
Fig. 1.
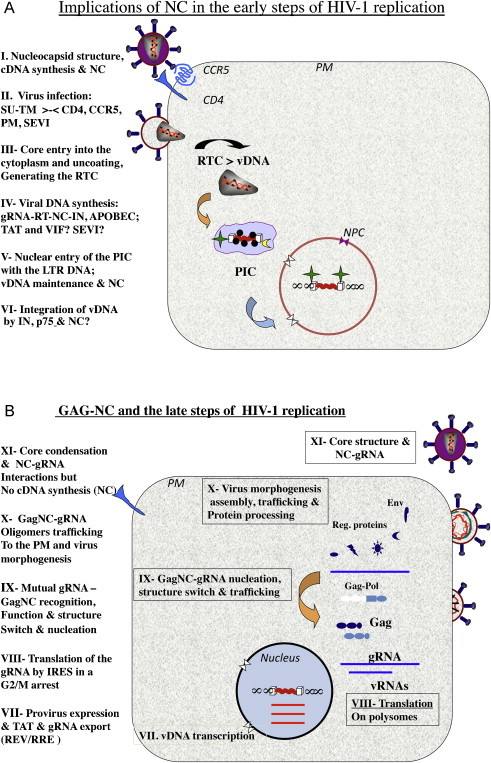
Schematic representation of HIV-1 replication. (a) The early steps of HIV-1 replication. The implications of mature NC protein (NCp7) are at the levels of (I) the structure of a stable, condensed nucleo-core where 1500–2000 NC molecules coat the dimeric RNA genome of 60S, providing protection and negatively regulating reverse transcription; (II) virus infection is mediated by molecular recognition and interactions between the virus trimeric SUgp120-TMgp41 proteins and the cellular CD4 receptor and CCR5 coreceptor at the plasma membrane (PM), that are believed to be augmented in vivo by peptides present in the semen (SEVI; see Section 4.1). Next the viral core (III) is delivered into the cytoplasm where two dynamic processes take place, namely partial core uncoating and viral DNA (vDNA) synthesis. (IV) Viral DNA synthesis by the viral RT enzyme, chaperoned by NC molecules from initiation to completion; this slow process (6–12 h in cells effectively targeted by HIV-1 in vivo (see Section 4.1)) is believed to be facilitated by the viral VIF and TAT proteins and by the cellular SEVI that are all nucleic acid chaperones. In the absence of VIF expression in virus producer cells, the restriction factor APOBEC 3G can be incorporated into the newly made viral particles by interacting with NC, which in turn causes deamination of C residues in nescent cDNA. (V) Once the viral DNA flanked by the LTRs is made it is found in a poorly defined molecular complex called the preintegration complex (PIC) (Arhel, 2010). Integrity of the vDNA is essential since the LTR ends called inverted repeats (ir) are required for the IN-mediated integration of the vDNA into the host genome. Protection of the LTR ends against cellular exonucleases such as TREX-1 (Yan et al., 2010) is thought to be mediated by the binding of molecules of NC and IN. In agreement with this notion, mutating the highly conserved CCHC zinc fingers results in partial trimming of the LTR DNA ends (Buckman et al., 2003, Tanchou et al., 1998), which impairs vDNA integration. (VI) Once the PIC is actively delivered into the nucleus via the NPC (nucleopore complex), the tetrameric IN enzyme in a complex with the cellular helper factor LEDGF/p75 mediates vDNA integration into the host genome. This reaction is believed to be chaperoned by NC but this is still a matter of debate. Also once in the nucleus and before its integration the vDNA can be expressed to produce the viral factors TAT, REV and NEF that are essential for the late steps of HIV-1 replication and the immune response (Wu and Marsh, 2001). (b) Schematic representation of the late steps of HIV-1 replication. (VII) The integrated proviral DNA is transcribed by the host cell RNA polymerase B machinery, with the assistance of the viral trans-acting factor TAT at the levels of initiation and elongation (Wu, 2004). The newly made full length (FL) vRNA is either exported from the nucleus which requires the viral protein REV bound to the RRE or spliced to generate the single and multiple spliced subgenomic RNAs. (VIII) The Gag and GagPol polyprotein precursors to the structural proteins and enzymes, respectively, of the virus are synthesized by the host cell translation machinery using the FL RNA as the messenger RNA template. This is carried out by an original mechanism whereby the host ribosomes directly bind to sequences within the multifunctional 5′ leader, involved in RNA synthesis, splicing and export, and translation, dimerization and packaging, and vDNA synthesis. Such a small domain is called internal ribosome entry site (IRES), which ensures a high level of viral translation throughout the cell cyle, notably in the G2M arrest caused by the viral protein VPR (Balvay et al., 2007). (IX) As soon as Gag accumulates in the cytosol several Gag molecules recognize and bind to the 5′ Packaging region of the FL viral RNA (such as SL2; see Fig. 2d). This in turn would render the 5′ leader inaccessible to translating ribosomes due to structural rearrangements (see text) together with the presence of bound Gag molecules (Huthoff and Berkhout, 2001a, Huthoff and Berkhout, 2001b, Huthoff and Berkhout, 2002). Thus the specific interactions between the NC domain of Gag, notably the ZFs, and the Psi packaging signal would cause a functional switch from translation to virus assembly, representing the nucleation event where the FL RNA acts as the initial platform or scaffold for Gag assembly. (X) Further to the nucleation event, another molecular switch takes place at the opposite end of Gag where the N-terminal myristate becomes accessible, and addresses the Gag–RNP complexes to cellular membranes to pursue and complete virus assembly via interactions with, and recruitment of viral and cellular proteins and NAs (Cimarelli and Darlix, 2002, Freed and Mouland, 2006, Hamard-Peron and Muriaux, 2011) and references therein. (XI) As virion formation progresses, GagPol and Gag become oligomeric which in turn activates the viral protease (PR). Subsequently the viral PR processes Gag and GagPol in a sophisticated manner, first giving rise to MAp17 and NCp15, and RT. Ultimately, upon and/or during virus budding the fully mature Gag proteins are generated, notably NCp7, leading to core condensation while the dimeric RNA genome becomes a mature 60S competent for reverse transcription.