

Abstract
Feline coronavirus (FCoV) is divided into two types I and II, based on their growth in vitro and antigenicity. In this study, virus neutralization (VN) test was applied for type differentiation of FCoV infections. Sera of cats which were clinically and serologically diagnosed as feline infectious peritonitis (FIP) possessed significantly higher VN titers to type I FCoV, and sera from cats experimentally infected with FIPV type II had high VN titers to type II but not type I viruses. A total of 79 cat sera collected in the years between 2004 and 2005 were examined to evaluate seroprevalence by the VN test, showing the following results: (1) 50 cats (63.3%) were sero-positive to FCoV; (2) of the 50 FCoV positive cat serum samples, 49 (98%) showed significantly higher titers to type I virus and only one (2%) for type II virus. These results indicate that the VN test described here can be used for serological differentiation of FCoV infections of cats, and that FCoV type I is a dominant type in recent years of Japan.
Keywords: Feline coronavirus, Feline infectious peritonitis virus, Virus neutralization test
1. Introduction
Feline infectious peritonitis (FIP) is a highly fatal and immune-mediated pyogranulomatous disease that occurs in both domestic and wild felidae. The disease is caused by feline coronavirus (FCoV). FCoV is a member of the family Coronaviridae, genus Coronavirus which is a group of enveloped and positive-stranded RNA viruses. Antigenically and genetically, coronaviruses are currently grouped into at least four groups. FCoV belongs to group I coronavirus which includes canine coronavirus (CCoV), transmissible gastroenteritis virus (TGEV), porcine respiratory coronavirus and human coronavirus 229E (Wege et al., 1982).
Two pathotypes of FCoVs are also known: one is feline enteric coronavirus (FECV) which causes from asymptomatic infection to severe enteritis and another is feline infectious peritonitis virus (FIPV) that causes fatal immune-mediated disease FIP (Pedersen, 1987, Pedersen et al., 1981). It was demonstrated that FIPV arises inside of cats by mutation from infected FECV (Vennema et al., 1998). However, it is difficult to distinguish between FECV and FIPV by in vitro serological and genetical methods (Boyle et al., 1984, Fiscus and Teramoto, 1987, Pedersen et al., 1984).
Both FECV and FIPV are divided into two types I and II, based on their neutralization reactivity with spike (S) protein-specific mAbs (Fiscus and Teramoto, 1987, Hohdatsu et al., 1991b, Hohdatsu et al., 1992) and sequence analysis of the S protein gene (Motokawa et al., 1995, Motokawa et al., 1996). In addition to this serological property, some other biological characteristics to each type have been described. While FCoV type I grows poorly in cell culture, type II can grow well in many different cell lines (Pedersen et al., 1984). Furthermore, type II FCoV shows close antigenic and genetic relationship to CCoV and TGEV, suggesting that FCoV type II arose from double recombination between FCoV type I and CCoV (Herrewegh et al., 1998)
FCoV type II strains are more frequently utilized than type I strains for in vitro experiments because of higher efficient growth. For serological survey of FCoV infection, antibody has been detected by indirect fluorescent antibody assay (IFA) or enzyme-linked immunosorbent assay (ELISA) using FCoV (Pedersen, 1976, Scott, 1979, Horzinek and Osterhaus, 1979, Ishida et al., 1987). Alternatively CCoV or TGEV are also used as a target antigen because of serological close relations to FCoV. However, discrimination ability (specificity) of these methods using heterologous antigens seems to be low since they detect also cross-reactive antibodies between types I and II, such as antibody to nucleocapsid (N) protein.
In this study, we developed a plaque-reduction neutralization test (PRNT) to serologically distinguish FCoV type I and II infections in cats, and we applied it for field cases.
2. Materials and methods
2.1. Cell cultures
Feline whole fetus cells (fcwf-4 cells), which are fetal feline lung cells having characteristics of macrophages (Jacobse-Geels and Horzinek, 1983) were grown in Dulbecco's modified Eagle's medium (DMEM) containing 10% fetal calf serum (FCS), 100 U/ml penicillin and 100 μg/ml streptomycin. The cells were maintained in a humidified 5% CO2 incubator at 37 °C.
2.2. Viruses
FIPV type I strains C3663 and Yayoi and FIPV type II strains KUK-H/L and M91-267 were used in this study. KUK-H strain was isolated from the spleen sample of effusive form FIP case in 1987 by using CRFK cells, and KUK-H/L was plaque-purified from the KUK-H strain (Mochizuki et al., 1997). M91-267 strain was isolated from the spleen sample taken at the postmortem examination of effusive form FIP case in 1991(Mochizuki et al., 1997). These two strains were classified into type II by using FCoV type-specific mAbs, kindly provided by Dr. T. Hohdatsu (Hohdatsu et al., 1991a, Hohdatsu et al., 1991b). C3663 strain was isolated from a cat with an effusive form of FIP in 1994 and used as a reference FCoV type I (Mochizuki et al., 1997). In addition, it was genetically confirmed that C3663 belongs to type I FCoV and M91-267 and KUK-H/L do to type II (manuscript in preparation). Yayoi strain was isolated from liver homogenate from non-effusive form of FIP by serial passage in suckling mouse brain and then in fcwf-4 cells and have been used as Japanese prototype strain of FCoV type I (Hayashi et al., 1981). C3663, KUK-H/L and M91-267 strains were used in subsequent experiments within 10 passages in fcwf-4 cells. Although the passage history of Yayoi strain is unknown, it was propagated less than three times in our laboratory.
2.3. Cat sera
Four sera (Nos.1–4) were collected from cats clinically diagnosed as FIP and serologically identified as type I infection by indirect immunofluorescent assay using unfixed fcwf-4 cells infected with FCoV type I or II. On the other hand, we could not obtain any sera from naturally FCoV type II infected cats. Therefore, two sera collected from cats experimentally infected with M91-267 or KUK-H/L were used in this study (Mochizuki et al., 1997).
To evaluate the seroprevalence of FCoV types I and II in Japan, sera were collected from 79 cats, which visited to one veterinary hospital in 2004 and 2005. Two of them were clinically and serologically diagnosed as FIP in the hospital.
2.4. Plaque-reduction neutralization test (PRNT)
Serological test was performed by PRNT using serum samples inactivated by heating at 56 °C for 30 min. The sera diluted with DMEM containing 2% FCS and each virus diluted to approximately 2 × 103 PFU/ml with DMEM containing 2% FCS were equally mixed and incubated at 37 °C for 1 h. Then 50 μl of the mixture was inoculated onto fcwf-4 cells monolayer in 24-well plate (SUMITOMO BAKELITE, Japan). After absorption at 37 °C 1 h, the inoculum was removed and 0.8% agarose (SeaPlaque® Agarose, TaKaRa, Japan) in growth medium was overlaid. The infected cells were incubated at 37 °C until cytopathic effect (CPE) appeared, fixed with phosphate-buffered formalin and stained with crystal violet. When the number of plaques was less than 75% of those in control wells, the diluted sera were judged to be positive. For screening of cat sera, all sera finally diluted to 1:10 were examined for PRNT. The positive sera were further diluted to 1:33, 1:100, 1:333, 1:1000, 1:3333, and 1:10000 to determine virus-neutralizing titers. The titers were expressed as the highest serum dilution showing 75% plaque reduction or more.
3. Results
3.1. Development of virus neutralization (VN) test
To determine the antigenic relationship between FCoV types I and II, serum samples from cats experimentally infected with FIPV type II M91-267 or KUK-H/L and naturally infected with FIPV type I were examined by PRNT. Four sera from FIPV type I infected cats showed VN titers to C3663 and Yayoi ranging from 1:1000 to 1:16000, and those to M91-267 and KUK-H/L ranging from <1:10 to 1:40 (Table 1 ). Sera collected from the experimentally FIPV type II infected cats had higher VN titers to type II ranging from 1:4000 to 1:128000 and lower VN titers to type I ranging from 1:80 to 1:1280 (Table 1). These results indicated that FCoV-infected cat sera had significantly higher VN titers to homologous type, but not to heterologous type. Therefore, this PRNT using both types of FCoV must be able to distinguish serologically between FCoV type I and II infections.
Table 1.
VN titers to FCoV types I and II in FIPV-infected cats
| FCoV type | Strain | Sera to FCoV type Ia |
Sera to FCoV type II |
||||
|---|---|---|---|---|---|---|---|
| No. 1 | No. 2 | No. 3 | No. 4 | No. 5b | No. 6c | ||
| Type I | C3663 | 4000 | 4000 | 4000 | 4000 | 160 | 1280 |
| Yayoi | 1000 | 16000 | 2000 | 2000 | 80 | 640 | |
| Type II | M91-267 | 40 | <10 | 40 | <10 | 20000 | 128000 |
| KUK-H/L | 10 | <10 | 40 | <10 | 4000 | 128000 | |
VN titers are expressed as the reciprocal value of the highest dilution.
Sera were collected from cats clinically diagnosed as FIP and serologically diagnosed as FIPV type I infection.
Serum was collected from a cat experimentally inoculated with M91-267.
Serum was collected from a cat experimentally inoculated with KUK-H/L.
3.2. Prevalence of FCoV types in Japan
A total of 79 cat sera collected in Japan were examined for the prevalence of antibodies to FCoV types I and II by PRNT using C3663 and M91-267 strains. The result showed that 50 (63.3%) cats were seropositive to FCoV(Fig. 1 ). Among these seropositive samples, 49 (98%) possessed significantly higher titers against type I, and only one (2%) did against type II. Although three samples had VN titers of 1:33 to type II, two of them had higher VN titers to type I (1:3333). Since one sample did not have VN titer to type I, it was regarded as positive for type II. All two samples from FIP cases had the highest VN titers to type I (1:10000), meanwhile, these were negative (<1:10) to type II.
Fig. 1.
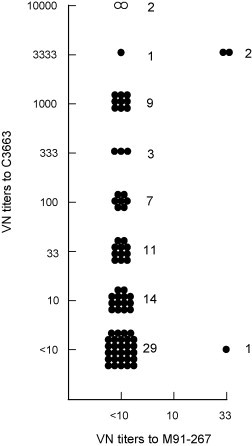
Distribution of virus-neutralizing titers to FIPV types I and II among cats. VN titers are expressed as the reciprocal value of the highest dilution. Open circles show cats diagnosed as FIP.
4. Discussion
In this study, we distinguished types of FCoV which had infected cats by VN test. At first, we determined cross-reactivities of the known sera from FIP cats (Table 1). VN titers difference between homologous type was five-fold or less, and to heterologous type was 25-fold or more, indicating that the present PRNT can clearly differentiate FCoV types. It is known that a major difference between types I and II is located in S protein responsible for eliciting VN antibodies (Fiscus and Teramoto, 1987, De Groot et al., 1989, Corapi et al., 1992).
Using the PRNT, we examined the recent status of FCoV seroprevalence in field cats. More than 60% of the cats were found to be FCoV seropositive and this was similar to the positive rate described in other countries (Pedersen et al., 1981, Loeffler et al., 1978, Kummrow et al., 2005, Bell et al., 2006). In Japan, Hohdatsu et al. (1992) first described that 21.3% of chronically diseased cats and 13.4% of non-diseased cats had antibodies to FCoV by IFA to detect FCoV infection. The seroprevalence rate obtained in the present study is about three times higher than that in the previous report. Since different methods and samples were used for epidemiology, it is impossible to directly compare the prevalence of FCoV in Japan. However, most probable explanation for this discrepancy is that FCoV has been widespreading among Japanese cat population during the last decade.
Previous studies conducted in several countries including Japan have revealed that 69–91% of sero-positive cats were suspected to be type I infection (Hohdatsu et al., 1992, Vennema, 1999, Kummrow et al., 2005). In the present study, it was found that 49 cats (98%) possessed type I specific antibodies and only one cat (2%) to type II. The prevalence of type I infection revealed here was significantly higher than the previous report (80.4% in chronically diseased cats) by competitive ELISA (Hohdatsu et al., 1992). Although this and the previous report (Hohdatsu et al., 1992) were different in methods and samples, it might suggest that type II FCoV infection has been decreasing in Japanese domestic cats in recent years.
In conclusion, our newly established VN test can specifically detect and distinguish between types of FCoV infection. In addition, FCoV type I, but not type II, has widely spread among cats population in Japan.
Acknowledgements
This study was supported by Grand-in-Aid from the Ministry of Education, Culture, Sports, Science and Technology of Japan and the Research Fund of Kyoritsu Seiyaku Corporation. We would like to thank Dr. T. Hohdatsu for providing mAbs and Drs. K. Hirata, Y. Hirata and M. Iwata for collecting sera.
References
- Bell E.T., Toribio J.A., White J.D., Malik R., Norris J.M. Seroprevalence study of feline coronavirus in owned and feral cats in Sydney, Australia. Aust. Vet. J. 2006;84:74–81. doi: 10.1111/j.1751-0813.2006.tb12231.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boyle J.F., Pedersen N.C., Evermann J.F., McKeirnan A.J., Ott R.L., Black J.W. Plaque assay, polypeptide composition and immunochemistry of feline infectious peritonitis virus and feline enteric coronavirus isolates. Adv. Exp. Med. Biol. 1984;173:133–147. doi: 10.1007/978-1-4615-9373-7_12. [DOI] [PubMed] [Google Scholar]
- Corapi W.V., Olsen C.W., Scott F.W. Monoclonal antibody analysis of neutralization and antibody-dependent enhancement of feline infectious peritonitis virus. J. Virol. 1992;66:6695–6705. doi: 10.1128/jvi.66.11.6695-6705.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Groot R.J., Van Leen R.W., Dalderup M.J.M., Vennema H., Horzinek M.C., Spaan W.J.M. Stably expressed FIPV peplomer protein induces cell fusion and elicits neutralizing antibodies in mice. Virology. 1989;171:493–502. doi: 10.1016/0042-6822(89)90619-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fiscus S.A., Teramoto Y.A. Antigenic comparison of feline coronavirus isolates: evidence for markedly different peplomer glycoproteins. J. Virol. 1987;61:2607–2613. doi: 10.1128/jvi.61.8.2607-2613.1987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hayashi T., Yanai T., Tsurudome M., Nakayama H., Watabe Y., Fujiwara K. Serodiagnosis for feline infectious peritonitis by immunofluorescence using infected suckling mouse brain sections. Jpn. J. Vet. Sci. 1981;43:669–676. doi: 10.1292/jvms1939.43.669. [DOI] [PubMed] [Google Scholar]
- Herrewegh A.A.P.M., Smeenk I., Horzinek M.C., Rottier P.J.M., De Groot R.J. Feline coronavirus type II strains 79-1683 and 79-1146 originate from a double recombination between feline coronavirus type I and canine coronavirus. J. Virol. 1998;72:4508–4514. doi: 10.1128/jvi.72.5.4508-4514.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hohdatsu T., Okada S., Koyama H. Characterization of monoclonal antibodies against feline infectious peritonitis virus type II and antigenic relationship between feline, porcine, and canine coronaviruses. Arch. Virol. 1991;117:85–95. doi: 10.1007/BF01310494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hohdatsu T., Sasamoto T., Okada S., Koyama H. Antigenic analysis of feline coronaviruses with monoclonal antibodies (MAbs): preparation of MAbs which discriminate between FIPV strain 79-1146 and FECV strain 79-1683. Vet. Microbiol. 1991;28:13–24. doi: 10.1016/0378-1135(91)90096-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hohdatsu T., Okada S., Ishizuka Y., Yamada H., Koyama H. The prevalence of types I and II feline coronavirus infections in cats. J. Vet. Med. Sci. 1992;54:557–562. doi: 10.1292/jvms.54.557. [DOI] [PubMed] [Google Scholar]
- Horzinek M.C., Osterhaus A.D.M.E. Feline infectious peritonitis: a worldwide serosurvey. Am. J. Res. 1979;40(10):1487–1492. [PubMed] [Google Scholar]
- Ishida T., Washizu T., Fukuoka J., Toriyabe K., Uchino T., Motoyoshi S. Feline infectious peritonitis virus antibody test using enzyme-linked immunosorbent assay. Jpn. J. Vet. Sci. 1987;49(1):145–149. doi: 10.1292/jvms1939.49.145. [DOI] [PubMed] [Google Scholar]
- Jacobse-Geels H.E.L., Horzinek M.C. Expression of feline infectious peritonitis coronavirus antigens on the surface of feline macrophage-like cells. J. Gen. Virol. 1983;64:1859–1866. doi: 10.1099/0022-1317-64-9-1859. [DOI] [PubMed] [Google Scholar]
- Kummrow M., Meli M.L., Haessig M., Goenczi E., Poland A., Pedersen N.C., Hofmann-Lehmann R., Lutz H. Feline coronavirus serotypes 1 and 2: seroprevalence and association with disease in Switzerland. Clin. Diagn. Lab. Immunol. 2005;12:1209–1215. doi: 10.1128/CDLI.12.10.1209-1215.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Loeffler D.G., Ott R.L., Evermann J.F., Alexander J.E. The incidence of naturally occurring antibodies against feline infectious peritonitis in selected cat populations. Feline Pract. 1978;8:43–45. [Google Scholar]
- Mochizuki M., Mitsutake Y., Miyanohara Y., Higashihara T., Shimizu T., Hohdatsu T. Antigenic and plaque variations of serotype II feline infectious peritonitis coronaviruses. J. Vet. Med. Sci. 1997;59(4):253–258. doi: 10.1292/jvms.59.253. [DOI] [PubMed] [Google Scholar]
- Motokawa K., Hohdatsu T., Aizawa C., Koyama H., Hashimoto H. Molecular cloning and sequence determination of the peplomer protein gene of feline infectious peritonitis virus type I. Arch. Virol. 1995;140:469–480. doi: 10.1007/BF01718424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Motokawa K., Hohdatsu T., Hashimoto H., Koyama H. Comparison of the amino acid sequence and phylogenetic analysis of the peplomer, integral membrane and nucleocapsid proteins of feline, canine and porcine coronaviruses. Microbiol. Immunol. 1996;40:425–433. doi: 10.1111/j.1348-0421.1996.tb01089.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pedersen N.C. Serologic studies of naturally occurring feline infectious peritonitis. Am. J. Vet. Res. 1976;37(12):1449–1453. [PubMed] [Google Scholar]
- Pedersen N.C. Virologic and immunologic aspects of feline infectious peritonitis virus infection. Adv. Exp. Med. Biol. 1987;218:529–550. doi: 10.1007/978-1-4684-1280-2_69. [DOI] [PubMed] [Google Scholar]
- Pedersen N.C., Boyle J.F., Floyd K., Fudge A., Barker J. An enteric coronavirus infection of cats and its relationship to feline infectious peritonitis. Am. J. Vet. Res. 1981;42:368–377. [PubMed] [Google Scholar]
- Pedersen N.C., Black J.W., Boyle J.F., Evermann J.F., McKeirnan A.J., Ott R.L. Pathogenic differences between various feline coronavirus isolates. Adv. Exp. Med. Biol. 1984;173:365–380. doi: 10.1007/978-1-4615-9373-7_36. [DOI] [PubMed] [Google Scholar]
- Scott F.W. FIP antibody test—interpretation and recommendations. J. Am. Vet. Med. Assoc. 1979;175:1164–1168. [PubMed] [Google Scholar]
- Vennema H., Poland A., Foley J., Pedersen N.C. Feline infectious peritonitis viruses arise by mutation from endemic feline enteric coronaviruses. Virology. 1998;243:150–157. doi: 10.1006/viro.1998.9045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vennema H. Genetic drift and genetic shift during feline coronavirus evolution. Vet. Microbiol. 1999;69:139–141. doi: 10.1016/S0378-1135(99)00102-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wege H., Siddell S., Ter Meulen V. The biology and pathogenesis of coronaviruses. Curr. Top. Microbiol. Immunol. 1982;99:165–200. doi: 10.1007/978-3-642-68528-6_5. [DOI] [PubMed] [Google Scholar]