

Abstract
Background: Diffuse large B-cell lymphoma (DLBCL) is considered the most common aggressive subtype of lymphoma. A number of DLBCL patients fail to achieve a response to currently available therapies or develop resistance. β-Elemene is derived from herb Curcuma wenyujin, and exhibits anti-tumor activity in both solid and non-solid tumors through modulating several molecular signaling pathways. We aimed to explore the role of β-elemene in DLBCL treatment and elucidate the involved mechanism.
Materials and methods: Cell viability, apoptosis and expressions of related proteins were assessed and in vivo study were performed to determine the tumor suppressive effect of β-elemene and explore the molecular mechanisms.
Results: β-Elemene significantly suppressed the viability of DLBCL cells, and β-elemene down-regulated the lncRNA HULC expression and regulated key pro-apoptotic and anti-apoptotic proteins to induce significant apoptosis of DLBCL cells. HULC overexpression could decrease the β-elemene induced apoptosis, while HULC knockdown increased the apoptosis in DLBCL cells. In vivo study further confirmed that β-elemene could suppress the growth of DLBCL xenograft and regulate the HULC expression and the critical proteins of the apoptotic pathway.
Conclusion: β-Elemene performs as a tumor suppressor and modulator of HULC-mediated apoptotic pathway in DLBCL and will be an alternative candidate for clinical application.
Keywords: β-elemene, Apoptosis, DLBCL, HULC, lncRNA
Introduction
It is known that diffuse large B-cell lymphoma (DLBCL) is regarded as the most common aggressive subtype of non-Hodgkin lymphoma (NHL). DLBCL represents the leading cause of disease-specific mortality and accounts for approximately 40% of all newly diagnosed NHL cases [1]. DLBCL patients have a variable clinical response to conventional therapeutic treatments, ranging from cure to failure, refractory disease or death. Although the cure rate of localized DLBCL is high, the cure rate of advanced disease is still less than 50% [2]. Recently, advances in novel therapeutic regimens, such as combined chemoimmunotherapy, stem cell transplantation, have greatly improved the overall survival of DLBCL patients. However, 30–40% of DLBCL patients fail to achieve a response to currently available therapies or develop resistance [3]. Therefore, more effective therapeutic agents based on novel targets and oncogenic signaling pathways are necessarily needed.
Recently, numerous active anti-tumor compounds derived from natural herbs have been identified and developed for clinical application. Natural herbs derived compounds show pleiotropic biological activities including anti-carcinogenic activity and fewer side effects [4–12]. β-Elemene (C15H24) is derived from herb Curcuma wenyujin, and exhibits a wide spectrum of biological functions including anti-microbial, anti-inflammatory, anti-atherosclerosis and anti-tumor. Previous studies have shown that β-elemene exerted anti-proliferative in both solid and non-solid tumors through modulating several molecular signaling pathways [13–15]. However, the effects of β-elemene on treating lymphoma, especially against DLBCL, and the possible molecular mechanisms remain unelucidated.
Long non-coding RNA (lncRNA) that is greater than 200 nucleotides has been reported to be associated with the development of lymphoma, including DLBCL [16–18]. Aberrant expression of lncRNA is associated with DLBCL occurrence and development. Highly up-regulated in liver cancer (HULC), located on chromosome 6p24.3, is a 500 nt-nucleotide-long lncRNA which is first found overexpressed in liver cancer and is closely related to the tumor progression. HULC is also aberrantly up-regulated in a wide spectrum of malignancies, including liver cancer, prostate cancer, ovarian cancer, bladder cancer, osteosarcoma, gastric cancer, and myeloid leukemia [19–25]. A meta-analysis indicated that HULC overexpression is a predictor of poor prognosis in various cancer types and the higher incidence of tumor metastasis [26]. Up-regulated HULC promotes cellular proliferation, migration, and invasion, while silencing of HULC inhibits tumor growth and enhances chemotherapy-induced apoptosis [24,25]. HULC predicted poor clinical outcome and represented pro-oncogenic activity in DLBCL [27]. Therefore, HULC could serve as a therapeutic target for DLBCL treatment.
In the present study, the antitumor effect of β-elemene in DLBCL was investigated and the potential mechanism through regulation of lncRNA HULC-mediated apoptotic pathway was examined. Our study might provide a new insight in the DLBCL treatment.
Materials and methods
Chemicals
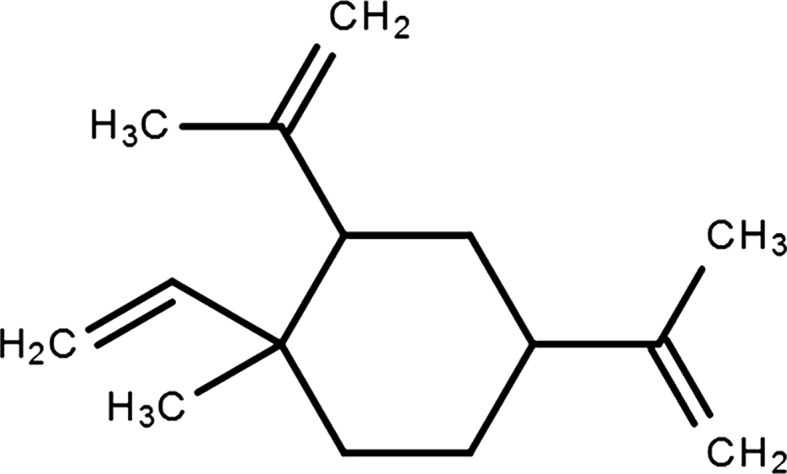
β-Elemene (Figure 1) was purchased from Holley Kingkong Pharmaceutical Co. (Dalian, China). Fetal bovine serum was purchased from Gibco (MA, U.S.A.). RPMI-1640 medium was purchased from Hyclon (UT, U.S.A.). Annexin V-FITC apoptosis kit was purchased from Abcam (MA, U.S.A.). Bax, Bcl-2 antibodies and GAPDH were purchased from Santa Cruz Biotechnology (CA, U.S.A.).
Figure 1. The structural formula of β-elemene.
Cell lines
Human diffuse large B-cell lymphoma cell lines SU-DHL-8, SU-DHL-10 were purchased from American Type Culture Collection (VA, U.S.A.). Cells were maintained in RPMI-1640 medium supplemented with 10% fetal bovine serum at 37°C in a humidified incubator containing 5% CO2.
Cell proliferative inhibition and clonogenic assay
The cytotoxicity of β-elemene on DLBCL cells was measured by MTT assay. Cells (1 × 104 cells/well) were seeded into 96-well plates overnight and then exposed to various concentrations of β-elemene for 12, 24, 48 h. The microplate reader was used to measure the absorbance at 550 nm wavelength and calculate the cell viability rates.
Cells (1 × 103 cells/well) were seeded into 6-well plates and cultured overnight, then exposed to increasing concentrations of β-elemene for 48 h. Cells were cultured in RPMI-1640 medium for further 2 weeks. At the end of the incubation, the colonies were stained and scored under an optical inverted microscope.
Cell apoptosis analysis
The apoptotic rate of DLBCL cells induced by β-elemene was measured by the Annexin-V/PI apoptosis assay. Cells (5 × 104 cells/well) were seeded into 6-well plates overnight and then exposed to various concentrations of β-elemene for 48 h. Cells were stained with FITC and PI according to the manufacturer’s protocol. Apoptotic cells were calculated as either FITC+/PI− staining or FITC+/PI+ staining by flow cytometry.
Quantitative real-time transcription-polymerase chain reaction (qRT-PCR)
Total RNAs were extracted from cells by Trizol reagent. qRT-PCR was performed and all kits were used following the manufacturer’s protocol. The primes of HULC: forward, 5′-ACCTCCAGAACTGTGATCCAAAATG-3′, reverse, 5′-TCTTGCTTGATGCTTTGGTCTG-3′. β-Actin was used as endogenous control.
Cell transfection
HULC full-length cDNA was amplified using PCR and HULC was knocked down using a specific hairpin shRNA. The shRNA sequences for HULC were 5′-GAAGTAAAGGCCGGAATATTC-3′(shHULC-1), 5′-GGAATTGGAGCCTTTACAAGG-3′(shHULC-2). Then, the objective products were cloned into a lentiviral plasmid and infected into cells according to the manufacturer’s protocol.
Western blot analysis
Cells were exposed to β-elemene (60 μg/ml) for 48 h and washed with PBS and lysed in a RIPA buffer. The lysates were centrifuged (10,000 rpm, 10 min, 4°C) and the supernatant was collected. Equivalent amounts of protein were loaded and resolved on 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis. Then, the protein was transferred onto a polyvinylidene fluoride membrane which was blocked with TBS-T containing 5% defatted milk. The proteins on the membranes were incubated with Bax, Bcl-2 primary antibodies (1:1000) at 4°C overnight, and then incubated with HRP-conjugated secondary antibody (1:2000) at room temperature for 60 min. The enhanced chemiluminescence system was used to visualize the bands.
Nude mice xenograft study
The animal experiment was performed at Zhejiang Hospital following the NIH guidelines. The protocol was approved by the ethics committee of Zhejiang Hospital (Hangzhou, China). Male BALB/C mice (4-week-old) were purchased from Shanghai SLAC Laboratory Animal Co. (Shanghai, China). The establishment of DLBCL xenograft was carried out according to the protocol previously described [28,29]. Briefly, the SU-DHL-8 cells (1 × 107 cells) were planted into the upper flank region of mice subcutaneously. Tumor sizes were calculated by the formula (width)2 × length/2. When the average tumor size reached 0.2 cm3, β-elemene (45 mg/kg) was injected into nude mice intraperitoneally once a day. Saline was used in the control group accordingly. Body weight of nude mice and tumor volume were scored every week. Finally, mice were killed by cervical dislocation at day 28 and tumor samples were collected.
Immunohistochemistry
Immunohistochemistry was used to determine the expression of Bax and Bcl-2. Paraffin slides (4 μm-thick) were deparaffinized and rehydrated. The slides were under 98°C, 10 min for antigen retrieval prior to incubation with endogenous peroxidase blocking solution. The slides were then incubated with Bax and Bcl-2 antibodies at 4°C overnight, and then incubated with secondary antibody for 1 h. DAB substrate was used and all slides were counterstained with hematoxylin for 4 min. The number of immunopositive cells was identified as positive cells (200× magnification).
Statistical analysis
The data were presented as mean ± SD from triplicated experiments. Statistical analyses were performed using SPSS version 17.0 (SPSS Inc., Chicago, IL, U.S.A.). One way analysis of variance and SNK-q test were used to assess the differences between groups. Statistical significantce was considered as p < 0.05.
Results
β-Elemene suppressed the viability of DLBCL cells
Cell viability of SU-DHL-8 and SU-DHL-10 cells exposed to β-elemene was determined by MTT assay. It showed that β-elemene suppressed the viability of SU-DHL-8 and SU-DHL-10 cells in a dose- and time-dependent manner (Figure 2A). Clonogenic assay also showed that the colony formation ability of SU-DHL-8 and SU-DHL-10 cells were inhibited by β-elemene (Figure 2B). The results showed the anti-proliferative effect of β-elemene on DLBCL cells.
Figure 2. Antitumor effect of β-elemene on the viability of DLBCL cells.

(A) β-Elemene significantly inhibited the viability of SU-DHL-8 and SU-DHL-10 cells in a dose- and time-dependent manner. (B) β-Elemene significantly inhibited the colony formation of SU-DHL-8 and SU-DHL-10 cells in a dose-dependent manner at 48 h. (C) β-Elemene significantly induced the apoptosis of SU-DHL-8 and SU-DHL-10 cells in a dose-dependent manner at 48 h. *P < 0.05 between β-elemene and control group.
β-Elemene triggered apoptosis of DLBCL cells
The apoptosis of SU-DHL-8 and SU-DHL-10 cells exposed to β-elemene was further evaluated. The results showed that β-elemene triggered the apoptosis of SU-DHL-8 and SU-DHL-10 cells in a dose-dependent manner (Figure 2C). These data indicated that β-elemene triggered significant apoptosis of DLBCL cells through an apoptotic pathway.
β-Elemene down-regulated HULC and activated apoptotic pathway
We further investigated the effect of β-elemene on HULC and key pro-apoptotic and anti-apoptotic proteins in SU-DHL-8 and SU-DHL-10 cells. It showed that β-elemene significantly down-regulated the HULC expression in a dose-dependent manner at 48 h (Figure 3A). The result of Western blot analysis showed the up-regulated Bax expression and down-regulated Bcl-2 expression by β-elemene in SU-DHL-8 and SU-DHL-10 cells at 48 h (Figure 3B).
Figure 3. β-Elemene down-regulated HULC and activated apoptotic pathway in DLBCL cells.

(A) β-Elemene significantly down-regulated the HULC expression in a dose-dependent manner. *P < 0.05 between β-elemene and control group. (B) β-Elemene (60 μg/ml) up-regulated Bax expression, while down-regulated Bcl-2 expression in SU-DHL-8 and SU-DHL-10 cells at 48 h.
To explore the role of HULC in β-elemene induced apoptosis in DLBCL cells, SU-DHL-8 cells were infected with lentivirus to stably overexpress HULC or knockdown of HULC (Figure 4A). The results showed that the apoptosis rate decreased in SU-DHL-8 cells with HULC overexpression, while no significant change was found in the cells with HULC knockdown, which were exposed to β-elemene (60 μg/ml) at 48 h (Figure 4B). The Bax expression decreased, while Bcl-2 expression increased in the β-elemene (60 μg/ml) treated SU-DHL-8 cells with HULC overexpression, and the Bax expression increased, while Bcl-2 expression decreased in the β-elemene (60 μg/ml) treated HULC knockdown SU-DHL-8 cells (Figure 4C).
Figure 4. β-Elemene induced apoptosis through HULC-mediated apoptotic pathway in DLBCL cells.

(A) HULC was up-regulated after transfection and down-regulated by knockdown of HULC in SU-DHL-8 cells. *P < 0.05 between β-elemene and control group. (B) Apoptosis rate was decreased in HULC overexpression SU-DHL-8 cells treated by β-elemene (60 μg/ml) and no significant change was found in HULC knockdown SU-DHL-8 cells treated by β-elemene (60 μg/ml) at 48 h. *P < 0.05 between HULC and vector group. #P > 0.05 between shHULC and shNC group. (C) The expression of Bax decreased, while Bcl-2 increased in HULC overexpression SU-DHL-8 cells treated by β-elemene (60 μg/ml), and the expression of Bax increased, while Bcl-2 decreased in HULC knockdown SU-DHL-8 cells treated by β-elemene (60 μg/ml) at 48 h.
β-Elemene suppressed the growth of DLBCL xenograft through regulating HULC-mediated apoptotic pathway in vivo
SU-DHL-8 xenograft was established to evaluate the antitumor effect of β-elemene in vivo. It showed that β-elemene suppressed the tumor growth of SU-DHL-8 xenograft without mice body weight changed (Figure 5A). qRT-PCR showed that the expression of HULC significantly reduced (Figure 5B), and immunohistochemical analysis showed that the Bax expression increased, while Bcl-2 expression decreased by β-elemene treatment (Figure 5C).
Figure 5. β-Elemene inhibited tumor growth of DLBCL xenograft.

(A) β-Elemene inhibited tumor volume and weight of SU-DHL-8 xenograft without mice body weight changed. *P<0.05 between β-elemene and control group. #P > 0.05 between β-elemene and control group. (B) β-Elemene down-regulated the HULC expression in vivo. *P<0.05 between β-elemene and control group. (C) Immunohistochemical analysis showed β-elemene increased Bax expression, while decreased Bcl-2 expression in vivo. *P < 0.05 between β-elemene and control group.
Discussion
Cancer research and recent studies implicated that various lncRNAs are dysregulated in human malignancies. Aberrant expression of lncRNA is associated with the DLBCL patient’s clinical characteristics and poor prognosis and can be applied to serve as a therapeutic target [16–18]. LncRNA HULC has been reported to play a crucial role in tumor carcinogenesis and progression. High expression of HULC was positively associated with advanced malignancies and poor survival of patients. HULC promotes tumor growth and progression in several cancers by mediating multiple signaling pathways and interacting with miRNAs. It has been indicated that HULC accelerated liver cancer by inhibiting PTEN via autophagy cooperation to miR15a and increasing HMGA2 expression via sequestration of the miR186 [19,30]. HULC modulated the phosphorylation of YB-1 through serving as a scaffold of extracellular signal-regulated kinase and YB-1 to enhance hepatocarcinogenesis [31]. HULC promoted proliferation and osteogenic differentiation of bone mesenchymal stem cells via down-regulation of miR-195 [32]. HULC mediated radioresistance via autophagy in prostate cancer cells [33]. HULC functioned as an oncogene by targeting ATG7 and ITGB1 in epithelial ovarian carcinoma [21]. HULC promoted bladder cancer cells proliferation but inhibited apoptosis via regulation of ZIC2 and PI3K/AKT signaling pathway [22].
Therefore, lncRNA HULC may be a therapeutic target for cancer therapy. Knockdown of HULC could inhibit proliferation, migration, invasion, and promote apoptosis by sponging miR-122 in osteosarcoma [23]. Silencing of HULC enhanced chemotherapy-induced apoptosis in gastric cancer [24]. HULC silencing suppressed angiogenesis by regulating ESM-1 via the PI3K/Akt/mTOR signaling pathway in human gliomas [34]. Recently, a growing number of novel treatment strategies, such as compounds derived from natural products, have been developed for cancer therapy by targeting lncRNAs [35–37]. Previous study reported that β-elemene inhibited the growth of esophageal cancer by modulating lncRNA-mediated inhibition of hTERT [38]. However, the role of HULC in β-elemene induced apoptosis in DLBCL remains unclear.
In the present study, we evaluated the antitumor effect of β-elemene and preliminarily elucidated the HULC involved mechanism. The results revealed that β-elemene significantly suppressed the viability of DLBCL cells in a dose- and time-dependent manner. β-Elemene down-regulated the lncRNA HULC expression and regulated key pro-apoptotic and anti-apoptotic proteins to induce significant apoptosis of DLBCL cells. HULC overexpression could decrease the β-elemene induced apoptosis, while HULC knockdown increased the β-elemene induced apoptosis in DLBCL cells. In vivo study further confirmed the antitumor effect of β-elemene in the DLBCL xenograft, and the HULC expression and the critical proteins of the apoptotic pathway were regulated by β-elemene. These results suggested that HULC-mediated apoptotic pathway was essential for β-elemene to inhibit DLBCL progression.
In summary, our study indicated that the antitumor activity of β-elemene in DLBCL was attributed to the inhibition of cell viability, induction of apoptosis and regulation of HULC-mediated apoptotic pathway. LncRNA HULC may be a potential therapeutic target for β-elemene in treating DLBCL, and β-elemene will be an alternative candidate for clinical application in DLBCL patients. However, further investigations of the deep mechanisms involved in HULC-mediated apoptosis induced by β-elemene are needed.
Abbreviations
- DLBCL
diffuse large B-cell lymphoma
- lncRNA
long non-coding RNA
- NHL
non-Hodgkin lymphoma
- qRT-PCR
quantitative real-time transcription-polymerase chain reaction
Competing Interests
The authors declare that there are no competing interests associated with the manuscript.
Funding
This work was supported by a grant from Zhejiang Provincial Department of Health, General Research Project B [grant number 2015KYB264].
Author Contribution
T.H. and Y.G. conceived the experiments. Y.G. designed the experiments. T.H. and Y.G. performed the experiments. T.H. analyzed the data. T.H. and Y.G. prepared the manuscript. All authors read and approved the final manuscript.
References
- 1.Sehn L.H. and Gascoyne R.D. (2015) Diffuse large B-cell lymphoma: optimizing outcome in the context of clinical and biologic heterogeneity. Blood 125, 22–32 10.1182/blood-2014-05-577189 [DOI] [PubMed] [Google Scholar]
- 2.Martelli M., Ferreri A.J., Agostinelli C., Di Rocco A., Pfreundschuh M. and Pileri S.A. (2013) Diffuse large B-cell lymphoma. Crit. Rev. Oncol. Hematol. 87, 146–171 10.1016/j.critrevonc.2012.12.009 [DOI] [PubMed] [Google Scholar]
- 3.Pasqualucci L. and Dalla-Favera R. (2014) SnapShot: diffuse large B cell lymphoma. Cancer Cell 25, 132.e1–132.e1 [DOI] [PubMed] [Google Scholar]
- 4.Zheng R., Jiang H., Li J., Liu X. and Xu H. (2017) Polyphyllin II restores sensitization of the resistance of PC-9/ZD cells to gefitinib by a negative regulation of the PI3K/Akt/mTOR signaling pathway. Curr. Cancer Drug Targets 17, 376–385 10.2174/1568009616666161213141608 [DOI] [PubMed] [Google Scholar]
- 5.Wang H., Fei Z. and Jiang H. (2017) Polyphyllin VII increases sensitivity to gefitinib by modulating the elevation of P21 in acquired gefitinib resistant non-small cell lung cancer. J. Pharmacol. Sci. 134, 190–196 10.1016/j.jphs.2017.06.005 [DOI] [PubMed] [Google Scholar]
- 6.Jiang H., Zhao P.J., Su D., Feng J. and Ma S.L. (2014) Paris saponin I induces apoptosis via increasing the Bax/Bcl-2 ratio and caspase-3 expression in gefitinib-resistant non-small cell lung cancer in vitro and in vivo. Mol. Med. Rep. 9, 2265–2272 10.3892/mmr.2014.2108 [DOI] [PubMed] [Google Scholar]
- 7.Jiang H., Zhao P., Feng J., Su D. and Ma S. (2014) Effect of Paris saponin I on radiosensitivity in a gefitinib-resistant lung adenocarcinoma cell line. Oncol. Lett. 7, 2059–2064 10.3892/ol.2014.2020 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Hong F., Jiang J., Liu X. and Jiang H. (2019) Anticancer activity of Polyphyllin I in nasopharyngeal carcinoma by modulation of lncRNA ROR and P53 signaling. J. Drug Target. 27, 806–811 10.1080/1061186X.2018.1561887 [DOI] [PubMed] [Google Scholar]
- 9.Zhu X., Jiang H., Li J., Xu J. and Fei Z. (2016) Anticancer Effects of Paris Saponins by Apoptosis and PI3K/AKT Pathway in Gefitinib-Resistant Non-Small Cell Lung Cancer. Med. Sci. Monit. 22, 1435–1441 10.12659/MSM.898558 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Song S., Du L., Jiang H., Zhu X., Li J. and Xu J. (2016) Paris Saponin I Sensitizes Gastric Cancer Cell Lines to Cisplatin via Cell Cycle Arrest and Apoptosis. Med. Sci. Monit. 22, 3798–3803 10.12659/MSM.898232 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Zhao P.J., Song S.C., Du L.W., Zhou G.H., Ma S.L., Li J.H. et al. (2016) Paris Saponins enhance radiosensitivity in a gefitinib-resistant lung adenocarcinoma cell line by inducing apoptosis and G2/M cell cycle phase arrest. Mol. Med. Rep 13, 2878–2884 10.3892/mmr.2016.4865 [DOI] [PubMed] [Google Scholar]
- 12.Zheng R., Rao Y., Jiang H., Liu X., Zhu X., Li J. et al. (2016) Therapeutic potential of Ginsenoside Rg3 via inhibiting Notch/HES1 pathway in lung cancer cells. Transl. Cancer Res. 5, 464–469 10.21037/tcr.2016.07.17 [DOI] [Google Scholar]
- 13.Jiang Z., Jacob J.A., Loganathachetti D.S., Nainangu P. and Chen B. (2017) β-Elemene: Mechanistic Studies on Cancer Cell Interaction and Its Chemosensitization Effect. Front. Pharmacol. 8, 105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Jiang S., Ling C., Li W., Jiang H., Zhi Q. and Jiang M. (2016) Molecular Mechanisms of Anti-cancer Activities of β-elemene: Targeting Hallmarks of Cancer. Anticancer Agent. Med. Chem. 16, 1426–1434 10.2174/1871520616666160211123424 [DOI] [PubMed] [Google Scholar]
- 15.Zheng C., Cai X., Wu S., Liu Z., Shi Y. and Zhou W. (2014) Enhancing effect of β-elemene emulsion on chemotherapy with harringtonine, aclacinomycin, and Ara-c in treatment of refractory/relapsed acute myeloid leukemia. Pak. J. Med. Sci. 30, 1270–1272 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Dousti F., Shahrisa A., Ansari H., Hajjari M., Tahmasebi Birgani Y., Mohammadiasl J. et al. (2018) Long non-coding RNAs expression levels in diffuse large B-cell lymphoma: An in silico analysis. Pathol. Res. Pract. 214, 1462–1466 10.1016/j.prp.2018.08.006 [DOI] [PubMed] [Google Scholar]
- 17.Gao H.Y., Wu B., Yan W., Gong Z.M., Sun Q., Wang H.H. et al. (2017) Microarray expression profiles of long non-coding RNAs in germinal center-like diffuse large B-cell lymphoma. Oncol. Rep. 38, 1363–1372 10.3892/or.2017.5821 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Zhu D., Fang C., Li X., Geng Y., Li R., Wu C. et al. (2017) Predictive analysis of long non-coding RNA expression profiles in diffuse large B-cell lymphoma. Oncotarget 8, 23228–23236 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Xin X., Wu M., Meng Q., Wang C., Lu Y., Yang Y. et al. (2018) Long noncoding RNA HULC accelerates liver cancer by inhibiting PTEN via autophagy cooperation to miR15a. Mol. Cancer 17, 94 10.1186/s12943-018-0843-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Zheng P., Li H., Xu P., Wang X., Shi Z., Han Q. et al. (2018) High lncRNA HULC expression is associated with poor prognosis and promotes tumor progression by regulating epithelial-mesenchymal transition in prostate cancer. Arch. Med. Sci. 14, 679–686 10.5114/aoms.2017.69147 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Chen S., Wu D.D., Sang X.B., Wang L.L., Zong Z.H., Sun K.X. et al. (2017) The lncRNA HULC functions as an oncogene by targeting ATG7 and ITGB1 in epithelial ovarian carcinoma. Cell Death. Dis. 8, e3118 10.1038/cddis.2017.486 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Wang J., Ma W. and Liu Y. (2017) Long non-coding RNA HULC promotes bladder cancer cells proliferation but inhibits apoptosis via regulation of ZIC2 and PI3K/AKT signaling pathway. Cancer Biomark. 20, 425–434 10.3233/CBM-170188 [DOI] [PubMed] [Google Scholar]
- 23.Kong D. and Wang Y. (2018) Knockdown of lncRNA HULC inhibits proliferation, migration, invasion, and promotes apoptosis by sponging miR-122 in osteosarcoma. J. Cell. Biochem. 119, 1050–1061 10.1002/jcb.26273 [DOI] [PubMed] [Google Scholar]
- 24.Zhang Y., Song X., Wang X., Hu J. and Jiang L. (2016) Silencing of LncRNA HULC Enhances Chemotherapy Induced Apoptosis in Human Gastric Cancer. J. Med. Biochem. 35, 137–143 10.1515/jomb-2015-0016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Lu Y., Li Y., Chai X., Kang Q., Zhao P., Xiong J. et al. (2017) Long noncoding RNA HULC promotes cell proliferation by regulating PI3K/AKT signaling pathway in chronic myeloid leukemia. Gene 607, 41–46 10.1016/j.gene.2017.01.004 [DOI] [PubMed] [Google Scholar]
- 26.Chen X., Lun L., Hou H., Tian R., Zhang H. and Zhang Y. (2017) The Value of lncRNA HULC as a Prognostic Factor for Survival of Cancer Outcome: A Meta-Analysis. Cell. Physiol. Biochem. 41, 1424–1434 10.1159/000468005 [DOI] [PubMed] [Google Scholar]
- 27.Peng W., Wu J. and Feng J. (2016) Long noncoding RNA HULC predicts poor clinical outcome and represents pro-oncogenic activity in diffuse large B-cell lymphoma. Biomed. Pharmacother. 79, 188–193 10.1016/j.biopha.2016.02.032 [DOI] [PubMed] [Google Scholar]
- 28.Kong Y., Chen G., Xu Z., Yang G., Li B., Wu X. et al. (2016) Pterostilbene induces apoptosis and cell cycle arrest in diffuse large B-cell lymphoma cells. Sci. Rep. 6, 37417 10.1038/srep37417 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Li L.J., Chai Y., Guo X.J., Chu S.L. and Zhang L.S. (2017) The effects of the long non-coding RNA MALAT-1 regulated autophagy-related signaling pathway on chemotherapy resistance in diffuse large B-cell lymphoma. Biomed. Pharmacother. 89, 939–948 10.1016/j.biopha.2017.02.011 [DOI] [PubMed] [Google Scholar]
- 30.Wang Y., Chen F., Zhao M., Yang Z., Li J., Zhang S. et al. (2017) The long noncoding RNA HULC promotes liver cancer by increasing the expression of the HMGA2 oncogene via sequestration of the microRNA-186. J. Biol. Chem. 292, 15395–15407 10.1074/jbc.M117.783738 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Li D., Liu X., Zhou J., Hu J., Zhang D., Liu J. et al. (2017) Long noncoding RNA HULC modulates the phosphorylation of YB-1 through serving as a scaffold of extracellular signal-regulated kinase and YB-1 to enhance hepatocarcinogenesis. Hepatology 65, 1612–1627 10.1002/hep.29010 [DOI] [PubMed] [Google Scholar]
- 32.Jiang X.R., Guo N., Li X.Q., Yang H.Y., Wang K., Zhang C.L. et al. (2018) Long non-coding RNA HULC promotes proliferation and osteogenic differentiation of bone mesenchymal stem cells via down-regulation of miR-195. Eur. Rev. Med. Pharmacol. Sci. 22, 2954–2965 [DOI] [PubMed] [Google Scholar]
- 33.Chen C., Wang K., Wang Q. and Wang X. (2018) LncRNA HULC mediates radioresistance via autophagy in prostate cancer cells. Braz. J. Med. Biol. Res. 51, e7080 10.1590/1414-431x20187080 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Zhu Y., Zhang X., Qi L., Cai Y., Yang P., Xuan G. et al. (2016) HULC long noncoding RNA silencing suppresses angiogenesis by regulating ESM-1 via the PI3K/Akt/mTOR signaling pathway in human gliomas. Oncotarget 7, 14429–14440 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Fang Z., Chen W., Yuan Z., Liu X. and Jiang H. (2018) LncRNA-MALAT1 contributes to the cisplatin-resistance of lung cancer by upregulating MRP1 and MDR1 via STAT3 activation. Biomed. Pharmacother. 101, 536–542 10.1016/j.biopha.2018.02.130 [DOI] [PubMed] [Google Scholar]
- 36.Yang Q., Chen W.Y., Xu Y.F., Lv X.D., Zhang M. and Jiang H. (2018) Polyphyllin I modulates MALAT1/STAT3 signaling to induce apoptosis in gefitinib-resistant non-small cell lung cancer. Toxicol. Appl. Pharmacol. 356, 1–7 10.1016/j.taap.2018.07.031 [DOI] [PubMed] [Google Scholar]
- 37.Xiang S., Zou P., Wu J., Zheng F., Tang Q., Zhou J. et al. (2018) Crosstalk of NF-κB/P65 and LncRNA HOTAIR-Mediated Repression of MUC1 Expression Contribute to Synergistic Inhibition of Castration-Resistant Prostate Cancer by Polyphyllin 1-Enzalutamide Combination Treatment. Cell. Physiol. Biochem. 47, 759–773 10.1159/000490028 [DOI] [PubMed] [Google Scholar]
- 38.Hu Z., Wu H., Li Y., Hou Q., Wang Y., Li S. et al. (2015) β-Elemene inhibits the proliferation of esophageal squamous cell carcinoma by regulating long noncoding RNA-mediated inhibition of hTERT expression. Anticancer Drugs 26, 531–539 10.1097/CAD.0000000000000216 [DOI] [PubMed] [Google Scholar]