
Figure 3: Slowing peptidoglycan synthesis compensates for loss of LytH.
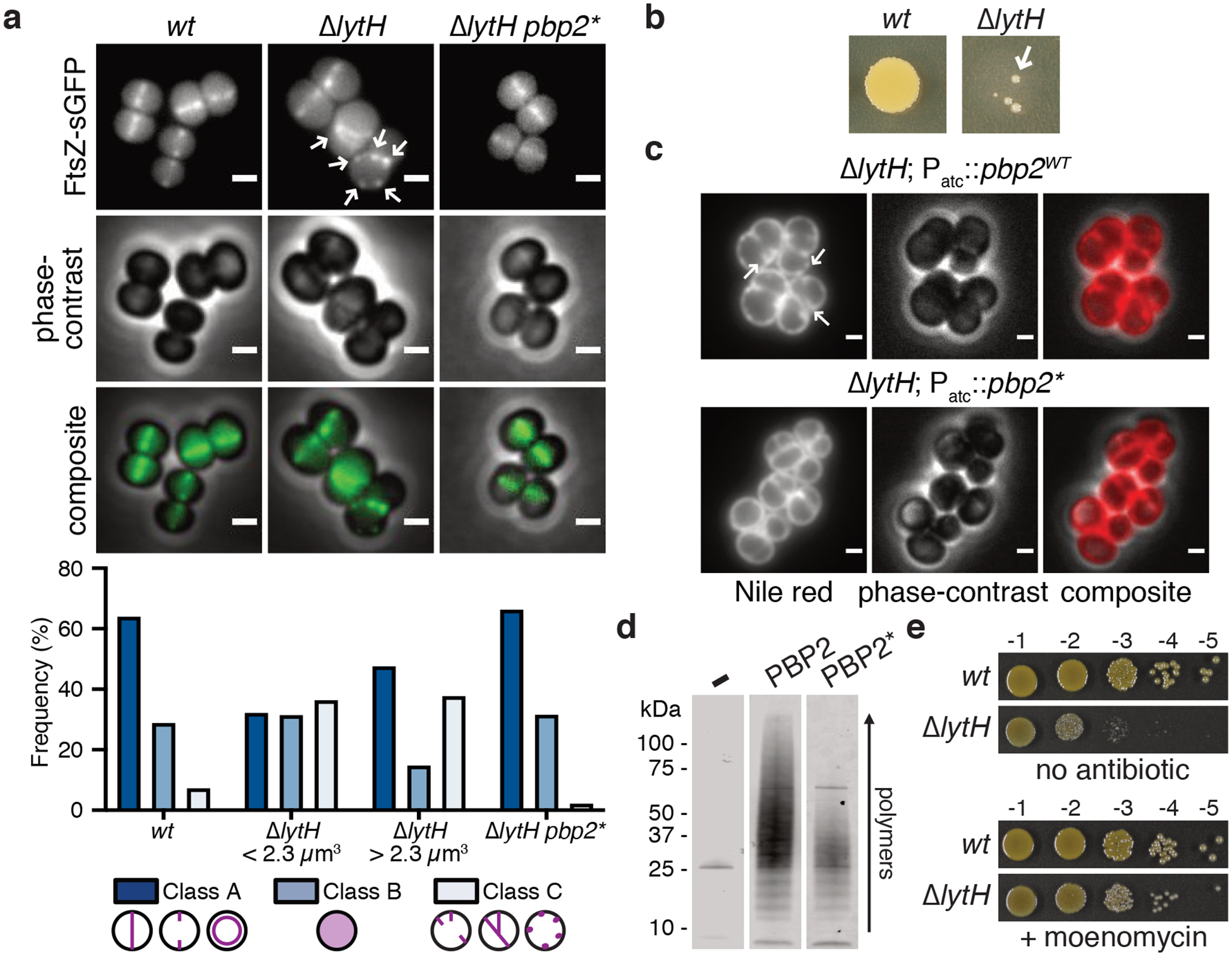
a, FtsZ was frequently mislocalized in the absence of LytH (arrows). Quantitation of FtsZ-sGFP localization either at midcell where a nascent or complete septum was formed (Class A), as diffuse fluorescence (Class B), or as peripheral puncta/multiple septa (Class C). ΔlytH cells were divided into two categories – cells that are smaller or larger than the median cell volume of 2.3 μm14 – which were separately analyzed. The suppressor allele pbp2F158L (pbp2*) corrected ΔlytH defects. From left to right, n = 965, 550, 549, and 1963 cells. Scale bars, 1 μm. b, High-temperature (42°C) sensitivity of lytH deletion was suppressed (arrow) by reducing PBP2 activity. c, Overexpression of pbp2WT from a plasmid in a lytH deletion background exacerbated cell division defects. Overexpression of a suppressor allele, pbp2N220→KDLN (pbp2*), was not toxic. In these strains, the endogenous copy of pbp2WT was still intact. Cells were stained with the membrane dye Nile red. Scale bars, 1 μm. d, Purified PBP2 and a representative variant, PBP2F158L (PBP2*), were incubated with Lipid II to assess polymerization activity. e, Reducing peptidoglycan synthesis activity with a compound suppressed ΔlytH high-temperature sensitivity. Moenomycin inhibits PBP2. Data are representative of two (a-e) independent experiments.