
Figure 14.
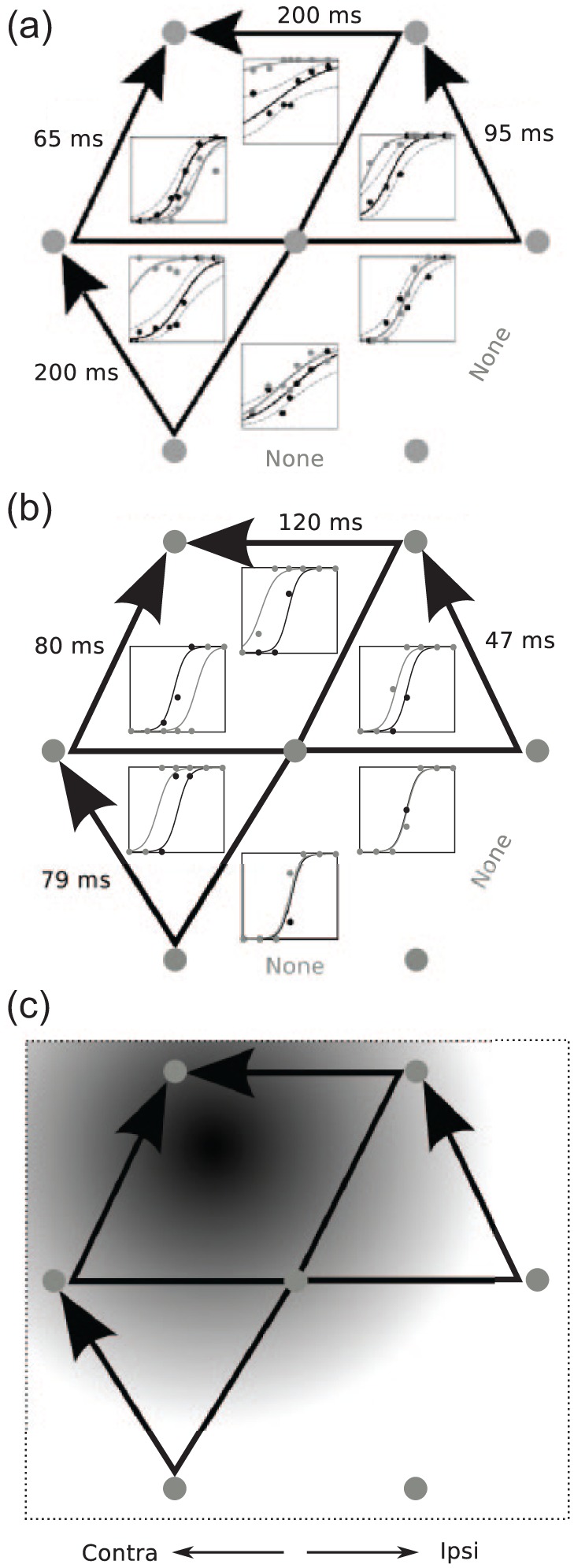
SEF microstimulation causes saccade trajectories to converge. Each arrow denotes the initial and final position of a saccadic eye movement during a memory-based saccadic task. In this task, Histed and Miller (2006) trained monkeys to perform a task in which two spatial positions were sequentially cued during an initial fixation phase, remembered during a memory delay, and then visited in order with a sequence of saccades following the offset of the fixation point. Microstimulation did not disrupt the monkey’s ability to correctly saccade to the remembered target positions, thereby supporting the hypothesis that a spatial working memory encodes the saccadic target positions. However, microstimulation did disrupt the order of saccades. The bias of these movements to converge to a position at the upper left target position illustrates the effect of microstimulation. (a) Observed saccade trajectories that converge toward the upper left target. (b) Model simulations reproduce the convergence effect. (c) In model simulations, microstimulation habituates synapses according to a two-dimensional Gaussian function centred over the microstimulation site. The depth of habituation diminishes in a Gaussian fashion across space from black (maximal) to white (minimum). Saccade trajectories after following microstimulation tend to ‘climb’ the Gaussian habituation gradient. Saccades that are furthest from the microstimulation site are least affected by it and thus most likely to serve as the first saccade target position. Data adapted from Histed and Miller (2006). Another important finding of Histed and Miller (2006) was that microstimulation had no effect on saccade accuracy, peak velocity, or latency. These observations suggest that SEF is not involved in the storage, generation, or fine timing of saccades. A target selection role is consistent with these data because SEF microstimulation manipulates only the potency with which plans compete and, during fixation while basal ganglia SNr gates are closed, do not generate new saccade targets. Moreover, if SEF selects targets but does not issue the motor commands that move the eyes, saccade velocity remains unchanged. So long as the duration of the selection process is not changed by microstimulation, saccade latency will also remain unchanged. Reprinted with permission from Silver et al. (2011).