
Fig. 6. Translocation over an mRNA hairpin encountered at the distal active site.
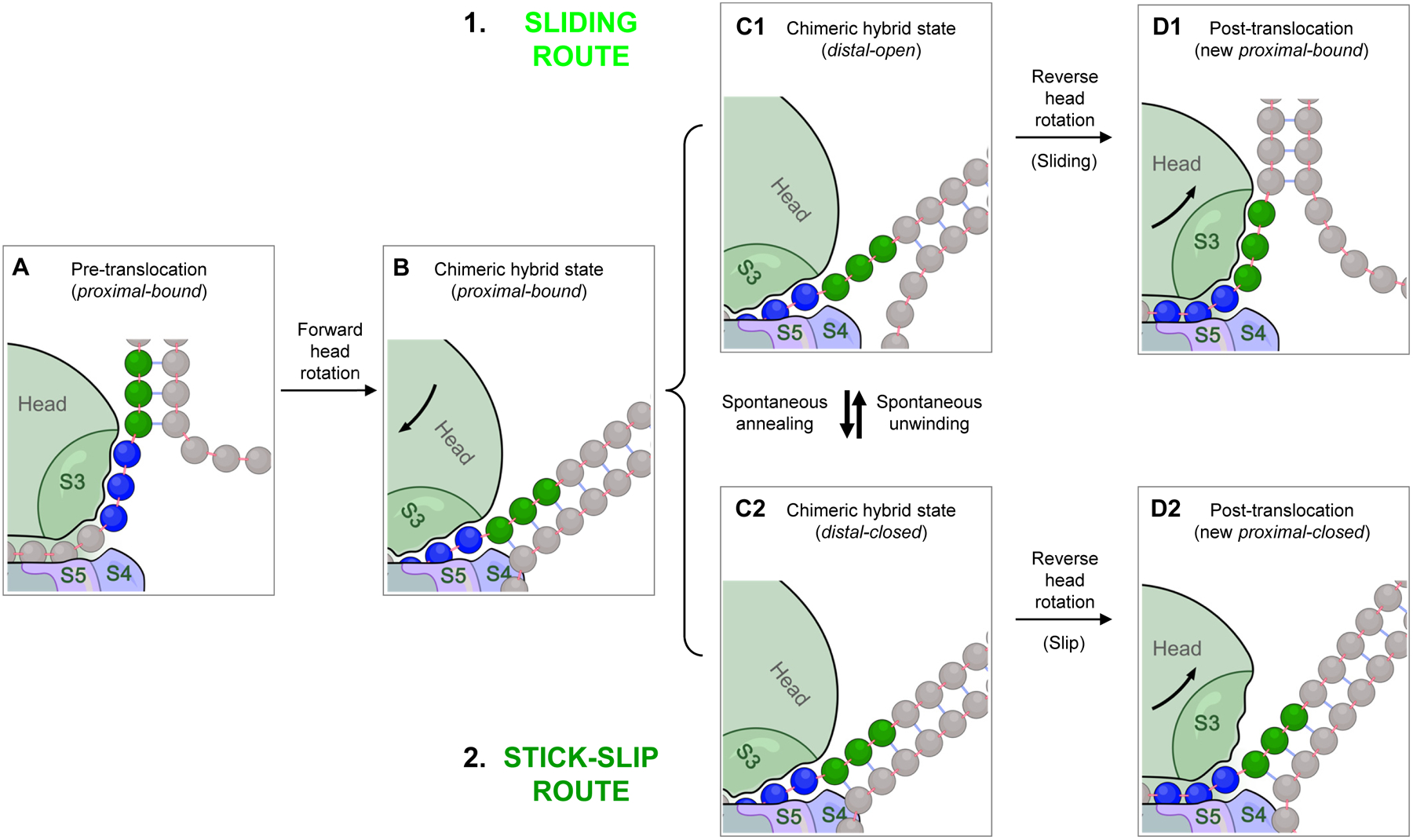
A. Before translocation starts, the proximal mRNA segment (blue) must be in the bound state. B. Forward head rotation moves S3 (part of the head domain) along with its bound mRNA as a rigid body relative to the body domain, with no unwinding. C1 and C2. The head-rotated intermediate in panel B is in fact composed of two different states of the distal segment (green) in equilibrium: (C1) the distal-open state, and (C2) the distal-closed state. The same equilibrium exists in the pre-translocation state (panel A), but not shown for simplicity. D1 and D2. During reverse head rotation, the head domain moves back to its classical state, whereas the mRNA remains in place, forcing S3 to move relative to the mRNA. If the distal segment is open, S3 can slide along the mRNA, resulting in a post-translocation state in which the now-proximal segment is in the bound state (D1). If the distal segment is closed, S3 loses its bound mRNA without acquiring compensatory binding; i.e. stick-slip occurs, resulting in a proximal-closed post-translocation state (D2). The latter has to spontaneously convert to proximal-bound state before the next translocation step can take place. Note that the distal-open state (panel C1) can also undergo stick-slip to yield a proximal-open post-translocation product (not shown; see Fig. 5), but this occurs relatively rarely for this state, and omitted here for simplicity.