome in health and disease. The Journals of Gerontology Series A: Biological Sciences and Medical Sciences, 69(Suppl. 1), S33–S38. 10.1093/gerona/glu049 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morselli, E. , Marino, G. , Bennetzen, M. V. , Eisenberg, T. , Megalou, E. , Schroeder, S. , … Kroemer, G. (2011). Spermidine and resveratrol induce autophagy by distinct pathways converging on the acetylproteome. Journal of Cell Biology, 192(4), 615–629. 10.1083/jcb.201008167 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Müezzinler, A. , Zaineddin, A. K. , & Brenner, H. (2013). A systematic review of leukocyte telomere length and age in adults. Ageing Research Reviews, 12(2), 509–519. 10.1016/j.arr.2013.01.003 [DOI] [PubMed] [Google Scholar]
- Mundstock, E. , Sarria, E. E. , Zatti, H. , Mattos Louzada, F. , Kich Grun, L. , Herbert Jones, M. , … Mattiello, R. (2015). Effect of obesity on telomere length: Systematic review and meta‐analysis. Obesity, 23(11), 2165–2174. 10.1002/oby.21183 [DOI] [PubMed] [Google Scholar]
- Muñoz‐Espín, D. , & Serrano, M. (2014). Cellular senescence: From physiology to pathology. Nature Reviews Molecular Cell Biology, 15(7), 482–496. 10.1038/nrm3823 [DOI] [PubMed] [Google Scholar]
- Nagel, Z. D. , Margulies, C. M. , Chaim, I. A. , McRee, S. K. , Mazzucato, P. , Ahmad, A. , … Samson, L. D. (2014). Multiplexed DNA repair assays for multiple lesions and multiple doses via transcription inhibition and transcriptional mutagenesis. Proceedings of the National Academy of Sciences, 111(18), E1823–E1832. 10.1073/pnas.1401182111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Najarro, K. , Nguyen, H. , Chen, G. , Xu, M. , Alcorta, S. , Yao, X. , … Weng, N.‐P. (2015). Telomere length as an indicator of the robustness of B‐ and T‐cell response to influenza in older adults. Journal of Infectious Diseases, 212(8), 1261–1269. 10.1093/infdis/jiv202 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Neault, M. , Couteau, F. , Bonneau, É. , De Guire, V. , & Mallette, F. A. (2017). Molecular Regulation of Cellular Senescence by MicroRNAs: Implications in Cancer and Age-Related Diseases. Int Rev Cell Mol Biol, 334, 27–98. 10.1016/bs.ircmb.2017.04.001 [DOI] [PubMed] [Google Scholar]
- Netea‐Maier, R. T. , Plantinga, T. S. , van de Veerdonk, F. L. , Smit, J. W. , & Netea, M. G. (2015). Modulation of inflammation by autophagy: Consequences for human disease. Autophagy, 12(2), 245–260. 10.1080/15548627.2015.1071759 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Newman, J. C. , Milman, S. , Hashmi, S. K. , Austad, S. N. , Kirkland, J. L. , Halter, J. B. , & Barzilai, N. (2016). Strategies and challenges in clinical trials targeting human aging. The Journals of Gerontology Series A: Biological Sciences and Medical Sciences, 71(11), 1424–1434. 10.1093/gerona/glw149 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Niccoli, T. , & Partridge, L. (2012). Ageing as a risk factor for disease. Current Biology, 22(17), R741–R752. 10.1016/j.cub.2012.07.024 [DOI] [PubMed] [Google Scholar]
- Niedernhofer, L. J. , Kirkland, J. L. , & Ladiges, W. (2017). Molecular pathology endpoints useful for aging studies. Ageing Research Reviews, 35, 241–249. 10.1016/j.arr.2016.09.012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Noren Hooten, N. , & Evans, M. K. (2017). Techniques to induce and quantify cellular senescence. Journal of Visualized Experiments, 123 10.3791/55533 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Olshansky, S. J. (2016). Articulating the case for the longevity dividend. Cold Spring Harbor Perspectives in Medicine, 6(2), a025940 10.1101/cshperspect.a025940 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Osler, M. , Bendix, L. , Rask, L. , & Rod, N. H. (2016). Stressful life events and leucocyte telomere length: Do lifestyle factors, somatic and mental health, or low grade inflammation mediate this relationship? Results from a cohort of Danish men born in 1953. Brain, Behavior, and Immunity, 58, 248–253. 10.1016/j.bbi.2016.07.154 [DOI] [PubMed] [Google Scholar]
- Palmer, A. K. , Tchkonia, T. , LeBrasseur, N. K. , Chini, E. N. , Xu, M. , & Kirkland, J. L. (2015). Cellular senescence in type 2 diabetes: A therapeutic opportunity. Diabetes, 64(7), 2289–2298. 10.2337/db14-1820 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Panda, A. C. , Abdelmohsen, K. , & Gorospe, M. (2017). SASP regulation by noncoding RNA. Mechanisms of Ageing and Development, 168, 37–43. 10.1016/j.mad.2017.05.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pang, W. W. , Price, E. A. , Sahoo, D. , Beerman, I. , Maloney, W. J. , Rossi, D. J. , … Weissman, I. L. (2011). Human bone marrow hematopoietic stem cells are increased in frequency and myeloid‐biased with age. Proceedings of the National Academy of Sciences, 108(50), 20012–20017. 10.1073/pnas.1116110108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pembrey, M. , Saffery, R. , & Bygren, L. O. (2014). Human transgenerational responses to early‐life experience: Potential impact on development, health and biomedical research. Journal of Medical Genetics, 51(9), 563–572. 10.1136/jmedgenet-2014-102577 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pietrocola, F. , Demont, Y. , Castoldi, F. , Enot, D. , Durand, S. , Semeraro, M. , … Kroemer, G. (2017). Metabolic effects of fasting on human and mouse blood in vivo. Autophagy, 13(3), 567–578. 10.1080/15548627.2016.1271513 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pratsinis, H. , Mavrogonatou, E. , & Kletsas, D. (2018). Scarless wound healing: From development to senescence. Advanced Drug Delivery Reviews, 146, 325–343. 10.1016/j.addr.2018.04.011 [DOI] [PubMed] [Google Scholar]
- Pucciarelli, S. , Moreschini, B. , Micozzi, D. , De Fronzo, G. S. , Carpi, F. M. , Polzonetti, V. , … Napolioni, V. (2012). Spermidine and spermine are enriched in whole blood of nona/centenarians. Rejuvenation Research, 15(6), 590–595. 10.1089/rej.2012.1349 [DOI] [PubMed] [Google Scholar]
- Puterman, E. , Gemmill, A. , Karasek, D. , Weir, D. , Adler, N. E. , Prather, A. A. , & Epel, E. S. (2016). Lifespan adversity and later adulthood telomere length in the nationally representative US Health and Retirement Study. Proceedings of the National Academy of Sciences, 113(42), E6335–E6342. 10.1073/pnas.1525602113 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Putin, E. , Mamoshina, P. , Aliper, A. , Korzinkin, M. , Moskalev, A. , Kolosov, A. , … Zhavoronkov, A. (2017). Deep biomarkers of human aging: Application of deep neural networks to biomarker development. Aging, 8(5), 1021–1033. 10.18632/aging.100968 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qian, Y. , Butler, T. J. , Opsahl‐Ong, K. , Giroux, N. S. , Sidore, C. , Nagaraja, R. , … Ding, J. (2017). fastMitoCalc: An ultra‐fast program to estimate mitochondrial DNA copy number from whole‐genome sequences. Bioinformatics, 33(9), 1399–1401. 10.1093/bioinformatics/btw835 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Quach, A. , Levine, M. E. , Tanaka, T. , Lu, A. T. , Chen, B. H. , Ferrucci, L. , … Horvath, S. (2017). Epigenetic clock analysis of diet, exercise, education, and lifestyle factors. Aging, 9(2), 419–446. 10.18632/aging.101168 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rae, M. J. , Butler, R. N. , Campisi, J. , de Grey, A. D. N. J. , Finch, C. E. , Gough, M. , … Logan, B. J. (2010). The demographic and biomedical case for late‐life interventions in aging. Science Translational Medicine, 2(40), 40cm21–40cm21. 10.1126/scitranslmed.3000822 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raz, Y. , Guerrero‐Ros, I. , Maier, A. , Slagboom, P. E. , Atzmon, G. , Barzilai, N. , & Macian, F. (2017). Activation‐induced autophagy is preserved in CD4+ T‐cells in familial longevity. Journals of Gerontology. Series A, Biological Sciences and Medical Sciences, 72(9), 1201–1206. 10.1093/gerona/glx020 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reddy, P. T. , Jaruga, P. , Nelson, B. C. , Lowenthal, M. S. , Jemth, A.‐S. , Loseva, O. , … Dizdaroglu, M. (2016). Production, Purification, and Characterization of ¹⁵N-Labeled DNA Repair Proteins as Internal Standards for Mass Spectrometric Measurements. Methods Enzymol, 566, 305–332. 10.1016/bs.mie.2015.06.044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ren, R. , Ocampo, A. , Liu, G.‐H. , & Izpisua Belmonte, J. C. (2017). Regulation of stem cell aging by metabolism and epigenetics. Cell Metabolism, 26(3), 460–474. 10.1016/j.cmet.2017.07.019 [DOI] [PubMed] [Google Scholar]
- Richardson, A. , Fischer, K. E. , Speakman, J. R. , de Cabo, R. , Mitchell, S. J. , Peterson, C. A. , … Austad, S. N. (2015). Measures of healthspan as indices of aging in mice—A recommendation. The Journals of Gerontology Series A: Biological Sciences and Medical Sciences, 71(4), 427–430. 10.1093/gerona/glv080 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rigotti, G. , Charles‐de‐Sá, L. , Gontijo‐de‐Amorim, N. F. , Takiya, C. M. , Amable, P. R. , Borojevic, R. , … Sbarbati, A. (2016). Expanded stem cells, stromal‐vascular fraction, and platelet‐rich plasma enriched fat: Comparing results of different facial rejuvenation approaches in a clinical trial. Aesthetic Surgery Journal, 36(3), 261–270. 10.1093/asj/sjv231 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robbins, P. D. , & Niedernhofer, L. J. (2017). Advances in therapeutic approaches to extend healthspan: A perspective from the 2nd Scripps Symposium on the Biology of Aging. Aging Cell, 16(4), 610–614. 10.1111/acel.12620 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rocca, W. A. , Boyd, C. M. , Grossardt, B. R. , Bobo, W. V. , Finney Rutten, L. J. , Roger, V. L. , … St. Sauver, J. L. (2014). Prevalence of multimorbidity in a geographically defined american population. Mayo Clinic Proceedings, 89(10), 1336–1349. 10.1016/j.mayocp.2014.07.010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rodier, F. , & Campisi, J. (2011). Four faces of cellular senescence. The Journal of Cell Biology, 192(4), 547–556. 10.1083/jcb.201009094 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rossi, L. , Challen, G. A. , Sirin, O. , Lin, K. K. , & Goodell, M. A. (2011). Hematopoietic stem cell characterization and isolation. Methods Mol Biol, 750, 47–59. 10.1007/978-1-61779-145-1_3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rossiello, F. , Aguado, J. , Sepe, S. , Iannelli, F. , Nguyen, Q. , Pitchiaya, S. , … d’Adda di Fagagna, F. (2017). DNA damage response inhibition at dysfunctional telomeres by modulation of telomeric DNA damage response RNAs. Nature Communications, 8, 13980 10.1038/ncomms13980 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ryu, D. , Mouchiroud, L. , Andreux, P. A. , Katsyuba, E. , Moullan, N. , Nicolet‐Dit‐Felix, A. A. , … Auwerx, J. (2016). Urolithin A induces mitophagy and prolongs lifespan in C. elegans and increases muscle function in rodents. Nature Medicine, 22(8), 879–888. 10.1038/nm.4132 [DOI] [PubMed] [Google Scholar]
- Saha, D. T. , Davidson, B. J. , Wang, A. , Pollock, A. J. , Orden, R. A. , & Goldman, R. (2008). Quantification of DNA repair capacity in whole blood of patients with head and neck cancer and healthy donors by comet assay. Mutation Research/Genetic Toxicology and Environmental Mutagenesis, 650(1), 55–62. 10.1016/j.mrgentox.2007.10.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanada, F. , Taniyama, Y. , Muratsu, J. , Otsu, R. , Shimizu, H. , Rakugi, H. , & Morishita, R. (2018). Source of chronic inflammation in aging. Frontiers in Cardiovascular Medicine, 5, 12 10.3389/fcvm.2018.00012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanders, J. L. , & Newman, A. B. (2013). Telomere length in epidemiology: A biomarker of aging, age‐related disease, both, or neither? Epidemiologic Reviews, 35(1), 112–131. 10.1093/epirev/mxs008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saretzki, G. (2018). Telomeres, telomerase and ageing. SubCellular Biochemistry, 90, 221–308. 10.1007/978-981-13-2835-0_9 [DOI] [PubMed] [Google Scholar]
- Sehl, M. E. , Henry, J. E. , Storniolo, A. M. , Ganz, P. A. , & Horvath, S. (2017). DNA methylation age is elevated in breast tissue of healthy women. Breast Cancer Research and Treatment, 164(1), 209–219. 10.1007/s10549-017-4218-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Settembre, C. , Fraldi, A. , Rubinsztein, D. C. , & Ballabio, A. (2008). Lysosomal storage diseases as disorders of autophagy. Autophagy, 4(1), 113–114. 10.4161/auto.5227 [DOI] [PubMed] [Google Scholar]
- Shavlakadze, T. , Zhu, J. , Wang, S. , Zhou, W. , Morin, B. , Egerman, M. A. , … Glass, D. J. (2018). Short‐term low‐dose mTORC1 inhibition in aged rats counter‐regulates age‐related gene changes and blocks age‐related kidney pathology. Journals of Gerontology. Series A, Biological Sciences and Medical Sciences, 73(7), 845–852. 10.1093/gerona/glx249 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shen, J.‐C. , Fox, E. J. , Ahn, E. H. , & Loeb, L. A. (2014). A rapid assay for measuring nucleotide excision repair by oligonucleotide retrieval. Scientific Reports, 4(1), 4894 10.1038/srep04894 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sidler, C. , Kovalchuk, O. , & Kovalchuk, I. (2017). Epigenetic regulation of cellular senescence and aging. Frontiers in Genetics, 8, 10.3389/fgene.2017.00138 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sierra, F. (2016). The emergence of geroscience as an interdisciplinary approach to the enhancement of health span and life span. Cold Spring Harbor Perspectives in Medicine, 6(4), a025163 10.1101/cshperspect.a025163 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sillanpää, E. , Laakkonen, E. K. , Vaara, E. , Rantanen, T. , Kovanen, V. , Sipilä, S. , … Ollikainen, M. (2018). Biological clocks and physical functioning in monozygotic female twins. BMC Geriatrics, 18(1), 83–90. 10.1186/s12877-018-0775-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Solomon, A. , Tennakoon, S. , Leeansyah, E. , Arribas, J. , Hill, A. , Van Delft, Y. , … Lewin, S. R. (2014). No difference in the rate of change in telomere length or telomerase activity in HIV‐infected patients after three years of darunavir/ritonavir with and without nucleoside analogues in the MONET trial. PLoS ONE, 9(11), e109718 10.1371/journal.pone.0109718 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spaak, J. (2017). Novel combined management approaches to patients with diabetes, chronic kidney disease and cardiovascular disease. Journal of the Royal College of Physicians of Edinburgh, 47(1), 83–87. 10.4997/jrcpe.2017.118 [DOI] [PubMed] [Google Scholar]
- Spiegel, A. M. , Sewal, A. S. , & Rapp, P. R. (2014). Epigenetic contributions to cognitive aging: Disentangling mindspan and lifespan. Learning & Memory, 21(10), 569–574. 10.1101/lm.033506.113 [DOI] [PMC free article] [PubMed] [Google Scholar]
- St Sauver, J. L. , Boyd, C. M. , Grossardt, B. R. , Bobo, W. V. , Finney Rutten, L. J. , Roger, V. L. , … Rocca, W. A. (2015). Risk of developing multimorbidity across all ages in an historical cohort study: Differences by sex and ethnicity. British Medical Journal Open, 5(2), e006413–e006413. 10.1136/bmjopen-2014-006413 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Starkov, A. A. (2010). Measurement of mitochondrial ROS production. Methods Mol Biol, 648, 245–255. 10.1007/978-1-60761-756-3_16 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Strzyz, P. (2016). Controlling the senescence‐associated secretory phenotype. Nature Reviews Molecular Cell Biology, 17(12), 740–740. 10.1038/nrm.2016.157 [DOI] [PubMed] [Google Scholar]
- Szulwach, K. E. , Li, X. , Li, Y. , Song, C. X. , Wu, H. , Dai, Q. , … Vasanthakumar, A. (2011). 5-hmC–mediated epigenetic dynamics during postnatal neurodevelopment and aging. Nature neuroscience, 14(12), 1607–16. 10.1038/nn.2959 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Szwagierczak, A. , Bultmann, S. , Schmidt, C. S. , Spada, F. , & Leonhardt, H. (2010). Sensitive enzymatic quantification of 5-hydroxymethylcytosine in genomic DNA. Nucleic acids research, 38(19), e181–e181. 10.1093/nar/gkq684 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tahiliani, M. , Koh, K. P. , Shen, Y. , Pastor, W. A. , Bandukwala, H. , Brudno, Y. , … Rao, A. (2009). Conversion of 5‐methylcytosine to 5‐hydroxymethylcytosine in mammalian DNA by MLL partner TET1. Science, 324(5929), 930–935. 10.1126/science.1170116 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tanaka, T. , Biancotto, A. , Moaddel, R. , Moore, A. Z. , Gonzalez‐Freire, M. , Aon, M. A. , … Ferrucci, L. (2018). Plasma proteomic signature of age in healthy humans. Aging Cell, 17(5), e12799 10.1111/acel.12799 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Terragni, J. , Bitinaite, J. , Zheng, Y. , & Pradhan, S. (2012). Biochemical characterization of recombinant β-glucosyltransferase and analysis of global 5-hydroxymethylcytosine in unique genomes. Biochemistry, 51(5), 1009–1019. 10.1021/bi2014739 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tisminetzky, M. , Bayliss, E. A. , Magaziner, J. S. , Allore, H. G. , Anzuoni, K. , Boyd, C. M. , … Gurwitz, J. H. (2017). Research priorities to advance the health and health care of older adults with multiple chronic conditions. Journal of the American Geriatrics Society, 65(7), 1549–1553. 10.1111/jgs.14943 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tomaska, L. , & Nosek, J. (2009). Telomere heterogeneity: Taking advantage of stochastic events. FEBS Letters, 583(7), 1067–1071. 10.1016/j.febslet.2009.02.032 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tompkins, B. A. , DiFede, D. L. , Khan, A. , Landin, A. M. , Schulman, I. H. , Pujol, M. V. , … Hare, J. M. (2017). Allogeneic mesenchymal stem cells ameliorate aging frailty: A phase II randomized, double‐blind, placebo‐controlled clinical trial. The Journals of Gerontology: Series A, 72(11), 1513–1522. 10.1093/gerona/glx137 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trzeciak, A. R. , Barnes, J. , Ejiogu, N. , Foster, K. , Brant, L. J. , Zonderman, A. B. , & Evans, M. K. (2008). Age, sex, and race influence single‐strand break repair capacity in a human population. Free Radical Biology and Medicine, 45(12), 1631–1641. 10.1016/j.freeradbiomed.2008.08.031 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trzeciak, A. R. , Barnes, J. , & Evans, M. K. (2008). A modified alkaline comet assay for measuring DNA repair capacity in human populations. Radiation Research, 169(1), 110–121. 10.1667/rr1101.1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ungar, R. A. , Giri, N. , Pao, M. , Khincha, P. P. , Zhou, W. , Alter, B. P. , & Savage, S. A. (2018). Complex phenotype of dyskeratosis congenita and mood dysregulation with novel homozygous RTEL1 and TPH1 variants. American Journal of Medical Genetics Part A, 176(6), 1432–1437. 10.1002/ajmg.a.38706 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Valentini, E. , Zampieri, M. , Malavolta, M. , Bacalini, M. G. , Calabrese, R. , Guastafierro, T. , … Bernhardt, J. (2016). Analysis of the machinery and intermediates of the 5hmC-mediated DNA demethylation pathway in aging on samples from the MARK-AGE Study. Aging (Albany NY), 8(9), 1896–1922. 10.18632/aging.101022 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vera, E. , Bernardes de Jesus, B. , Foronda, M. , Flores, J. M. , & Blasco, M. A. (2012). The rate of increase of short telomeres predicts longevity in mammals. Cell Reports, 2(4), 732–737. 10.1016/j.celrep.2012.08.023 [DOI] [PubMed] [Google Scholar]
- Vetrano, D. L. , Calderón‐Larrañaga, A. , Marengoni, A. , Onder, G. , Bauer, J. M. , Cesari, M. , … Fratiglioni, L. (2017). An International perspective on chronic multimorbidity: Approaching the ELEPHANT IN THE ROOm. The Journals of Gerontology: Series A, 73(10), 1350–1356. 10.1093/gerona/glx178 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Volarevic, V. , Ljujic, B. , Stojkovic, P. , Lukic, A. , Arsenijevic, N. , & Stojkovic, M. (2011). Human stem cell research and regenerative medicine–present and future. British Medical Bulletin, 99(1), 155–168. 10.1093/bmb/ldr027 [DOI] [PubMed] [Google Scholar]
- Volarevic, V. , Markovic, B. S. , Gazdic, M. , Volarevic, A. , Jovicic, N. , Arsenijevic, N. , … Stojkovic, M. (2018). Ethical and safety issues of stem cell‐based therapy. International Journal of Medical Sciences, 15(1), 36–45. 10.7150/ijms.21666 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Waaijer, M. E. , Parish, W. E. , Strongitharm, B. H. , van Heemst, D. , Slagboom, P. E. , de Craen, A. J. , … Maier, A. B. (2012). The number of p16INK4a positive cells in human skin reflects biological age. Aging Cell, 11(4), 722–725. 10.1111/j.1474-9726.2012.00837.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wachsmuth, M. , Hübner, A. , Li, M. , Madea, B. , & Stoneking, M. (2016). Age‐related and heteroplasmy‐related variation in human mtDNA copy number. PLOS Genetics, 12(3), e1005939 10.1371/journal.pgen.1005939 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wadhwa, P. , Buss, C. , Entringer, S. , & Swanson, J. (2009). Developmental origins of health and disease: Brief history of the approach and current focus on epigenetic mechanisms. Seminars in Reproductive Medicine, 27(05), 358–368. 10.1055/s-0029-1237424 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang, L. , Huang, J. , Pang, S. , Qin, X. , Qi, Z. , Hawley, R. G. , & Yan, B. (2017). Genetic analysis of the ATG16L1 gene promoter in sporadic Parkinson's disease. Neuroscience Letters, 646, 30–35. 10.1016/j.neulet.2017.03.007 [DOI] [PubMed] [Google Scholar]
- Waters, D. W. , Blokland, K. E. C. , Pathinayake, P. S. , Burgess, J. K. , Mutsaers, S. E. , Prele, C. M. , … Knight, D. A. (2018). Fibroblast senescence in the pathology of idiopathic pulmonary fibrosis. American Journal of Physiology‐Lung Cellular and Molecular Physiology, 315(2), L162–L172. 10.1152/ajplung.00037.2018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weber, D. , Stuetz, W. , Toussaint, O. , Debacq‐Chainiaux, F. , Dollé, M. E. T. , Jansen, E. , … Grune, T. (2017). Associations between specific redox biomarkers and age in a large European Cohort: The MARK‐AGE project. Oxidative Medicine and Cellular Longevity, 2017, 1–12. 10.1155/2017/1401452 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wen, L. , Li, X. , Yan, L. , Tan, Y. , Li, R. , Zhao, Y. , … Yu, M. (2014). Whole-genome analysis of 5-hydroxymethylcytosine and 5-methylcytosine at base resolution in the human brain. Genome Biology, 15(3), R49 10.1186/gb-2014-15-3-r49 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Whittemore, K. , Vera, E. , Martinez‐Nevado, E. , Sanpera, C. , & Blasco, M. A. (2019). Telomere shortening rate predicts species life span. Proceedings of the National Academy of Sciences of the United States of America, 116(30), 15122–15127. 10.1073/pnas.1902452116 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wiley, C. D. , & Campisi, J. (2016). From ancient pathways to aging cells—Connecting metabolism and cellular senescence. Cell Metabolism, 23(6), 1013–1021. 10.1016/j.cmet.2016.05.010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wiley, C. D. , Velarde, M. C. , Lecot, P. , Liu, S. , Sarnoski, E. A. , Freund, A. , … Campisi, J. (2016). Mitochondrial dysfunction induces senescence with a distinct secretory phenotype. Cell Metabolism, 23(2), 303–314. 10.1016/j.cmet.2015.11.011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wong, E. , & Cuervo, A. M. (2010). Integration of clearance mechanisms: The proteasome and autophagy. Cold Spring Harbor Perspectives in Biology, 2(12), a006734–a006734. 10.1101/cshperspect.a006734 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xia, X. , Chen, W. , McDermott, J. , & Han, J.‐D.‐J. (2017). Molecular and phenotypic biomarkers of aging. F1000Research, 6, 860 10.12688/f1000research.10692.1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoshii, S. R. , & Mizushima, N. (2017). Monitoring and measuring autophagy. International Journal of Molecular Sciences, 18(9), 1865 10.3390/ijms18091865 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Young, J. I. , Hong, E. P. , Castle, J. C. , Crespo‐Barreto, J. , Bowman, A. B. , Rose, M. F. , … Zoghbi, H. Y. (2005). Regulation of RNA splicing by the methylation‐dependent transcriptional repressor methyl‐CpG binding protein 2. Proceedings of the National Academy of Sciences of the United States of America, 102(49), 17551–17558. 10.1073/pnas.0507856102 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yousefzadeh, M. J. , Schafer, M. J. , Noren Hooten, N. , Atkinson, E. J. , Evans, M. K. , Baker, D. J. , … Niedernhofer, L. J. (2017). Circulating levels of monocyte chemoattractant protein‐1 as a potential measure of biological age in mice and frailty in humans. Aging Cell, 17(2), e12706 10.1111/acel.12706 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu, M. , Hon, G. C. , Szulwach, K. E. , Song, C. X. , Zhang, L. , Kim, A. , … He, C. (2012). Base-resolution analysis of 5-hydroxymethylcytosine in the mammalian genome. Cell, 149(6), 1368–1380. 10.1016/j.cell.2012.04.027 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zane, A. C. , Reiter, D. A. , Shardell, M. , Cameron, D. , Simonsick, E. M. , Fishbein, K. W. , … Ferrucci, L. (2017). Muscle strength mediates the relationship between mitochondrial energetics and walking performance. Aging Cell, 16(3), 461–468. 10.1111/acel.12568 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao, J. , Zhu, Y. , Yang, J. , Li, L. , Wu, H. , De Jager, P. L. , … Bennett, D. A. (2017). A genome-wide profiling of brain DNA hydroxymethylation in Alzheimer's disease. Alzheimer's & Dementia, 13(6), 674–688. 10.1016/j.jalz.2016.10.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang, H. , Puleston, D. J. , & Simon, A. K. (2016). Autophagy and immune senescence. Trends in Molecular Medicine, 22(8), 671–686. 10.1016/j.molmed.2016.06.001 [DOI] [PubMed] [Google Scholar]
- Zhang, K. , Chen, C. , Liu, Y. , Chen, H. , & Liu, J.‐P. (2014). Cellular senescence occurred widespread to multiple selective sites in the fetal tissues and organs of mice. Clinical and Experimental Pharmacology and Physiology, 41(12), 965–975. 10.1111/1440-1681.12328 [DOI] [PubMed] [Google Scholar]
- Zhang, L. , Dong, X. , Lee, M. , Maslov, A. Y. , Wang, T. , & Vijg, J. (2019). Single‐cell whole‐genome sequencing reveals the functional landscape of somatic mutations in B lymphocytes across the human lifespan. Proceedings of the National Academy of Sciences of the United States of America, 116(18), 9014–9019. 10.1073/pnas.1902510116 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang, P. , Moudgill, N. , Hager, E. , Tarola, N. , DiMatteo, C. , McIlhenny, S. , … DiMuzio, P. J. (2011). Endothelial differentiation of adipose‐derived stem cells from elderly patients with cardiovascular disease. Stem Cells and Development, 20(6), 977–988. 10.1089/scd.2010.0152 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang, R. , Wang, Y. , Ye, K. , Picard, M. , & Gu, Z. (2017). Independent impacts of aging on mitochondrial DNA quantity and quality in humans. BMC Genomics, 18(1), 890. [DOI] [PMC free article] [PubMed] [Google Scholar]