

Abstract
Little is known about the ecology of many of the parasites and pathogens affecting bats, but host social behavior almost certainly plays an important role in bat-parasite dynamics. Understanding parasite dynamics for bats is important from a human public health perspective because of their role as natural reservoirs for recent high-profile emerging zoonotic pathogens (e.g. Ebola, Hendra) and from a bat conservation perspective because of the recent emergence of white-nose syndrome (WNS) in North America highlighting the potential population impacts of parasites and pathogens. Although some bat species are among the most gregarious of mammals, species vary widely in terms of their social behavior and this variation could influence pathogen transmission and impacts. Here, we review the literature on links between bat social behavior and parasite dynamics. Using standardized search terms in Web of Science, we identified articles that explicitly tested or discussed links between some aspect of bat sociality and parasite transmission or host population impacts. We identified social network analysis, epidemiological modeling, and interspecific comparative analyses as the most commonly used methods to quantify relationships between social behavior and parasite-risk in bats while WNS, Hendra virus, and arthropod ectoparasites were the most commonly studied host-parasite systems. We summarize known host-parasite relationships in these three systems and propose testable hypotheses that could improve our understanding of links between host sociality and parasite-dynamics in bats.
Keywords: Colony Size, Social Network Analysis, Fusion Dynamic, Parasite Species Richness, Hendra Virus
Introduction
Parasitism is ubiquitous in nature. Parasites affect fitness of their hosts and thus can shape host population dynamics. Defined broadly, a parasite is any organism that grows, feeds, and/or is sheltered at the expense of another organism (i.e. the host). Parasites can be categorized into different ways but one common approach is to differentiate microparasites, which are typically unicellular or multicellular microbes (e.g. bacteria, prions, viruses, protozoans, fungi) with short generation times and a life cycle that occurs entirely on/in the host, from macroparasites which are multicellular parasites with longer generation times and more complex life cycles that may include multiple host species (Table 5.1; Anderson and May 1979; Hudson et al. 2002). Many microparasites can be pathogenic and cause identifiable disease in their hosts with recognizable physiological or behavioral signs or symptoms (Table 5.1). For example, Batrachochytrium dendrobatiidis is a fungal microparasite of amphibians that infects keratinized tissues, thickens the epidermis, and eventually causes mortality, all of which are identifiable signs of the associated disease, chytridiomycosis (Voyles et al. 2009; Rosenblum et al. 2010). Macroparasites tend to result in chronic infections of their definitive hosts, decrease host fecundity, and usually cause morbidity rather than mortality (Hudson et al. 2002). Experimental infection of great tits (Parus major) with the hen flea (Ceratophyllus gallinae) resulted in reduced reproductive success via increased nest failure during incubation and the nestling period (Fitze et al. 2004). Parasite dynamics and impacts are often quantified using two metrics: intensity (i.e. load) and prevalence. Intensity quantifies the number of infections per individual host within a given population, while prevalence is measured as a proportion of infected individuals within a sample from a given host population (Table 5.1). Understanding variation in parasite prevalence and intensity can thus be important for making inferences about the evolution of social behavior and the potential fitness consequences associated with parasite infection.
Table 5.1.
Summary and definitions of commonly used terms associated with parasitism
| Term | Definition | Source |
|---|---|---|
| Host | An animal or plant on which a parasite lives. | Martin (2010) |
| Reservoir host (ecological definition) | Hosts that do not exhibit clinical disease as a result of infection. | Nunn and Altizer (2006) |
| Reservoir host (medical definition) | Hosts that serve as a source of infection and potential reinfection of people and sustain parasite populations when humans are not available. | Martin (2010) |
| Parasite (ecological definition) | Any organism that lives on and draws nutrients from another living organism (the host), usually to the host’s detriment. | Nunn and Alitzer (2006) |
| Parasite (medical definition) | An organism that grows, feeds, and is sheltered on (ectoparasite) or in (endoparasite) a different organism while contributing nothing to survival of its host. | Martin (2010) |
| Microparasite | Pathogens, or disease-causing microbes (viruses, bacteria, protozoa, fungi). | Anderson and May (1979) |
| Macroparasite | Multicellular parasites (helminthes, arthropods, most ectoparasites) | May and Anderson (1979) |
| Disease (ecological definition) | Pathology caused by infection, including outward physical signs and internal or behavioral changes. | Nunn and Alitzer (2006) |
| Disease (medical definition) | A disorder with a specific cause (may or may not be known) and recognizable signs and symptoms. | Martin (2010) |
| Pathogen | Disease-causing agent. | Nunn and Alitzer (2006) |
| Virulence (ecological definition) | Disease-induced host mortality and/or reductions in fecundity | Nunn and Alitzer (2006) |
| Virulence (medical definition) | The disease-producing (pathogenic) ability of a microorganism. | Martin (2010) |
| Infection | Invasion of the body by harmful organisms (pathogens), such as bacteria, fungi, protozoa, or viruses. | Martin (2010) |
| Prevalence | A measure of morbidity based on current levels of disease in a population; measured as a proportion (i.e. number of infections divided by number of individuals). | Martin (2010) |
| Intensity | A measure of morbidity based on current levels of disease in a population; measured as an absolute number (i.e. the total number of infections). | Martin (2010) |
| Epidemiology | The science concerned with the study of the factors determining and influencing the frequency and distribution of disease in a defined human or animal population. | Martin (2010) |
Although empirical data from wild host populations are surprisingly scarce, host behavior is considered an important predictor of parasite intensity, prevalence, and impacts (Moore 2002). A longstanding hypothesis in parasite ecology predicts an influence of the host social system, and the duration and frequency of social contacts within host populations, on parasite prevalence or intensity (Loehle 1995; Altizer et al. 2003). Social systems, defined as groups of conspecifics that regularly interact more frequently with one another than with members of other groups, represent the highest level of sociality, while social organization and social structure describe the size, composition, and spatiotemporal distribution and cohesion of social systems (Table 5.2; Whitehead 2008). Social systems can be further subdivided into two categories: colonies and aggregations, and this distinction is important for understanding host-parasite dynamics in bats. Colonies are groups of individuals that may or may not be genetically related but which exhibit non-random patterns of association, and frequent close contact with each other (Kerth 2008). It is often assumed that groups of bats roosting in a common structure represent a colony but often such groups may not meet the colony definition and, instead, represent aggregations. Aggregations are defined as assemblages of individuals that happen to occur in a shared environment at the same time, perhaps due to an attraction to that environment rather than social bonds with other individuals (Table 5.2). Variation in social dynamics within and between colonies and aggregations can mediate host-parasite dynamics (Webber et al. 2016).
Table 5.2.
Summary and definitions of commonly used terms associated with socioecology and personality in the context of host–parasite interactions of bats
| Term | Definition | Source |
|---|---|---|
| Social structure | The composition of groups and the spatial distribution of individuals. | Whitehead (2008) |
| Social organization | Size, sexual composition, and spatiotemporal cohesion of a society. | Whitehead (2008) |
| Social system | Set of conspecific animals that interact regularly and more so with each other than with members of other such societies. | Whitehead (2008) |
| Society | A group of individuals belonging to the same species and organized in a cooperative manner. | Wilson (1975) |
| Colony | Characterization of roosting associations of bats, which may or may not maintain body contact with each other in a communal roost. ‘Colony’ is commonly used for females breeding communally in maternity colonies. | Kerth (2008) |
| Aggregation | Anonymous assemblage of individuals at the same place. Bats in aggregations show no social bonds and do not engage in cooperative or other affiliate social interactions. | Kerth (2008) |
| Gregarious | An individual’s tendency to associate with conspecifics. | Godde et al. (2013) |
| Sociability | An individual’s reaction to the presence or absence of conspecifics. Sociability ranges along a continuum and sociable individuals tend to seek the presence of conspecifics while unsociable tend to avoid conspecifics. | Réale et al. (2007) |
| Activity | The general level of activity of an individual. | Réale et al. (2007) |
| Exploration | An individual’s reaction to a novel object or situation. | Réale et al. (2007) |
Relationships between host social systems and parasite dynamics are complex, but several mechanisms are predicted to influence these patterns and are likely important for bats. For example, hosts that occur in large, high-density colonies are predicted to have more frequent interactions resulting in more opportunities for parasite transmission (Stanko et al. 2002; Tompkins et al. 2011). Dense aggregations may result in contacts that are fewer and shorter in duration than those occurring in colonies, but are still likely to provide more opportunities for parasite transmission than might occur for solitary bats or those in very small colonies. This variation in host density also has the potential to influence one of the most fundamental parameters of disease ecology, the basic reproduction number or basic reproductive ratio (R0). R0 is an important metric of parasite fitness typically defined, for microparasites, as the number of secondary infections caused by an infectious individual in an entirely susceptible population or, for macroparasites, the number of female larvae established from a single female worm (Hudson et al. 2002). When R0 > 1 infection persists within the host population and when R0 < 1 infection cannot become established (Perkins et al. 2003). Variation in social behavior can impact R0 if certain individual hosts disproportionately infect a large number of conspecifics, and thus inflate R0 above the persistence threshold of one (Lloyd-Smith et al. 2005). Although empirical data from numerous vertebrates (e.g. ungulates: Ezenwa 2004) and invertebrates (e.g. bees: Otterstatter and Thomson 2007) support theoretical relationships between parasitism and sociality, there are few data for most host–pathogen systems involving wild mammals, including bats.
Bats are among the most ecologically diverse of mammals with an enormous range of social systems (Kunz and Lumsden 2003; Kerth 2008; Johnson et al. 2013). For example, colony or aggregation sizes of bats range from completely solitary to millions of individuals, while social systems range from small, closed societies with potentially long-term social bonds (e.g. Thyroptera tricolor: Chaverri 2010) to enormous, likely passive aggregations of individuals attracted to high-quality habitats (e.g. Myotis lucifugus during autumn swarming: Fenton 1969). Social behavior in bats presumably evolved in response to the costs and benefits associated with close conspecific contact. Benefits of social roosting may be numerous and include social thermoregulation, cooperative behavior, and information transfer. Many temperate bats rely on social thermoregulation to decrease energy expenditure during periods of energy limitation, such as pregnancy and lactation (e.g. Eptesicus fuscus: Willis and Brigham 2007), while some tropical species also appear to exploit social thermoregulation (e.g. Uroderma bilobatum: Lewis 1992). The evolution of cooperative behavior in bats was likely facilitated by strong female philopatry and stable group structure (Emlen 1994). Vampire bats (Desmodus rotundus) are well known for their cooperative behavior (i.e. reciprocal altruism) and females rarely transfer between groups (Wilkinson 1987; Carter and Wilkinson 2013). This combination suggests an evolutionary scenario, where stable group structure ultimately led to selection favoring cooperative behavior. Information transfer about predation risk (e.g. Kalcounis and Brigham 1994) and high-quality foraging sites (e.g. McCracken and Bradbury 1981) are also commonly cited as potential benefits of sociality for bats.
Despite these potential benefits of social behavior, risk of infection with micro- and macroparasites is thought to represent a potentially pronounced fitness cost of being social (Côté and Poulin 1995). This cost is illustrated most obviously by the recently emerged infectious disease white-nose syndrome (WNS), which is caused by the fungal microparasite Pseudogymnoascus destructans (Blehert et al. 2009). WNS has resulted in catastrophic declines of temperate hibernating bats in North America (Frick et al. 2010) and prompted urgent conservation and management attention (Foley et al. 2011; Fenton 2012). P. destructans is an invasive pathogen that appears to have evolved with bats from the old world, where it does not cause mortality of infected hosts, and to date, is known to occur on at least 15 hibernating bat species (Puechmaille et al. 2011; Zukal et al. 2016). P. destructans grows in exposed skin of the muzzle, ears, and wing membranes of bats during hibernation (Blehert et al. 2009; Warnecke et al. 2012). For North American species, infection with P. destructans causes an increase in energy expenditure (Verant et al. 2014) and arousal frequency (Boyles and Willis 2010; Reeder et al. 2012; Warnecke et al. 2012) which lead to premature depletion of fat stores during hibernation. Although the mechanism inducing increased energy expenditure and arousals by infected bats is still not fully understood (for review see Willis 2015), variation in social behavior could mediate fungal transmission and growth, especially since affected species tend to hibernate in large colonies or aggregations in caves or mines. Understanding host-parasite dynamics in the context of social behavior for WNS is therefore important from a conservation perspective.
In addition to serious conservation threats for some species, bats also appear to be reservoir hosts for a number of emerging infectious diseases (EIDs) of public health concern (Luis et al. 2013; Plowright et al. 2015). Reservoir hosts tend not to exhibit clinical disease as a result of infection (Baker et al. 2013) and reservoir host populations may therefore provide large pools of infected hosts that could facilitate spillover events to heterospecifics, including humans, livestock, or pets (Luis et al. 2013; Plowright et al. 2015). Interestingly, their apparent ability to tolerate infection with a wide diversity of viral parasites is one factor supporting the recent hypothesis that bats are ‘special’ with respect to their propensity to host zoonotic microparasites (Luis et al. 2013; Brook and Dobson 2015). The recent identification of a number of viral, protozoan, and bacterial microparasites in bats supports this hypothesis and has prompted significant analysis and discussion about bats as natural hosts to microparasites associated with EIDs of humans or livestock (Hayman et al. 2013; Luis et al. 2013; Olival and Hayman 2014; Veikkolainen et al. 2014). For example, recent evidence suggests that bats host more zoonotic viruses per species compared to rodents, and human encroachment into bat habitats, particularly in the tropics, could facilitate spillover events (Daszak et al. 2000; Luis et al. 2013). Several hypotheses have been proposed to explain the apparent zoonotic potential of bats. Enormous variation in body temperature (Tb) and metabolic rate (MR) between rest and sustained flight in many heterothermic bats could reduce fitness or pathogenicity for many viral parasites (O’Shea et al. 2014). Interestingly, widespread torpor expression in bats appears to reduce the likelihood of hosting zoonotic viruses, possibly because viral replication decreases as a result of reduced Tb and MR during torpor (Luis et al. 2013; Stawski et al. 2014). This hypothesis suggests physiological tolerance as a mechanism allowing bats to serve as reservoir hosts, but evidence linking host sociality as an additional mechanism is limited (Hayman et al. 2013).
Despite the fundamental evolutionary importance, and conservation and public health significance of bat-parasite interactions, studies of the influence of bat sociality on parasite transmission and acquisition are limited, especially for microparasites. Here, we review the literature on relationships between social behavior of bats and their associated parasites. First, we provide an overview of the tools, techniques, and methodologies that have been used to quantify relationships between sociality and parasitism in bats, as well as relevant techniques that have been used for other vertebrates and which could be applied to bats. Second, we summarize the role of sociality in three relatively well-studied bat-parasite systems: WNS, Hendra virus (HeV), and arthropod macroparasites. Finally, we propose testable hypotheses and observational and experimental studies important for understanding the influence on parasite dynamics of two important concepts in behavioral ecology of bats: fission–fusion social organization and individual behavioral tendencies (i.e. personality).
Tools, Techniques, and Methodologies
We conducted an extensive search of the literature and compiled a list of articles based on combinations of key word searches on Web of Science. We searched the term “Chiroptera” and “bat” with every possible combination of social*, gregarious*, colony, aggregation, fission–fusion (i.e. a common type of bat social system, see below), viral, pathogen, disease, ectoparasite*, endoparasite*, parasite*, infection* and epidemiology*. Our initial search yielded 223 unique articles but we eliminated articles that did not explicitly quantify or discuss a link between some aspect of sociality and parasite risk. This left only 35 articles that fully satisfied our search criteria. Four of these used comparative analyses to examine effects of species-specific socioecological traits on parasite risk (Table 5.4), 15 addressed sociality in the context of microparasites (Table 5.4), and 16 addressed sociality in the context of macroparasites (Table 5.5).
Table 5.4.
Summary of published articles explicitly addressing links between microparasites and some aspect of host social behavior in bats
| Species | Family | Location | Disease/Pathogen | Social aspect | Analysis | Ref |
|---|---|---|---|---|---|---|
| Myotis lucifugus, M. septentrionalis, M. sodalis, M. leibii, Perimyotis subflavus, Eptesicus fuscus | Vespertilionidae | North America | White-Nose Syndrome (WNS) | Effect of hibernation cluster size on WNS detection | Population surveys, statistical models | 1 |
| M. lucifugus, M. septentrionalis, M. sodalis, M. leibii, P. subflavus, E. fuscus | Vespertilionidae | North America | WNS | Variation in sociality between species | Population surveys, statistical models | 2 |
| M. lucifugus | Vespertilionidae | North America | WNS | Female philopatry and colony connectivity | Population Genetic Structure (FST) | 3 |
| M. lucifugus | Vespertilionidae | North America | WNS | Effect of colony size on WNS risk | Population surveys, statistical models | 4 |
| M. lucifugus, M. septentrionalis, M. sodalis, M. leibii, P. subflavus, E. fuscus, M. emarginatus, M. myotis, M dasycneme, M. brandtii, M. mystacinus, M. daubentonii, M. nattereri, Pipistrellus pipistrellus, E. nilssonii, E. serotinus | Vespertilionidae |
North America & Europe |
WNS | Variation in pre- and post-WNS colony size of North American and European bat species | Population surveys and statistical models | 5 |
| E. fuscus | Vespertilionidae | USA | Rabies | Effect of colony size on rabies acquisition | Field data, statistical models | 6 |
| E. fuscus | Vespertilionidae | Canada & USA | Hypothetical | Variation in fission–fusion dynamics between pregnant and lactating bats influences hypothetical pathogen dynamics | Social network analysis, epidemiological models | 7 |
| Nyctalus lasiopterus | Vespertilionidae | Spain | Hypothetical | Variation in roosting behavior and fission–fusion dynamics influences hypothetical pathogen dynamics | Social network analysis, epidemiological models | 8 |
| Desmodus rotundus | Phyllostomidae | Peru | Rabies | Effect of migration and group size on rabies prevalence | Field data, epidemiological models | 9 |
| D. rotundus | Phyllostomidae | Peru | Rabies | Effect of colony size on rabies seroprevalence | Field data, statistical models | 10 |
| M. myotis | Vespertilionidae | Germany | Coronaviruses, Astroviruses, and Adenoviruses | Effect of colony size on viral amplification | PCR to identify viruses, field data, and statistical models | 11 |
| Pteropus giganteus | Pteropodidae | Bangladesh | Nipah virus | Effect of roost selection and colony size on predicted Nipah outbreaks | Field data, epidemiological models | 12 |
| P. alecto, P. poliocephalus | Pteropodidae | Australia | Hendra virus | Colony size and absence of migratory behavior of urban populations | Field data, epidemiological models | 13 |
| Eptesicus serotinus, Hypsugo savii, M. blythii, M.capaccinii, M. daubentonii, M. emarginatus, M. escalerai, M. myotis, Nyctalus leisleri, Plecotus auritus, P.austriacus, P. kuhlii, P. nathusii, P. pipistrellus, P. pygmaeus, Miniopterus schreibersii, Rhinolophus euryale, R. ferrumequinum, R. hipposideros, Tadarida teniotis |
Vespertilionidae Miniopteridae Rhinolophidae Molossidae |
Spain |
European bat Lyssavirus |
Effect of colony size on Lyssavirus seroprevalence | Field data, seroprevalence, and statistical models | 14 |
| Hypothetical | – | – | Hypothetical | Effects of fission–fusion dynamics, group size, and information exchange on predicted pathogen dynamics | Social network analysis, epidemiological models | 15 |
[1] Langwig et al. (2012), [2] Langwig et al. (2015), [3] Miller-Butterworth et al. (2014), [4] Wilder et al. (2011), [5] Frick et al. (2015), [6] Pearce et al. (2007), [7] Webber et al. (2016), [8] Fortuna et al. (2009), [9] Blackwood et al. (2013), [10] Streicker et al. (2012), [11] Drexler et al. (2011), [12] Hahn et al. (2014), [13] Plowright et al. (2011), [14] Serra Cabo et al. (2013), [15] Kashima et al. (2013)
Table 5.5.
Summary of published articles explicitly addressing the links between macroparasites and some aspect of host social behavior in bats
| Species | Family | Location | Ectoparasite species | Social Aspect | Analysis | Ref |
|---|---|---|---|---|---|---|
| Myotis myotis, M. bechsteinii | Vespertilionidae | Switzerland, Italy, and Germany | Spinturnix myoti,S. bechsteini | Effect of host social system on parasite genetic structure | Field data, statistical analysis | 1 |
| Lophostoma silvicolum | Phyllostomidae | Panama | Streblid flies, Wing mites | Effect of mating and social systems | Review analysis | 2 |
| Noctilio albiventris, N. leporinus, Glossophaga soricina, Desmodus rotundus, Artibeus fimbriatus, A. jamaicensis, A. lituratus, Platyrrhinus lineatus, Pygoderma bilobatum, Sturnira lilium, Eptesicus fuscus, Lasiurus ega, M. albescens, M. nigricans, Eumops glaucinus, E. patagonicus, Molossops temminckii, Molossus ater, M. currentium, M. molossus, Nyctinomops laticaudatus |
Noctilionidae Phyllostomidae Vespertilionidae Molossidae |
Paraguay | Noctiliostrebla maai, N. aitkeni, Paradyschiria parvula, P fusca, Ornithodoros hasei, Trichobius joblingi, T. parasiticus, Strebla weidemanni, Radfordiella desmodi, Periglischrus iheringi, P. ojasti, Macronyssoides kochi, M. conciliates, Paratrichobius longicrus, Aspidoptera falcata, Megistopoda proxima, Parichoronyssus euthysternum, Spinturnix surinamensis, Steatonyssus joaquimi, S. furmani, Macronyssus crosbyi, Hesperoctenes sp., H. parvulus, Chiroptonyssus haematophagus, C. venezolanus, C. robustipes, Parkosa maxima, P. tadarida | Effect of sex-bias in host social system on ectoparasite abundance | Field data, statistical analysis | 3 |
| Antrozous pallidus | Vespertilionidae | USA | Steatonyssus antrozoi, Spinturnix orrii, Basilia antrozoi, Whartonia whartoni, Ornithodorus sp. | Host roost switching and grouping behavior | Field data, statistical analysis | 4 |
| N. albiventris, N. leporinus, Glossophaga soricina, Carollia perspicillata, D. rotundus, A. fimbriatus A. jamaicensis, S. lilium, M. nigricans, M. molossus, M. rufus |
Noctilionidae Phyllostomidae Vespertilionidae Molossidae |
Paraguay | Noctiliostrebla maai, N. aitkeni, N. dubia, Paradyschiria parvula, P. fusca, Xenotrichobius noctilionis, Strebla guajiro, S. weidemanni, Trichobius dugesii, T. uniformis, T. joblingi, T. parasiticus, T. jubatus Aspidoptera phyllostomatis, A. falcat,a Megistopoda aranea,.M proxima Metelasmus pseuodopterus, M. wenzeli, Basilia speiseri, B carteri, Hesperoctenes fumarius | Effect of host aggregation and social structure on ectoparasite abundance | Field data, statistical analysis | 5 |
| M. bechsteinii | Vespertilionidae | Germany | S. bechsteini | Effect of host social system on ectoparasite population structure | Field data, genetic analysis, statistical analysis | 6 |
| M. bechsteinii | Vespertilionidae | Germany | B. nana, S. bechsteini | Effect of colony size and social system on ectoparasite abundance | Field data, statistical analysis | 7 |
| M. bechsteinii | Vespertilionidae | Germany | B. nana | Effect of sociality and roost switching on ectoparasite reproductive success | Field data, statistical analysis | 8 |
| M. myotis, M. blythii, M. daubentonii, Plecotus auritus, Nyctalus noctula | Vespertilionidae | Switzerland | S. andegavinus, S. myoti | Host sex-bias in social system | Experimental trials, field data, and statistical analysis | 9 |
| Miniopterus schreibersii | Vespertilionidae | Portugal | S. psi, S. myoti, Macronyssus granulosus, M. longimanus | Host sex-bias in social system | Field data, statistical analysis | 10 |
| M. lucifugus | Vespertilionidae | Canada | Myodopsylla insignis, S. americanus, M. crosbyi, Leptotrombidium myotis, Cimex sp. | Host sex-bias in social aggregation | Field data, statistical analysis | 11 |
| M. myotis | Vespertilionidae | Poland | S. myoti | Effect of host colony size on ectoparasite abundance | Field data, statistical analysis | 12 |
| Tylonycteris pachypus, T. robustula | Vespertilionidae | China | Macronyssus radovskyi | Effect of host colony size on mite abundance | Field data, statistical analysis | 13 |
| Myotis daubentonii | Vespertilionidae | Czech Republic | S. andegavinus | Effect of host colony size and sex-bias in social aggregation | Field data, statistical analysis | 14 |
| M. daubentonii | Vespertilionidae | Germany | S. andegavinus | Effect of host colony size on mite abundance | Field data, statistical analysis | 15 |
| M. bechsteinii | Vespertilionidae | Germany | B. nana | Effect of host roost selection on ectoparasite infestation | Experimental trials, field data, statistical analysis | 16 |
[1] van Shaik et al. (2014), [2] Dechmann and Kerth (2008), [3] Presley and Willig (2008), [4] Lewis (1996), [5] Presley (2011), [6] Bruyndonckx et al. (2009), [7] Reckardt and Kerth (2009), [8] Reckardt and Kerth (2006), [9] Christe et al. (2007), [10] Lourenço and Palmeirim (2007), [11] Webber et al. (2015a), [12] Postawa and Szubert-Kruszynska (2014), [13] Zhang et al. (2010), [14] Luçan (2006), [15] Encarnaçao et al. (2012), [16] Reckardt and Kerth (2007)
The studies we identified employed a range of methodologies to quantify links between sociality and parasite risk (Tables 5.3, 5.4 and 5.5). Methods included social network analysis (n = 3), epidemiological modeling (n = 5), and interspecific comparative analyses (n = 4). Details about the underlying theory and the implementation of these methods is available elsewhere (e.g. social network analysis: Croft et al. 2008; epidemiological modeling: May 2006; comparative analyses: Garland et al. 2005), and here we focus on the progress that has been made to date in identifying knowledge gaps where future research on bat-parasite dynamics could be focused.
Table 5.3.
Summary of published articles using phylogenetically corrected comparative analyses to test the effects of species-specific socioecological traits on parasitism in bats
| Number of bat species (families) | Location | Metric of parasitism | Social aspect | Analysis | Source |
|---|---|---|---|---|---|
| 33 (7) | Southeast Asia |
Viral richness, ecto- and endoparasite richness |
Categorical metric of colony size and roost type | Phylogenetic least squares (PGLS) comparative analysis | 1 |
| 33 (5) | Global | Viral richness | Colony size and population genetics (FST) | PGLS comparative analysis | 2 |
| 66 (8) | Global | Viral richness | Life-history traits: nonsignificant | PGLS comparative analysis | 3 |
| 15 (6) | Africa | Viral richness | Colony size and roost type: nonsignificant | PGLS comparative analysis | 4 |
Social Network Analysis
Social network analysis is based in mathematical graph theory and has been widely applied in human sociobiology, since the 1950s (e.g. Cartwright and Harary 1956; Wasserman and Faust 1994). For studies of wildlife, network analysis was first applied in primate sociobiology (e.g. Sade and Dow 1994), and has recently been applied more broadly to many other taxa (e.g. Fewell 2003; Hamede et al. 2009; Drewe 2010), including bats (for review see Johnson et al. 2013). Networks consist of nodes (individuals or locations) and edges (interactions between nodes) through which a variety of individual and group level metrics can be quantified (Wey et al. 2008). The roles and importance of individuals or locations can then be assessed based on these metrics (see Croft et al. 2008 for review). For example, individuals with certain combinations of traits can influence network dynamics by potentially acting as intermediaries connecting smaller subgroups within larger groups (Wey et al. 2008; Krause et al. 2010). In addition, bipartite networks (or two-mode networks) can be constructed to assess associations between individuals and ecologically relevant locations (e.g. a population of organisms and their nesting sites). Network metrics are especially useful because they can be used as predictor variables for relevant-dependent variables (e.g. infection status or parasite intensity) in standard statistical models (e.g. general linear models). Network analysis has allowed disease ecologists and parasitologists to make important strides quantifying how non-random social interactions affect parasite transmission and dynamics (for review see Godfrey 2013).
Network analyses have become increasingly popular for studies of bats (Johnson et al. 2013). For instance, network analyses have been applied in a bat-habitat management context by identifying critical roosting locations that serve as ‘hubs’ within a roost network and simulating the consequences of removing those key sites for stability of the social group (Rhodes et al. 2006; Silvis et al. 2014). Network analyses have also been used to identify and quantify fission–fusion behavior within bat colonies (Fortuna et al. 2009; Patriquin et al. 2010; Kerth et al. 2011; Johnson et al. 2012). In the context of disease ecology, so far only two studies have connected social networks with epidemiological models to make inferences about host–pathogen dynamics (e.g. Fortuna et al. 2009; Webber et al. 2016). Fortuna et al. (2009) showed that giant noctule bats (Nyctalus lasiopterus) form highly modular, fission–fusion colonies, and used an epidemiological model, parameterized using network metrics, to show that this arrangement reduces the spread of information or disease within the population because the colony was effectively segregated into modules divided among many roost trees (n = 73 trees for 25 bats: Fortuna et al. 2009). For big brown bats (Eptesicus fuscus), Webber et al. (2016) recently showed that network structure depends on the habitat context. Although tree-roosting E. fuscus reuse tree hollows between years (Willis et al. 2003), they frequently switch roosts within years (on average every 1.7 days: Willis and Brigham 2004) and rarely return to the same roost within a given summer. In building roosts, however, E. fuscus switch much less frequently and commonly return to the same roost repeatedly within the same year (Ellison et al. 2007; Webber et al. 2016). Webber et al. (2016) applied epidemiological models to social networks constructed based on these patterns of roosting behavior in forests versus buildings and found that parasite dynamics should vary between these habitat contexts with more rapid pathogen dissemination within building roosting colonies.
Constructing epidemiological models from empirical data undoubtedly improves inference for predicting epidemic outcomes, but as yet there are no studies that quantify real-world relationships between the social behavior of bat hosts and their parasites to test predictions of epidemiological models (Table 5.3). Although this form of data collection can be labor-intensive and potentially expensive (Craft and Caillaud 2011), there are numerous examples of network analysis being used to integrate information on host-contact patterns and parasitism in systems with highly gregarious hosts. For example, network analysis elucidated the role of meerkat (Suricatta suricata) allogrooming as a predictor for the prevalence of Myobacterium tuberculosis, the microparasite that causes tuberculosis (Drewe 2010). Quantifying association patterns of meerkats was largely conducted via focal animal observation, a type of data collection that is nearly impossible for free-ranging bats. Thus, advances in technology, such as passive transponders (PIT tags), data-logging telemetry, and/or proximity data loggers may be useful for quantifying association patterns and constructing social networks for bats with implications for epidemiology and disease ecology (e.g. Willis and Brigham 2004; Patriquin et al. 2010; Kerth et al. 2011; Johnson et al. 2012).
Epidemiological Modeling
Although empirical data are sparse, host-parasite dynamics, and the impacts of parasites on host populations have a rich theoretical history founded on Anderson and May’s (1979) seminal models. Their classic microparasite epidemiological model conceptualizes host population dynamics in terms of susceptible (S), exposed (E), infected (I), and resistant/recovered (R) population pools, and these pools can be combined in a range of ways depending on the nature of host–pathogen interactions in the wild (e.g. SI, SIS, SIR, SEIR models). In a standard SIR model, individuals transition from S to I as a function of transmission rate (β) and from I to R as a function of survival of infection (v), while birth (a), and death rates (b, a + b for infected hosts) of each pool drive overall population dynamics (Fig. 5.1; Anderson and May 1979). Since being developed, SIR epidemiological models have been widely used to infer microparasite impacts on host populations for humans (e.g. Anderson and May 1982), wildlife (e.g. McCallum et al. 2009) and plants (e.g. Gilligan et al. 1997).
Fig. 5.1.
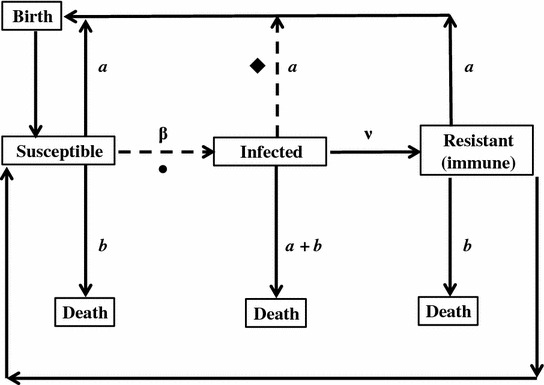
Flow chart of the original compartment model of microparasite impacts on host populations based on population pools of susceptible, infected, resistant individuals (modified from Anderson and May 1979). Hosts shift from susceptible to infected as a function of transmission rate (β) and infected to resistant as a function of survival of infection (v), while birth (a) and death (b, a + b for infected hosts) rates drive population dynamics. Variation in host social behavior could impact β (•), which mediates the number of infected individuals as well as the subsequent fitness of infected individuals via reproductive rate (♦)
Although appropriate for modeling the impacts of many infectious diseases (e.g. measles, whooping cough: Anderson and May 1992), classic SIR models compartmentalize individuals into broad categories which may fail to capture variation in aspects of host biology that could influence parasite dynamics and impacts (Keeling and Eames 2005). For example, classic SIR models often assume that individual hosts in the population associate at random but, particularly for highly social species, non-random association patterns are far more likely in nature. Incorporating network structure into epidemiological models eliminates random-mixing assumption of classic SIR models by assigning each individual a finite number of permanent or temporary contacts between which transmission can occur (Keeling and Eames 2005; May 2006). Implications for network epidemiological modeling include the ability to identify highly connected individuals that may be involved in a disproportionate number of transmission events (e.g. super-spreaders: Lloyd-Smith et al. 2005) and weakly connected individuals that may benefit from lower risk of infection. Epidemiological models informed by variation in social connectedness can be highly effective for predicting pathogen dynamics (Lloyd-Smith et al. 2005) but, as noted above, so far only two studies have applied these methods specifically to bats (Fortuna et al. 2009; Webber et al. 2016). However, to our knowledge no study has explicitly tested model predictions on a natural bat-parasite system in the wild.
An alternative to network epidemiological models informed by short-term (i.e. one season) association data, is the use of long-term population data for model parameterization. These data ideally include information on changes in population or colony sizes and changes in population demographic structure and/or vital rates. Most often for bats, long-term field data on colony size and population connectivity, in combination with laboratory data on immune physiology have been used (Table 5.4; George et al. 2011; Plowright et al. 2011). For example, field and laboratory data were combined to model ecological factors driving HeV spillover events occurring in Australia from flying foxes (Pteropus sp.) to horses and predicted that decreased migratory behavior of the host increased host density in urban areas, which contributed to the duration and intensity of HeV outbreaks (Plowright et al. 2011, see below). Similarly, mark-recapture data, combined with birth and death rates of different demographics, have been used to parameterize models of rabies dynamics (George et al. 2011). Rabies in E. fuscus is highly seasonal, with peaks in prevalence at maternity colonies (i.e. during June and July) after parturition when pups provide a supply of immunologically naive hosts (George et al. 2011). Thus, the chance of spillover is greatest during the post-parturition period when females roost in large colonies and the influx of juveniles in the population increases overall prevalence.
These studies highlight the value of predictive models incorporating the behavior of bats to make predictions about parasite dynamics, with potential implications for human public health. However, even models which incorporate host ecology and behavior (e.g. habitat selection) do not typically account for variation in social behavior which could also play an important role (Moore 2002). For microparasites, the two most likely parameters influenced by variation in social behavior are transmission rate (β) and the survival and subsequent reproduction of infected individuals (Fig. 5.1). For example, recent appreciation of individual differences in host behavior has led to the realization that homogenous mixing or so-called ‘mass action’ transmission may not reflect the dynamics of most host-parasite systems (McCallum et al. 2001; Moore 2002; Barber and Dingemanse 2010). Despite this realization, though, for many host–pathogen systems, estimating β can be difficult and modeling studies tend to rely on some plausible range of values for β (e.g. Griffin and Nunn 2012). For some microparasites, however, it may be possible to experimentally manipulate transmission and determine β empirically. For example, a basic pairwise experiment, where pairs of individuals (one infected, one susceptible) are housed together could help to determine the proportion of contacts that result in transmission, and narrow the range of plausible β values. Incorporating empirically derived data on association patterns of individual bats into epidemic models could help to improve our predictions of potential epidemic scenarios. These analyses (i.e. informed by empirical data) are still relatively scarce for bats (Table 5.4), but recent interest in host-parasite interactions and social network analysis provides an excellent opportunity to connect real data with model predictions.
Interspecific Comparative Analyses
Comparative analyses provide insight into potential species-specific variation in evolutionary adaption (i.e. changes in response to natural selection) within a group of organisms (Garland et al. 2005). From the perspective of parasites, a host is effectively a habitat, which means principles of community ecology, and biotic and abiotic characteristics of the “host ecosystem,” can be used to explain patterns of parasite community composition (Morand and Poulin 1998). Thus, in contrast to intraspecific analyses which typically assess variation in prevalence or intensity of infection among individuals, comparative studies typically examine interspecific variation in parasite species richness among host species because intensity and prevalence are unlikely to be comparable in terms of their impacts, across different host and parasite species. In most cases, comparative studies of parasite species richness “correct for” host phylogeny. Two closely related species are more likely to share similar functional traits compared to two more distantly related species because of their more recent evolutionary history (Ives and Garland 2010). More parasites are also likely to have been identified for host species that have been well-studied so, in addition, most comparative studies also account for publication bias by including the total number of publications on a given host species as a covariate in statistical models, or by using residuals of a linear regression between number of publications and parasite species richness as the response variable (e.g. Nunn et al. 2003; Lindenfors et al. 2007; Turmelle and Olival 2009).
To date, relatively few studies have used comparative analyses to test effects of social behavior on parasite species richness in bats and, so far, most have focused on viruses with one quantifying macroparasites (Table 5.3). In contrast, studies of primates, the best-studied mammalian taxon in terms of relationships between social behavior and parasite richness, focus on a wider breadth of parasites (i.e. ecto and endoparasites as well as bacterial and viral pathogens: Nunn et al. 2003; Vitone et al. 2004; Griffin and Nunn 2012). For bats, colony size appears to be one of the most important predictors of viral richness, and epidemiological and evolutionary theory predict that highly colonial bat species should harbor greater parasite diversity, because of increased opportunities for parasite reproduction and evolution (Altizer et al. 2003). However, observed relationships have not always followed this pattern for bats (Turmelle and Olival 2009; Gay et al. 2014). Turmelle and Olival (2009) found no effect of colony size on viral richness in bats but, rather, identified species’ conservation status and global FST (i.e. population genetic structure) as the most important predictors. On the other hand, among Southeast Asian bats, Gay et al. (2014) found a negative relationship between colony size and viral richness, which contradicts epidemiological theory (Loehle 1995). One potential limitation of these results could be that the authors used a categorical metric of population size (i.e. small, medium or large), which likely underrepresents the complexity of social behavior (Gay et al. 2014). Alternatively, in the case of Gay et al.’s (2014) results, it could be that bats in large colonies express immune or behavioral traits that provide protection from increased parasite risk in that social context.
Although they have still not been widely used, in our view comparative studies have great potential to help understand how bat social behavior influences parasite diversity and co-evolves with parasites, in part because bats are so diverse and exhibit a wide range of social and mating systems. For example, promiscuous bat species should host a greater number of parasite species compared to harem-breeding and monogamous bats because, independent of colony size or density, polygynandrous species presumably come in contact with a greater number of conspecifics during mating compared to polygynous species, which likely has implications for host-parasite coevolution. However, it is important to note that mating systems vary considerably among bats (McCracken and Wilkinson 2000) and other aspects of bat social systems (e.g. degree of fission-fusion or roost fidelity: see below) likely co-vary with mating system and are equally important as predictors of parasite richness. Thus, we recommend that future studies attempt to include more detailed information on bat social systems in comparative analyses. For example, a meta-analysis of primate social structure showed that differences in social network modularity (i.e. sub-grouping within a larger group) in 19 species led to differences in disease dynamics among species (Griffin and Nunn 2012). Although it may not be possible to include such detailed social association data for bats at present, as more social network studies accumulate, we recommend the use of network metrics as predictor variables of parasite richness in comparative analyses. Species-specific data on typical contact rates among individuals and demographics, and between sympatric species, as well as data on dispersal patterns and group stability within bat species (e.g. fission-fusion) would also be valuable as predictor variables to better understand how host social traits in bats influence their parasite ecology and evolution.
Examples from the Field: White-Nose Syndrome, Hendra Virus, and Arthropod Ectoparasites
Although very different in terms of their ecology and population impacts, based on our literature review, the relationship between bat sociality and parasite impacts and dynamics has been relatively well-studied for three sets of parasites: P. destructans, the cold-tolerant fungal pathogen of bats that causes WNS; HeV, which can lead to potentially fatal zoonotic disease in humans and horses; and the many species of arthropod ectoparasites which have been relatively well-studied because they can be easily sampled from bats captured in the field. We also identified four articles which addressed the relationship between rabies, or European Lyssavirus, and bat social behavior (Table 5.4), but rabies in bats has been the focus of a number of reviews (e.g. Messenger et al. 2003; Kuzmin and Rupprecht 2006; Rupprecht et al. 2011; O’Shea et al. 2011) so here we focus on less well-characterized bat-parasite interactions.
Host Sociality and Phenology in Bat WNS
WNS is an EID of urgent conservation concern because it is causing staggering rates of mortality among hibernating bats in eastern North America (Blehert et al. 2009; Frick et al. 2010; Wilder et al. 2011; Langwig et al. 2012; Frick et al. 2015). Despite its recent emergence (2006–07), however, WNS is among the most well-studied microparasites of bats in the context of social behavior. Interestingly, one of the hallmark behaviors observed for P. destructans infected bats, both in the wild and in the laboratory, is a reduction in clustering behavior as WNS infection progresses over the course of hibernation (Langwig et al. 2012; Wilcox et al. 2014). This phenomenon could lead to increased energy expenditure or evaporative water loss which has implications for survival and potentially future reproduction (Willis et al. 2011; Boratyński et al. 2015). Reduced clustering could be part of a stereotyped behavioral response by animals to infection, known as “sickness behavior” (Hart 1988), which could either reduce the likelihood of becoming infected, or reinfected, by sick individuals in the hibernaculum or reduce the likelihood of infecting susceptible individuals, which could be important for inclusive fitness if bats cluster with kin during hibernation (Wilcox et al. 2014; Bohn et al., in review). In the laboratory, transmission occurs via direct physical contact and there is no evidence of airborne transmission (Lorch et al. 2011), while environmental transmission in the wild likely contributes strongly to infection dynamics because P. destructans can survive in hibernacula in the absence of bats (Verant et al. 2012; Hoyt et al. 2015). Although the most adversely affected bat species tend to associate with large numbers of conspecifics throughout the year, transmission appears to occur almost exclusively within hibernacula during autumn and throughout the winter (Langwig et al. 2015).
Variation in sociality within and between bat species appears to influence host-parasite dynamics in WNS. In the context of host density, there are two theories relevant to the relationship between transmission and host-parasite dynamics. First, density-dependent transmission predicts that host infection scales as a function of host density so that, at low host density, pathogen transmission declines, and a given pathogen fades out (Greer et al. 2008). Second, frequency-dependent transmission predicts that host infection is driven by the total number of interactions among infected and susceptible individuals in the population, regardless of population density, so that when host density is low, a given pathogen can persist (Greer et al. 2008). Interestingly, the relationship between WNS transmission and host density during hibernation appears to be species-specific. Among relatively solitary species (i.e. Perimyotis subflavus and Myotis septentrionalis) that hibernate individually or in small clusters, aggregation size (i.e. the numbers of bats in the hibernaculum) best predicted the number of surviving bats within a given hibernaculum, with larger aggregations experiencing faster apparent declines (Langwig et al. 2012). This observation is consistent with density-dependent transmission models. In contrast, for species that vary more widely in aggregation size (i.e. M. lucifugus and M. sodalis), mortality was equally severe across a range of aggregation sizes (Langwig et al. 2012), which is consistent with frequency-dependent transmission models. This suggests that differences in species-specific social behavior during hibernation can modulate WNS transmission and infection.
Unlike other infectious diseases of bats (e.g. rabies: George et al. 2011) demographic structure in the active season does not appear to drive WNS dynamics. Healthy M. lucifugus emerge from hibernation over an approximately eight-week period in spring (Norquay and Willis 2014; Czenze and Willis 2015). Females tend to emerge prior to males, with the females in the best condition emerging first. This is presumably because larger fat reserves allow females to cope with potentially inclement weather, but also initiate reproduction earlier by exploiting warmer, passively heated maternity roosts, and occasional warm nights with flying insects available (Norquay and Willis 2014). However, bats infected with P. destructans tend to emerge from hibernation much earlier than healthy bats presumably because their fat stores are depleted. Moreover, bats that survive hibernation with WNS often have severe wing damage in spring and individuals with the most damaged wings tend to have the smallest energy reserves (Fuller et al. 2011). For many diseases, an influx of immunologically naive hosts (i.e. juveniles) leads to a dramatic increase in prevalence immediately following reproduction due to vertical transmission from mothers to offspring. In WNS, however, seasonal dynamics of the disease are driven by dramatic seasonal changes in host physiology, specifically sustained low body temperature during hibernation (Warnecke et al. 2012; Langwig et al. 2012, 2015). For P. destructans-infected bats in spring and early summer, an increase in body temperature limits infection and transmission despite the fact that females tend to aggregate in potentially large maternity colonies (Langwig et al. 2015).
Although transmission likely does not occur readily for females at maternity colonies, high mortality rates during hibernation may decrease the number of potentially reproductive females that form maternity colonies (Langwig et al. 2015). Therefore, normal benefits incurred from colonial roosting, such as social thermoregulation, may be dramatically reduced for WNS-surviving bats, which could have additional negative impacts on survivors. For example, the energetic costs of wound healing may be significant for infected individuals upon emergence from hibernation (Fuller et al. 2011) and the decreased availability of social thermoregulation and increased likelihood of roosting solitarily may further increase energetic expenditure (Wilcox and Willis 2016). Therefore, for species affected by WNS, the lack of potential colony members, and thus potential for social thermoregulation, during spring and early summer could ultimately result in decreased survival and reproduction (Langwig et al. 2015).
In addition to influencing transmission of P. destructans, theory predicts that population size influences risk of extinction (de Castro and Bolker 2005). Using counts of bats during winter hibernaculum surveys as a proxy for population size, Frick et al. (2015) showed that, for five of six WNS-affected species, probability of local extinction from a given hibernaculum decreased as population size increased. This suggests that host aggregation and social behavior could influence risk of extinction from WNS. One possible mechanism is that larger populations have greater flexibility to cope with decreases in population numbers, and are thus less likely to face declines below population thresholds where extinction becomes inevitable.
Interestingly, social behavior may also help to explain differences in the consequences of P. destructans infection for European versus North American bats. Prior to the emergence of WNS, hibernating aggregations of affected North American species were approximately tenfold higher, on average, than colony sizes of ecologically similar European species (Frick et al. 2015). However, after WNS emergence, colonies of affected North American species have declined precipitously and appear to be stabilizing at sizes similar to those of European bats (Frick et al. 2015). This suggests the possibility that European colony sizes could have been larger in the past and similar to pre-WNS colony sizes in North America (Frick et al. 2016). Taken together, empirical data from North America and inferences from Europe suggest that WNS could select against larger colony sizes (Frick et al. 2016) resulting in remnant colonies which are, on average, smaller and less social than pre-WNS colonies. We suggest that future studies attempt to quantify social tendencies at the individual and population levels for pre- and post-WNS infected populations to assess the impact of this conservation pathogen for social evolution in bats.
Anthropogenic Changes to Flying Fox Socioecology: Implications for HeV
In Australia, bats from the genus Pteropus are reservoir hosts for HeV, a recently emerged zoonotic virus from the family Paramyxoviridae. Flying foxes are only briefly infectious with no clinical disease but shed HeV in urine, saliva, feces, and placental fluids (Halpin et al. 2011). HeV is lethal to horses and humans, and transmission presumably occurs when horses ingest food or water contaminated by bats (Plowright et al. 2008). Horses appear to act as amplifier hosts (Daszak et al. 2006), as all human cases have been directly traced back to an infected horse (Field et al. 2012). Although, spillover events from flying foxes to horses are complex and few data exist linking individual social tendencies of the bats to HeV transmission, host socioecology likely mediates host–pathogen dynamics for this zoonotic disease.
Colony/aggregation sizes of Pteropus species can range from as few as 10,000 individuals in P. alecto maternity colonies to millions of individuals for P. scapulatus. Based on experimental data and closely related human paramyxoviruses (e.g. measles), HeV may require large host population sizes to provide enough susceptible individuals for persistence because of a relatively short infectious phase and life-long host immunity (Daszak et al. 2006). Historically, naturally occurring nectar resources in native forests supported large, seasonally-migrating flying fox populations (Eby 1991; Plowright et al. 2015) but human-mediated habitat alterations have resulted in patchily distributed natural food resources, which were already ephemeral in nature (i.e. seasonally produced nectar). Therefore, flying foxes have begun to colonize urban and periurban areas to exploit highly abundant, consistently available anthropogenic food resources, such as fruiting trees planted in gardens or horse paddocks (Eby et al. 1999). In many cases, urban bats do not migrate because food resources are consistently available, which increases the likelihood of HeV-positive bats coming in contact with horses (Plowright et al. 2011). Therefore, recent emergence and spillover of HeV in Australia appears directly linked to increases in the size and abundance of urban bat colonies as well as reduced migratory behavior (Plowright et al. 2015). Although urban bat aggregations appear to remain relatively large throughout the year, temporal dynamics of HeV are at least partially mediated by host socioecological traits such as colony size and migration (Plowright et al. 2011).
Host Colony Size and Arthropod Ectoparasites in Female Temperate Bats
As noted above, social group size has been identified as a potentially important predictor of parasitism, with larger groups generally hosting more parasites (Côté and Poulin 1995; Rifkin et al. 2012; Patterson and Ruckstuhl 2013). Among temperate bats, sexual segregation and variation in social group size between sexes appears to be among the most important mediators of ectoparasitism. Adult males tend to roost individually or in small groups, while females form maternity colonies (Kunz and Lumsden 2003). For males, parasitism tends to remain low-throughout spring and early summer and as the mating season progresses ectoparasite prevalence and intensity increases (Webber et al. 2015a). Most studies investigating the link between colony size and ectoparasitism in bats have found the predicted positive relationship (e.g. Luçan 2006; Reckardt and Kerth 2009; Encarnaçao et al. 2012), although this is not always the case (e.g. Zhang et al. 2010; Postawa and Szubert-Kruszynska 2014). One explanation for lack of correlation between group size and ectoparasitism at female colonies in some studies may be linked to variation in host association patterns and roost selection/ephemerality. For instance, bamboo shoots used as roosts by flat-headed bats (Tylonycteris pachypus and T. robustula) are highly ephemeral and likely contribute to frequent roost-switching (Lewis 1995) as well as variation in group size and composition (Zhang et al. 2010). Frequent roost switching breaks up social contacts that could favor direct host–host transmission of ectoparasites. Meanwhile, because some ectoparasites rely on stable roost structures for a portion of their reproductive cycles, bats that use ephemeral roosts rarely come in contact with ectoparasites that rely on a stable roost for reproduction (Lewis 1995), which could supersede the theoretical relationship between colony size and parasite risk.
Although, social contacts among females help to explain observed patterns of ectoparasitism at maternity colonies, social organization (i.e. composition of groups) can also mediate ectoparasitism. Most females present at maternity colonies are either pregnant or lactating and links between temporal variation in social organization (i.e. transition from pregnancy to lactation) and ectoparasitism are supported by co-evolution of some host-ectoparasite reproductive cycles (Christe et al. 2000). For example, reproductive cycles of some mite species respond to pregnancy hormones of their female hosts (Lourenço and Palmeirim 2007). Once pups are born there is also a pulse of mites, which results in significant vertical transmission from adult females to juveniles (Christe et al. 2000; Luçan 2006). Lactating females may also tolerate significant ectoparasite burdens. During lactation, females may have less energy available to mount behavioral (i.e. grooming) or immunological responses to infection (Zuk and McKean 1996; Christe et al. 2000). Thus, while females are lactating and pups are non-volant, ectoparasite prevalence, and intensity can be exceptionally high. However, once pups have been weaned and are volant, they tend to be more heavily parasitized than females until the colony disperses because they presumably have a naive immunoresponse and less-experience grooming (Christie et al. 2000; Czenze and Broders 2011). Although ectoparasitism tends to increase with colony size, juveniles are usually the most infected hosts at maternity colonies. Adult females still face significant ectoparasite burdens, while ectoparasitism for adult males generally increases throughout swarming as conspecific contact rates increase (Webber et al. 2015a).
Many bat ectoparasites are also vectors for bacterial pathogens (e.g. Bartonella sp., Veikkolainen et al. 2014). Transmission of bacterial or viral pathogens via ectoparasite vectors among bat hosts could be influenced by sociality, especially for highly gregarious females at maternity colonies. In theory, the same principles which regulate host–pathogen dynamics should also apply to host–vector–pathogen dynamics, where the duration and frequency of host-host contact facilitates ectoparasite transmission, which in turn facilitates vector-based pathogen transmission. For instance, recent empirical evidence suggests that wing mites (Spinturnix sp.) can transmit P. destructans propagules among hibernating bats in Europe, a chain of transmission which is facilitated by direct host-host contact (Luçan et al. 2016). We suggest that more future studies quantify links between host sociality, vector ectoparasites, and vector-borne bacterial, viral, and fungal pathogens.
Future Directions and Testable Hypotheses
Progress has been made in identifying relationships between some aspects of sociality and parasitism in bats, but much more empirical data from wild and captive bats of more species is certainly needed. We propose a series of testable hypotheses and possible experimental and observational studies about the role of bat sociality in parasite dynamics. We focus on two aspects of sociality which have been relatively little-studied for bats in the context of parasitism: fission–fusion dynamics and consistent individual differences in behavior or personality.
Fission-Fusion Dynamics
Fission–fusion is the temporary splitting and reforming of colonies, where lack of consensus, or even conflict, result in temporary fission, but the cost of remaining apart is greater than subsequent fusion (Sueur et al. 2011). Within maternity colonies, females exhibit fission–fusion sociality (e.g. M. bechsteinii: Kerth and König 1999; Nyctalus lasiopterus: Popa-Lisseanu et al. 2008) and change roosts every few days but not all members of the group will move together, resulting in variable group size and composition (e.g. Patriquin et al. 2010). Fission–fusion sociality and frequent roost switching by forest-living bats may have evolved as a mechanism to avoid parasites (Reckardt and Kerth 2007; Bartonička and Gaisler 2007), although a trade-off between the costs of parasitism and benefits of remaining loyal to high-quality roosts likely exists. Empirical data have linked fission–fusion behavior to ectoparasite avoidance (e.g. Reckardt and Kerth 2007) and some theoretical modeling also supports this hypothesis (Kashima et al. 2013). However, there are currently no studies investigating microparasite transmission within fission–fusion societies. We hypothesize that the relationship between fission–fusion dynamics and parasite risk and impacts will vary depending on the mode of transmission for a given type of parasite. We expect that for most microparasites and some ectoparasites (e.g. Basilia nana: Reckardt and Kerth 2007) that require direct contact between hosts for transmission, fission–fusion dynamics could serve to disrupt parasite transmission. Bats that display fission–fusion behavior typically switch roosts every 1–2 days (Willis and Brigham 2004) and data from M. bechsteinii suggest that bats are able to detect roosts that have recently been occupied by other bats (Reckardt and Kerth 2007). Thus, when bats switch roosts they appear to select sites that have not been occupied recently which could help them avoid infested roosts. In contrast, we expect that for macroparasites with alternative transmission strategies, such as mobile ectoparasites (e.g. crawling or flying), fission–fusion dynamics are likely to be less effective as a parasite-avoidance strategy. In fact, theory predicts that bats would be more likely to avoid mobile ectoparasites if they remained in a single, large group because of encounter-dilution effects (Côté and Poulin 1995; Rifkin et al. 2012). We suggest studies employing social network analysis of wild bats, combined with estimates of micro- and macroparasite prevalence, and intensity to disentangle relationships between host social behavior, including fission–fusion dynamics, and the ecology of parasite transmission (for review see Godfrey 2013).
Consistent Individual Differences in Behavior
Animal personality refers to consistent individual differences in behavior that are stable within an individual across time and situations (Sih et al. 2004; Wolf and Weissing 2012). Recently, personality has become more widely appreciated as a possible explanatory variable for a number of ecologically relevant traits including juvenile development (Stamps and Groothuis 2010), energy expenditure (Careau et al. 2008) and social structure (Krause et al. 2010; Wolf and Krause 2014). Because personality influences non-random association patterns observed within social groups (Krause et al. 2010), between-individual variation in personality traits are expected to influence parasite transmission patterns within and between groups (Barber and Dingemanse 2010; Kortet et al. 2010). In general, the most active, exploratory or sociable (see Table 5.2 for definitions) individuals are expected to face greater risk of transmitting and/or acquiring parasites (Barber and Dingemanse 2010; Kortet et al. 2010). Despite enormous recent interest in animal personality among behavioral ecologists, evolutionary physiologists, and conservationists (for review see Sih et al. 2004; Réale et al. 2007; Careau et al. 2008; Wolf and Weissing 2012), there are currently only three published studies examining effects of personality on any ecological trait in bats (Menzies et al. 2013; Kilgour and Brigham 2013; Webber et al. 2015b). These studies highlight that individual bats, like many other vertebrates exhibit differences in personality but there is clearly room for much more research on personality in bats, especially in the context of parasite dynamics.
We propose a series of hypotheses that would help to understand relationships between personality and parasite prevalence/intensity and transmission in the context of sociality. As for the implications of fission–fusion dynamics described above, these hypotheses vary for contact-mediated micro- and macroparasites versus mobile macroparasites with host-seeking behavior. First, we hypothesize a relationship between individual sociability and parasitism for contact-mediated parasites. Theory predicts a positive relationship between host sociability and parasite risk, because the most sociable individuals are the most highly connected group-members within the social network and may therefore be exposed to, and themselves transmit, a disproportionate number of infections (Barber and Dingemanse 2010). By contrast, for parasites with active host-seeking behavior, individual activity and exploration may be most important because less active or exploratory individuals may be easier targets for mobile macroparasites. Webber et al. (2015b) identified the expected negative relationship between the prevalence and intensity of fleas (a relatively mobile ectoparasite) and the activity component of personality for female, but not male M. lucifugus, which highlights the potential complexity of these relationships. Personality may have had a larger effect on females because females may be selective with their mating partners, so if less active females mate with fewer males they may retain a higher proportion of fleas, which would otherwise be transmitted during mating (Webber et al. 2015b). We suggest that future studies attempt to disentangle relationships between contact versus mobile parasites and sociability, activity, and exploration components of personality in bats.
Conclusion
Studies of host-parasite dynamics in bats are important from both a conservation and human public health perspective and, as we describe above, the implications of bat social behavior for parasite risk can be dramatic. Based on our review of the literature, we identified social network analysis, epidemiological modeling (often, though not always, parameterized using empirically-derived data), and phylogenetically-informed comparative analyses as the methods most commonly used for quantifying links between sociality and parasitism in bats. In general, these studies suggest complex relationships and studies aimed at fully elucidating these links may require integration of techniques and data collection from multiple disciplines (e.g. behavior, ecology, epidemiology, parasitology, and physiology/immunology).
We also found that, despite its recent emergence, WNS is already among the best-studied pathogens of bats in the context of sociality perhaps, in part because researchers who focus on bats have brought behavioral and ecological expertise to bear on this important conservation problem. The link between host social aggregation and HeV is also well established, again likely because of perceived societal importance, in this case due to the public health implications of a zoonotic pathogen. Relationships between bats and their ectoparasites have also been relatively well studied, perhaps partly because ectoparasites are relatively easy to quantify when bats are in the hand. Ectoparasite studies have identified links between parasite risk, colony size, and fission–fusion dynamics which have broad implications for understanding how sociality affects host–parasite interactions in bats. A few studies have also begun to understand bats in the context of individual animal personality. In general, however, we advocate for more research on bat parasites in the context of socioecology, especially for bat and parasite species which have received little attention to date.
Acknowledgments
We thank Jorge Ortega for inviting us to contribute to this interesting book project and an anonymous reviewer for comments on an earlier version of this chapter. QMRW was funded through a Manitoba Graduate Scholarship (MGS). Funding for CKRW’s research on host–parasite dynamics in bats is provided by a Discovery Grant and a Discovery Accelerator Supplement from the Natural Sciences and Engineering Research Council (NSERC, Canada).
Contributor Information
Jorge Ortega, Email: artibeus2@aol.com.
Quinn M. R. Webber, Email: webber.quinn@gmail.com
Craig K. R. Willis, Email: c.willis@uwinnipeg.ca
References
- Altizer S, Nunn CL, Thrall PH, et al. Social organization and parasite risk in mammals: integrating theory and empirical studies. Annu Rev Ecol Evol S. 2003;34:517–547. doi: 10.1146/annurev.ecolsys.34.030102.151725. [DOI] [Google Scholar]
- Anderson RM, May RM. Population biology of infectious diseases: Part I. Nature. 1979;280:361–367. doi: 10.1038/280361a0. [DOI] [PubMed] [Google Scholar]
- Anderson RM, May RM. Coevolution of hosts and parasites. Parasitology. 1982;85:411–426. doi: 10.1017/S0031182000055360. [DOI] [PubMed] [Google Scholar]
- Anderson RM, May RM. Infectious diseases of humans: dynamics and control. Oxford: University Press; 1992. [Google Scholar]
- Baker ML, Schountz T, Wang LF. Antiviral immune responses of bats: a review. Zoonoses Public Health. 2013;60:104–116. doi: 10.1111/j.1863-2378.2012.01528.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barber I, Dingemanse NJ. Parasitism and the evolutionary ecology of animal personality. Phil Trans R Soc B. 2010;365:4077–4088. doi: 10.1098/rstb.2010.0182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bartonicka T, Gaisler J. Seasonal dynamics in the number of parasitic bugs (Heteroptera, Cimicidae): a possible cause of roost switching in bats (Chiroptera, Vespertilionidae) Parasitol Res. 2007;100:1323–1330. doi: 10.1007/s00436-006-0414-6. [DOI] [PubMed] [Google Scholar]
- Blackwood JC, Streiker DG, Altizer S, Rohani P. Resolving the roles of immunity, pathogenesis and immigration for rabies persistence in vampire bats. P Natl Acad Sci USA. 2013;110:20837–20842. doi: 10.1073/pnas.1308817110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blehert DS, Hicks AC, Behr M, et al. Bat white-nose syndrome: an emerging fungal pathogen? Science. 2009;323:227. doi: 10.1126/science.1163874. [DOI] [PubMed] [Google Scholar]
- Bohn SJ, Turner JM, Warnecke L, Mayo C, McGuire LP, Misra V, Bollinger TK, Willis CKR (in review) Evidence of ‘sickness behaviour’ in bats with white-nose syndrome. Behaviour
- Boratyński JS, Willis CKR, Jefimow M, Wojciechowski MS. Huddling reduces evaporative water loss in torpid Natterer’s bats, Myotis nattereri. Comp Biochem Physiol A. 2015;179:125–132. doi: 10.1016/j.cbpa.2014.09.035. [DOI] [PubMed] [Google Scholar]
- Boyles JG, Willis CKR. Could localized warm areas inside cold caves reduce mortality of hibernating bats affected by white-nose syndrome? Front Ecol Environ. 2010;8:92–98. doi: 10.1890/080187. [DOI] [Google Scholar]
- Brook CE, Dobson AP. Bats as ‘special’ reservoirs for emerging zoonotic pathogens. Trends Microbiol. 2015;23:172–180. doi: 10.1016/j.tim.2014.12.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bruyndonckx N, Henry I, Christe P, Kerth G. Spatio-temporal population genetic structure of the parasitic mite Spinturnix bechsteini is shaped by its own demography and the social system of its bat host. Mol Ecol. 2009;18:3581–3592. doi: 10.1111/j.1365-294X.2009.04299.x. [DOI] [PubMed] [Google Scholar]
- Careau V, Thomas D, Humphries MM, Réale D. Energy metabolism and animal personality. Oikos. 2008;117:641–653. doi: 10.1111/j.0030-1299.2008.16513.x. [DOI] [Google Scholar]
- Carter GG, Wilkinson GS. Food sharing in vampire bats: reciprocal help predicts donations more than relatedness or harassment. Proc R Soc Lond B. 2013;280:201222573. doi: 10.1098/rspb.2012.2573. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cartwright D, Harary F. Structural balance: a generalization of Heider theory. Psychol Rev. 1956;63:277–293. doi: 10.1037/h0046049. [DOI] [PubMed] [Google Scholar]
- Chaverri G. Comparative social network analysis in a leaf-roosting bat. Behav Ecol Sociobiol. 2010;64:1619–1630. doi: 10.1007/s00265-010-0975-3. [DOI] [Google Scholar]
- Christe P, Arlettaz R, Vogel P. Variation in intensity of a parasitic mite (Spinturnix myoti) in relation to the reproductive cycle and immunocompetence of its bat host (Myotis myotis) Ecol Lett. 2000;3:207–212. doi: 10.1046/j.1461-0248.2000.00142.x. [DOI] [Google Scholar]
- Christe P, Glaizot O, Evanno G, Bruyndonckx N, Devevey G, Yannic G, Patthey P, Maeder A, Vogel P, Arlettaz R. Host sex and ectoparasite choice: preference for, and higher survival on female hosts. J Anim Ecol. 2007;76:703–710. doi: 10.1111/j.1365-2656.2007.01255.x. [DOI] [PubMed] [Google Scholar]
- Côté IM, Poulin R. Parasitism and group size in social animals: a meta-analysis. Behav Ecol. 1995;6:159–165. doi: 10.1093/beheco/6.2.159. [DOI] [Google Scholar]
- Craft ME, Cauillaud D. Network models: an underutilized tool in wildlife epidemiology? Interdiscip Perspect Infect Dis. 2011 doi: 10.1155/2011/676949. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Croft DP, James R, Krause J. Exploring animal social networks. Princeton: Princeton University Press; 2008. [Google Scholar]
- Czenze Zenon J., Broders Hugh G. Ectoparasite Community Structure of Two Bats (Myotis lucifugusandM. septentrionalis) from the Maritimes of Canada. Journal of Parasitology Research. 2011;2011:1–9. doi: 10.1155/2011/341535. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Czenze ZJ, Willis CKR. Warming up and shipping out: cues for arousal and emergence in hibernating bats. J Comp Physiol B. 2015;185:575–586. doi: 10.1007/s00360-015-0900-1. [DOI] [PubMed] [Google Scholar]
- Daszak P, Cunningham AA, Hyatt AD. Emerging infectious diseases of wildlife—threats to biodiversity and human health. Science. 2000;287:443–449. doi: 10.1126/science.287.5452.443. [DOI] [PubMed] [Google Scholar]
- Daszak P, Plowright RK, Epstein JH, et al. The emergence of Nipah and Hendra virus: pathogen dynamics across a wildlife-livestock-human continuum. In: Collinge SK, Ray C, et al., editors. Disease ecology: community structure and pathogen dynamics. Oxford: University Press; 2006. pp. 186–201. [Google Scholar]
- de Castro F, Bolker B. Mechanisms of disease-induced extinction. Ecol Lett. 2005;8:117–126. doi: 10.1111/j.1461-0248.2004.00693.x. [DOI] [Google Scholar]
- Dechmann DKN, Kerth G. My home is your castle: roost making is sexually selected in the bat Lophostoma silvicolum. J Mammal. 2008;89:1379–1390. doi: 10.1644/08-MAMM-S-061.1. [DOI] [Google Scholar]
- Drewe JA. Who infects whom? social networks and tuberculosis transmission in wild meerkats. Proc Roy Soc Lond B. 2010;227:633–642. doi: 10.1098/rspb.2009.1775. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Drexler JF, Corman VM, Wegner T, Tateno AF, Zerbinati RM, Gloza-Rausch F, Seebens A, Muller MA, Drosten C. Amplification of emerging viruses in a bat colony. Emerg Infect Dis. 2011;17:449–456. doi: 10.3201/eid1703.100526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eby P. Seasonal movements of grey-headed flying foxes, Pteropus poliocephalus (Chiroptera: Pteropodidae), from two maternity camps in northern New South Wales. Wildlife Res. 1991;18:547–559. doi: 10.1071/WR9910547. [DOI] [Google Scholar]
- Eby P, Richards G, Collins L, Parry-Jones K. The distribution, abundance and vulnerability to population reduction of a nomadic nectarivore, the grey-headed flying-fox Pteropus poliocephalus in New South Wales, during a period of resource concentration. Aust Zoo. 1999;31:240–253. doi: 10.7882/AZ.1999.024. [DOI] [Google Scholar]
- Ellison LE, O’Shea TJ, Neubaum DJ, Bowen RA. Factors influencing movement probabilities of big brown bats (Eptesicus fuscus) in buildings. Ecol Appl. 2007;17:620–627. doi: 10.1890/06-0315. [DOI] [PubMed] [Google Scholar]
- Emlen ST. Benefits, constraints and the evolution of the family. Trends Ecol Evol. 1994;9:282–284. doi: 10.1016/0169-5347(94)90030-2. [DOI] [PubMed] [Google Scholar]
- Encarnaçąo JA, Baulechner D, Becker NI. Seasonal variations of wing mite infestations in male Daubenton’s bats (Myotis daubentonii) in comparison to female and juvenile bats. Acta Chriopterol. 2012;14:153–159. doi: 10.3161/150811012X654367. [DOI] [Google Scholar]
- Ezenwa VO. Host social behavior and parasitic infection: a multifactorial approach. Behav Ecol. 2004;15:446–454. doi: 10.1093/beheco/arh028. [DOI] [Google Scholar]
- Fenton MB. Summer activity of Myotis lucifugus (Chiroptera: Vespertilionidae) at hibernacula in Ontario and Quebec. Can J Zool. 1969;47:597–602. doi: 10.1139/z69-103. [DOI] [Google Scholar]
- Fenton MB. Bats and white-nose syndrome. P Natl Acad Sci U.S.A. 2012;109:6794–6795. doi: 10.1073/pnas.1204793109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fewell JH. Social insect networks. Science. 2003;301:1867. doi: 10.1126/science.1088945. [DOI] [PubMed] [Google Scholar]
- Field H, Crameri G, Kung NY, Wang LF. Ecological aspects of Hendra virus. Curr Top Microbiol. 2012;359:11–23. doi: 10.1007/82_2012_214. [DOI] [PubMed] [Google Scholar]
- Fitze PS, Tschirren B, Richner H. Life history and fitness consequences of ectoparasites. J Anim Ecol. 2004;73:216–226. doi: 10.1111/j.0021-8790.2004.00799.x. [DOI] [Google Scholar]
- Foley J, Clifford D, Castle K, Cryan P, Ostfeld RS. Investigating and managing the rapid emergence of white-nose syndrome, a novel, fatal, infectious disease of hibernating bats. Conserv Biol. 2011 doi: 10.1111/j.1523-1739.2010.01638.x. [DOI] [PubMed] [Google Scholar]
- Fortuna MA, Popa-Lisseanu AG, Ibanez C, Bascompte J. The roosting spatial network of a bird-predator bat. Ecology. 2009;90:934–944. doi: 10.1890/08-0174.1. [DOI] [PubMed] [Google Scholar]
- Frick WF, Pollock JF, Hicks AC, Langwig KE, Reynolds DS, Turner GG, Butchkoski CM, Kunz TH. An emerging disease causes regional population collapse of a common North American bat species. Science. 2010;329:679–682. doi: 10.1126/science.1188594. [DOI] [PubMed] [Google Scholar]
- Frick WF, Puechmaille SJ, Hoyt JR, et al. Disease alters macroecological pattern of North American bats. Global Ecol Biogeogr. 2015 [Google Scholar]
- Frick WF, Puechmaille SJ, Willis CKR. White-nose syndrome in bats. In: Voigt CC, Kingston T, editors. Bats in the anthropocene: conservation of bats in a changing world. London: Springer; 2016. pp. 245–262. [Google Scholar]
- Fuller NW, Reichard JD, Nabhan ML, Fellows SR, Pepin LC, Kunz TH. Free-ranging little brown myotis (Myotis lucifugus) heal from wing damage associated with white-nose syndrome. EcoHealth. 2011;8:154–162. doi: 10.1007/s10393-011-0705-y. [DOI] [PubMed] [Google Scholar]
- Garland T, Jr, Bennet AF, Rezende EL. Phylogenetic approaches in comparative physiology. J Exp Biol. 2005;208:3015–3035. doi: 10.1242/jeb.01745. [DOI] [PubMed] [Google Scholar]
- Gay N, Olival KJ, Bumrungsri S, Siriaroonrat B, Bourgarel M, Morand S. Parasite and viral species richness of Southeast Asian bats: fragmentation of area distribution matters. Int J Parasitol. 2014;3:161–170. doi: 10.1016/j.ijppaw.2014.06.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- George DB, Webb CT, Farnsworth ML, O’Shea TJ, Bowen RA, Smith DL, Stanley TR, Ellison LE, Rupprecht CE. Host and viral ecology determine bat rabies seasonality and maintenance. P Natl Acad Sci U.S.A. 2011;108:10208–10213. doi: 10.1073/pnas.1010875108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gilligan CA, Gubbins S, Simons SA. Analysis and fitting of an SIR model with host response to infection load for a plant disease. Phil Trans R Soc Lond. 1997;352:353–364. doi: 10.1098/rstb.1997.0026. [DOI] [Google Scholar]
- Godde S, Humbert L, Côté SD, Réale D, Whitehead H. Correcting for the impact of gregariousness in social network analyses. Anim Behav. 2013;85:553–558. doi: 10.1016/j.anbehav.2012.12.010. [DOI] [Google Scholar]
- Godfrey SS. Networks and the ecology of parasite transmission: a framework for wildlife parasitology. Int J Parasitol Parasites Wildl. 2013;2:235–245. doi: 10.1016/j.ijppaw.2013.09.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Greer AL, Briggs CL, Collins JP. Testing a key assumption of host-pathogen theory: density and disease transmission. Oikos. 2008;117:1667–1673. doi: 10.1111/j.1600-0706.2008.16783.x. [DOI] [Google Scholar]
- Griffin RH, Nunn CL. Community structure and the spread of infectious disease in primate social networks. Evol Ecol. 2012;26:779–800. doi: 10.1007/s10682-011-9526-2. [DOI] [Google Scholar]
- Hahn MB, Epstein JH, Gurley ES, Islam MS, Luby SP, Daszak P, Patz JA. Roosting behaviour and habitat selection of Pteropus giganteus reveal potential links to Nipah virus epidemiology. J Appl Ecol. 2014;51:376–387. doi: 10.1111/1365-2664.12212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Halpin K, Hyatt AD, Fogarty R, et al. Pteropid bats are confirmed as the reservoir hosts of henipaviruses: a comprehensive experimental study of viral transmission. Am J Trop Med Hyg. 2011;85:946–951. doi: 10.4269/ajtmh.2011.10-0567. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hamede RK, Bashford J, McCallum H, Jones M. Contact networks in a wild tasmanian devil (Sarcophilus harrisii) population: using social network analysis to reveal seasonal variability in social behaviour and its implications for transmission of devil facial tumour disease. Ecol Lett. 2009;12:1147–1157. doi: 10.1111/j.1461-0248.2009.01370.x. [DOI] [PubMed] [Google Scholar]
- Hart BL. Biological basis of the behavior of sick animals. Neurosci Biobehav R. 1988;12:123–137. doi: 10.1016/S0149-7634(88)80004-6. [DOI] [PubMed] [Google Scholar]
- Hayman DTS, Bowen RA, Cryan PM, McCracken GF, O’Shea TJ, Peel AJ, Gilbert A, Webb CT, Wood JLN. Ecology of zoonotic infectious diseases in bats: current knowledge and future directions. Zoonoses Public Health. 2013;60:2–21. doi: 10.1111/zph.12000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoyt JR, Langwig KE, Okoniewski J, Frick WF, Stone WB, Kilpatrick AM. Long-term persistence of Pseudogymnoascus destructans, the causative agent of white-nose syndrome, in the absence of bats. EcoHealth. 2015;12:330–333. doi: 10.1007/s10393-014-0981-4. [DOI] [PubMed] [Google Scholar]
- Hudson PJ, Rizzoli A, Grenfell BT, Heesterbeek H, Dobson AP. The ecology of wildlife diseases. Oxford: University Press; 2002. [Google Scholar]
- Ives AR, Garland T., Jr Phylogenetic logistic regression for binary dependent variables. Syst Biol. 2010;59:9–26. doi: 10.1093/sysbio/syp074. [DOI] [PubMed] [Google Scholar]
- Johnson JS, Kropczynski JN, Lacki MJ, Langlois GD. Social networks of Rafinesque’s big-eared bats (Corynorhinus rafinesquii) in bottomland hardwood forests. J Mammal. 2012;93:1545–1558. doi: 10.1644/12-MAMM-A-097.1. [DOI] [Google Scholar]
- Johnson JS, Kropczynski JN, Lacki MJ. Social network analysis and the study of sociality in bats. Acta Chiropterol. 2013;15:1–17. doi: 10.3161/150811013X667821. [DOI] [Google Scholar]
- Kalcounis MC, Brigham RM. Impact of predation risk on emergence by little brown bats, Myotis lucifugus (Chiroptera: Vespertilionidae), from a maternity colony. Ethology. 1994;98:201–209. doi: 10.1111/j.1439-0310.1994.tb01071.x. [DOI] [Google Scholar]
- Kashima K, Ohtsuki H, Satake A (2013) Fission-fusion bat behaviour as a strategy for balancing the conflicting needs of maximizing information accuracy and minimizing infection risk. 318:101–109 [DOI] [PubMed]
- Keeling MJ, Eames KTD. Networks and epidemic models. J R Soc Interface. 2005;2:295–307. doi: 10.1098/rsif.2005.0051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kerth G. Causes and consequences of sociality in bats. Bioscience. 2008;58:737–746. doi: 10.1641/B580810. [DOI] [Google Scholar]
- Kerth G, König B. Fission, fusion and nonrandom associations in female Bechstein’s bats (Myotis bechsteinii) Behaviour. 1999;136:1187–1202. doi: 10.1163/156853999501711. [DOI] [Google Scholar]
- Kerth G, Perony N, Schweitzer F. Bats are able to maintain long-term social relationships despite the high fission-fusion dynamics of their groups. Proc R Soc Lond B. 2011;278:2761–2767. doi: 10.1098/rspb.2010.2718. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kilgour RJ, Brigham RM. The relationship between behavioural correlates and social influences in the gregarious big brown bat (Eptesicus fuscus) Ethology. 2013;119:1–10. doi: 10.1111/eth.12052. [DOI] [Google Scholar]
- Kortet R, Hedrick AV, Vainikka A. Parasitism, predation and the evolution of animal personalities. Ecol Lett. 2010;13:1449–1458. doi: 10.1111/j.1461-0248.2010.01536.x. [DOI] [PubMed] [Google Scholar]
- Krause J, James R, Croft DP. Personality in the context of social networks. Phil Trans R Soc B. 2010;365:4099–4106. doi: 10.1098/rstb.2010.0216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kunz TH, Lumsden LF. Ecology of cavity and foliage roosting bats. In: Kunz TH, Fenton MB, editors. Bat ecology. 3. Chicago: University Chicago Press; 2003. pp. 3–89. [Google Scholar]
- Kuzmin IV, Rupprecht CE. Bat rabies. In: Wunner WH, Jackson AC, editors. Rabies: scientific basis of the disease and its management. 2. London: Academic Press; 2006. pp. 259–307. [Google Scholar]
- Langwig KE, Frick WF, Bried JT, Hicks AC, Kunz TH, Kilpatrick AM. Sociality, density-dependence and microclimates determine the persistence of populations suffering from a novel fungal disease, white-nose syndrome. Ecol Lett. 2012;15:1050–1057. doi: 10.1111/j.1461-0248.2012.01829.x. [DOI] [PubMed] [Google Scholar]
- Langwig KE, Frick WF, Reynolds R, Parise KL, Drees KP, Hoyt JR, Cheng TL, Kunz TH, Foster JT, Kilpatrick AM. Host and pathogen ecology drive the seasonal dynamics of a fungal disease white-nose syndrome. Proc R Soc Lond B. 2015;282:20142335. doi: 10.1098/rspb.2014.2335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lewis SE. Behavior of Peter’s tent-making bat, Uroderma bilobatum, at maternity roosts in costa rica. J Mammal. 1992;73:541–546. doi: 10.2307/1382020. [DOI] [Google Scholar]
- Lewis SE. Roost fidelity of bats: a review. J Mammal. 1995;76:481–496. doi: 10.2307/1382357. [DOI] [Google Scholar]
- Lewis SE. Low roost-site fidelity in pallid bats: associated factors and effect on group stability. Behav Ecol Sociobiol. 1996;39:335–344. doi: 10.1007/s002650050298. [DOI] [Google Scholar]
- Lindenfors P, Nunn CL, Jones KE, Cunningham AA, Sechrest W, Gittleman JL. Parasite species richness in carnivores: effects of host body mass, latitude, geographical range and population density. Global Ecol Biogeogr. 2007;16:496–509. doi: 10.1111/j.1466-8238.2006.00301.x. [DOI] [Google Scholar]
- Lloyd-Smith JO, Schreiber SJ, Kopp PE, Getz WM. Superspreading and the effect of individual variation on disease emergence. Nature. 2005;438:355–359. doi: 10.1038/nature04153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Loehle C. Social barriers to pathogen transmission in wild animal populations. Ecology. 1995;76:326–335. doi: 10.2307/1941192. [DOI] [Google Scholar]
- Lorch JM, Meteyer CU, Behr MJ, et al. Experimental infection of bats with Geomyces destructans causes white-nose syndrome. Nature. 2011;480:376–378. doi: 10.1038/nature10590. [DOI] [PubMed] [Google Scholar]
- Lourenço SI, Palmeirim JM. Can mite parasitism affect the condition of bat hosts? implications for the social structure of colonial bats. J Zool. 2007;273:161–168. doi: 10.1111/j.1469-7998.2007.00322.x. [DOI] [Google Scholar]
- Luçan RK. Relationships between the parasitic mite Spinturnix andegavinus (Acari: Spinturnicidae) and its bat host, Myotis daubentonii (Chiroptera: Vespertilionidae): seasonal, sex- and age-related variation in infestation and possible impact of the parasite on host condition and roosting behaviour. Folia Parasitol. 2006;53:147–152. doi: 10.14411/fp.2006.019. [DOI] [PubMed] [Google Scholar]
- Luçan RK, Bandouchova H, Bartonička T, Pikula J, Zahradníkova A, Jr, Zukal J, Martínková N. Ectoparasites may serve as vectors for the white-nose syndrome fungus. Parasite Vector. 2016;2016:9–16. doi: 10.1186/s13071-016-1302-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luis AD, Hayman DTS, O’Shea TJ, et al. A comparison of bats and rodents as reservoirs of zoonotic viruses: are bats special? Proc Roy Soc Lond B. 2013;280:1–9. doi: 10.1098/rspb.2012.2753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maganga GD, Bourgarel M, Vallo P, Dallo TD, Ngoagouni C, Drexler JD, Drosten C, Nakoune ER, Leroy EM, Morand S. Bat distribution size or shape as determinant of viral richness in African bats. PLoS ONE. 2014;9:e100172. doi: 10.1371/journal.pone.0100172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin E. Concise medical dictionary. Oxford: University Press; 2010. [Google Scholar]
- May RM. Network structures and the biology of populations. Trends Ecol Evol. 2006;21:394–399. doi: 10.1016/j.tree.2006.03.013. [DOI] [PubMed] [Google Scholar]
- May RM, Anderson RM. Population biology of infectious diseases: part II. Nature. 1979;280:455–461. doi: 10.1038/280455a0. [DOI] [PubMed] [Google Scholar]
- McCallum H, Barlow N, Hone J. How should pathogen transmission be modelled? Trends Ecol Evol. 2001;16:295–300. doi: 10.1016/S0169-5347(01)02144-9. [DOI] [PubMed] [Google Scholar]
- McCallum H, Jones M, Hawkins C, Hamede R, Lachish S, Sinn DL, Beeton N, Lazenby B. Transmission dynamics of tasmanian devil facial tumor disease may lead to disease-induced extinction. Ecology. 2009;90:3379–3392. doi: 10.1890/08-1763.1. [DOI] [PubMed] [Google Scholar]
- McCracken GF, Bradbury JW. Social organization and kinship in the polygynous bat Phyllostomus hastatus. Behav Ecol Sociobiol. 1981;8:11–34. doi: 10.1007/BF00302840. [DOI] [Google Scholar]
- McCracken Gary F., Wilkinson Gerald S. Reproductive Biology of Bats. 2000. Bat Mating Systems; pp. 321–362. [Google Scholar]
- Menzies AK, Timonin ME, McGuire LP, Willis CKR. Personality variation in little brown bats. PLoS ONE. 2013;8:e80230. doi: 10.1371/journal.pone.0080230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Messenger SL, Rupprecht CE, Smith JS. Bats, emerging virus infections, and the rabies paradigm. In: Kunz TH, Fenton MB, editors. Bat ecology. 3. Chicago: University Chicago Press; 2003. pp. 622–666. [Google Scholar]
- Miller-Butterworth CM, Vonhof MJ, Rosenstern J, Turner GG, Russell AL. Genetic structure of little brown bats (Myotis lucifugus) corresponds with spread of white-nose syndrome among hibernacula. J Hered. 2014;105:354–364. doi: 10.1093/jhered/esu012. [DOI] [PubMed] [Google Scholar]
- Moore J. Parasites and the behaviour of animals. Oxford: University Press; 2002. [Google Scholar]
- Morand S, Poulin R. Density, body mass and parasite species richness of terrestrial mammals. Evol Ecol. 1998;12:717–727. doi: 10.1023/A:1006537600093. [DOI] [Google Scholar]
- Norquay KJO, Willis CKR. Hibernation phenology of Myotis lucifugus. J Zool. 2014;294:85–92. doi: 10.1111/jzo.12155. [DOI] [Google Scholar]
- Nunn CL, Altizer S. Infectious diseases in primates. Oxford: University Press; 2006. [Google Scholar]
- Nunn CL, Altizer S, Jones KE, Sechrest W. Comparative tests of parasite species richness in primates. Am Nat. 2003;162:597–614. doi: 10.1086/378721. [DOI] [PubMed] [Google Scholar]
- O’Shea TJ, Neubaum DJ, Neubaum MA, Cryan PM, Ellison LE, Stanley TR, Rupprecht CE, Pape WJ, Bowen RA. Bat ecology and public health surveillance for rabies in an urbanizing region of Colorado. Urban Ecosyst. 2011;14:665–697. doi: 10.1007/s11252-011-0182-7. [DOI] [Google Scholar]
- O’Shea TJ, Cryan PM, Cunningham AA, Fooks AR, Hayman DTS, Luis AD, Peel AJ, Plowright RK, Wood JLN. Bat flight and zoonotic viruses. Emerg Infect Dis. 2014;20:741–745. doi: 10.3201/eid2005.130539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Olival KJ, Hayman DTS. Filoviruses in bats: current knowledge and future directions. Viruses. 2014;6:1759–1788. doi: 10.3390/v6041759. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Otterstatter MC, Thomson JD. Contact networks and transmission of an intestinal pathogen in bumble bee (Bombus impatiens) colonies. Oecologia. 2007;154:411–421. doi: 10.1007/s00442-007-0834-8. [DOI] [PubMed] [Google Scholar]
- Patriquin KJ, Leonard ML, Broders HG, Garroway CJ. Do social networks of female northern long-eared bats vary with reproductive period and age? Behav Ecol Sociobiol. 2010;64:899–913. doi: 10.1007/s00265-010-0905-4. [DOI] [Google Scholar]
- Patterson JEH, Ruckstuhl KE. Parasite infection and host group size: a meta-analytical review. Parasitology. 2013;140:803–813. doi: 10.1017/S0031182012002259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pearce RD, O’Shea TJ, Shankar V, Rupprecht CE. Lack of association between ectoparasite intensities and rabies virus neutralizing antibody seroprevalence in wild big brown bats (Eptesicus fuscus), Fort Collins, Colorado. Vector-Borne Zoonot. 2007;7:489–495. doi: 10.1089/vbz.2007.0572. [DOI] [PubMed] [Google Scholar]
- Perkins SE, Cattadori IM, Tagliapietra V, Rizzoli AP, Hudson PJ. Empirical evidence for key hosts in persistence of tick-borne disease. Int J Parasitol. 2003;33:909–917. doi: 10.1016/S0020-7519(03)00128-0. [DOI] [PubMed] [Google Scholar]
- Plowright RK, Field HE, Smith C, Divljan A, Palmer C, Tabor G, Daszak P, Foley JE. Reproduction and nutritional stress are risk factors for hendra virus infection in little flying foxes (Pteropus scapulatus) Proc R Soc Lond B. 2008;275:861–869. doi: 10.1098/rspb.2007.1260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Plowright RK, Foley P, Field HE, Dobson AP, Foley JE, Eby P, Daszak P. Urban habituation, ecological connectivity and epidemic dampening: the emergence of Hendra virus from fyling foxes (Pteropus spp.) Proc R Soc Lond B. 2011;278:3703–3712. doi: 10.1098/rspb.2011.0522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Plowright RK, Eby P, Hudson PJ, et al. Ecological dynamics of emerging bat virus spillover. Proc R Soc Lond B. 2015;282:1–9. doi: 10.1098/rspb.2014.2124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Popa-Lisseanu AG, Bontadina F, Mora O, Ibañez C. Highly structured fission-fusion societies in an aerial-hawking carnivorous bat. Anim Behav. 2008;75:471–482. doi: 10.1016/j.anbehav.2007.05.011. [DOI] [Google Scholar]
- Postawa T, Szubert-Kruszynska A. Is parasite load dependent on host aggregation size? The case of the greater mouse-eared bat Myotis myotis (Mammalia: Chiroptera) and its parasitic mite Spinturnix myoti (Acari: Gamasida) Parasitol Res. 2014;113:1803–1811. doi: 10.1007/s00436-014-3826-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Presley SJ. Interspecific aggregation of ectoparasites on bats: importance of hosts as habitats supersedes interspecific interactions. Oikos. 2011;120:832–841. doi: 10.1111/j.1600-0706.2010.19199.x. [DOI] [Google Scholar]
- Presley SJ, Willig MR. Intraspecific patterns of ectoparasite abundances on Paraguayan bats: effects of host sex and body size. J Trop Ecol. 2008;24:75–83. doi: 10.1017/S0266467407004683. [DOI] [Google Scholar]
- Puechmaille SJ, Wibbelt G, Korn V, et al. Pan-European distribution of white-nose syndrome fungus (Geomyces destructans) not associated with mass mortality. PLoS ONE. 2011;6:e19167. doi: 10.1371/journal.pone.0019167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Réale D, Reader SM, Sol D, McDougall PT, Dingemanse NJ. Integrating animal temperament with ecology and evolution. Biol Rev. 2007;82:291–318. doi: 10.1111/j.1469-185X.2007.00010.x. [DOI] [PubMed] [Google Scholar]
- Reckardt K, Kerth G. The reproductive success of the parasite bat fly Basilia nana (Diptera: Nycteribiidae) is affected by the low roost fidelity of its host, the Bechstein’s bat (Myotis bechsteinii) Parasitol Res. 2006;98:237–243. doi: 10.1007/s00436-005-0051-5. [DOI] [PubMed] [Google Scholar]
- Reckardt K, Kerth G. Roost selection and roost switching of female Bechstein’s bats (Myotis bechsteinii) as a strategy of parasite avoidance. Oecologia. 2007;154:581–588. doi: 10.1007/s00442-007-0843-7. [DOI] [PubMed] [Google Scholar]
- Reckardt K, Kerth G. Does the mode of transmission between hosts affect the host choice strategies of parasites? implications from a field study of bat fly and wing mite infestation of Bechstein’s bats. Oikos. 2009;118:183–190. doi: 10.1111/j.1600-0706.2008.16950.x. [DOI] [Google Scholar]
- Reeder DM, Frank CL, Turner GG, et al. Frequent arousal from hibernation linked to severity of infection and mortality in bats with white-nose syndrome. PLoS ONE. 2012;7:e3892. doi: 10.1371/journal.pone.0038920. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rhodes M, Wardell-Johnson GW, Rhodes MP, Raymond B. Applying network analysis to the conservation of habitat trees in urban environments: a case study from Brisbane, Australia. Conserv Biol. 2006;3:861–870. doi: 10.1111/j.1523-1739.2006.00415.x. [DOI] [PubMed] [Google Scholar]
- Rifkin JL, Nunn CL, Garamszegi LZ. Do animals living in larger groups experience greater parasitism? a meta-analysis. Am Nat. 2012;180:70–82. doi: 10.1086/666081. [DOI] [PubMed] [Google Scholar]
- Rosenblum EB, Voyles J, Poorten TJ, Stajich JE. The deadly chytrid fungus: a story of an emerging pathogen. PLoS Pathog. 2010;6:e1000550. doi: 10.1371/journal.ppat.1000550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rupprecht CE, Turmelle A, Kuzmin IV. A perspective on lyssavirus emergence and perpetuation. Curr Opin Vir. 2011;1:622–670. doi: 10.1016/j.coviro.2011.10.014. [DOI] [PubMed] [Google Scholar]
- Sade DS, Dow M. Primate social networks. In: Wasserman S, Galaskiewicz J, editors. Advances in social network analysis: research in the social and behavioral sciences. California Sage: Thousand Oaks; 1994. pp. 152–166. [Google Scholar]
- Serra-Cobo J, Lopez-Roig M, Segui M, Sanchez LP, Nadal J, Borras M, Lavenir R, Bourhy H. Ecological factors associated with European bat Lyssavirus seroprevalence in Spanish bats. PLoS ONE. 2013;8:e64467. doi: 10.1371/journal.pone.0064467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sih A, Bell AM, Chadwick-Johnson J, Ziemba RE. Behavioural syndromes: an integrative overview. Q Rev Biol. 2004;79:241–277. doi: 10.1086/422893. [DOI] [PubMed] [Google Scholar]
- Silvis A, Ford WM, Britzke ER, Johnson JB. Association, roost use and simulated disruption of Myotis septentrionalis maternity colonies. Behav Process. 2014;103:283–290. doi: 10.1016/j.beproc.2014.01.016. [DOI] [PubMed] [Google Scholar]
- Stamps J, Groothuis TGG. The development of animal personality: relevance, concepts and perspectives. Biol Rev. 2010;85:301–325. doi: 10.1111/j.1469-185X.2009.00103.x. [DOI] [PubMed] [Google Scholar]
- Stanko M, Miklisová D, de Bellocq JG, Morand S. Mammal diversity and patterns of ectoparasite species richness and abundance. Oecologia. 2002;131:289–295. doi: 10.1007/s00442-002-0889-5. [DOI] [PubMed] [Google Scholar]
- Stawski C, Willis CKR, Geiser F. The importance of temporal heterothermy in bats. J Zool. 2014;292:86–100. doi: 10.1111/jzo.12105. [DOI] [Google Scholar]
- Streicker DG, Recuenco S, Valderrama W, et al. Ecological and anthropogenic drivers of rabies exposure in vampire bats: implications for transmission and control. Proc R Soc Lond B. 2012;279:3384–3392. doi: 10.1098/rspb.2012.0538. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sueur C, King AJ, Conradt L, Kerth G, Lusseau D, Mettke-Hofmann C, Schaffner CM, Williams L, Zinner D, Aureli F. Collective decision-making and fission-fusion dynamics: a conceptual framework. Oikos. 2011;120:1608–1617. doi: 10.1111/j.1600-0706.2011.19685.x. [DOI] [Google Scholar]
- Tompkins DM, Dunn AM, Smith MJ, Telfer S. Wildlife disease: from individuals to ecosystems. J Anim Ecol. 2011;80:19–38. doi: 10.1111/j.1365-2656.2010.01742.x. [DOI] [PubMed] [Google Scholar]
- Turmelle AS, Olival KJ. Correlates of viral richness in bats (Order Chiroptera) EcoHealth. 2009;6:522–539. doi: 10.1007/s10393-009-0263-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Shaik J, Kerth G, Bruyndinckx N, Christe P. The effect of host social system on parasite population genetic structure: comparative population genetics of two ectoparasitic mite and their bat hosts. BMC Evol Biol. 2014;14:1–15. doi: 10.1186/1471-2148-14-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Veikkolainen V, Vesterinen EJ, Lilley TM, Pulliainen AT. Bats as reservoir hosts of human bacterial pathogen, Bartonella mayotimonensis. Emerg Inf Dis. 2014;6:960–967. doi: 10.3201/eid2006.130956. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Verant ML, Boyles JG, Waldrep W, Jr, Wibbelt G, Blehert DS. Temperature -dependent growth of Geomyces destructans, the fungus that causes bat white-nose syndrome. PLoS ONE. 2012;7:e46280. doi: 10.1371/journal.pone.0046280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vertant ML, Meteyer CU, Speakman JR, Cryan PM, Lorch JM, Blehert DS. White-nose syndrome initiates a cascade of physiologic disturbances in the hibernating bat host. BMC Physiol. 2014 doi: 10.1186/s12899-014-0010-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vitone ND, Altizer S, Nunn CL. Body size, diet and sociality influence the species richness of parasitic worms in anthropoid primates. Evol Ecol Res. 2004;6:183–199. [Google Scholar]
- Voyles J, Young S, Berger L, et al. Pathogenesis of chytridiomycosis, a cause of catastrophic amphibian declines. Science. 2009;326:582–585. doi: 10.1126/science.1176765. [DOI] [PubMed] [Google Scholar]
- Warnecke L, Turner JM, Bollinger TK, Lorch JM, Misra V, Cryan PM, Wibbelt G, Blehert DS, Willis CKR. Inoculation of bats with European Geomyces destructans supports the novel pathogen hypothesis for the origin of white-nose syndrome. P Natl Acad Sci USA. 2012;109:6999–7003. doi: 10.1073/pnas.1200374109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wasserman S, Faust K. Social network analysis: methods and applications. New York: Cambridge University Press; 1994. [Google Scholar]
- Webber QMR, Czenze ZJ, Willis CKR. Host demographic predicts ectoparasite dynamics for a colonial host during pre-hibernation mating. Parasitology. 2015;142:1260–1269. doi: 10.1017/S0031182015000542. [DOI] [PubMed] [Google Scholar]
- Webber QMR, McGuire LP, Smith SB, Willis CKR. Host behaviour, age and sex correlate with ectoparasite prevalence and intensity in a colonial mammal, the little brown bat. Behaviour. 2015;152:83–105. doi: 10.1163/1568539X-00003233. [DOI] [Google Scholar]
- Webber Quinn M. R., Brigham R. Mark, Park Andrew D., Gillam Erin H., O’Shea Thomas J., Willis Craig K. R. Social network characteristics and predicted pathogen transmission in summer colonies of female big brown bats (Eptesicus fuscus) Behavioral Ecology and Sociobiology. 2016;70(5):701–712. [Google Scholar]
- Wey Tina, Blumstein Daniel T., Shen Weiwei, Jordán Ferenc. Social network analysis of animal behaviour: a promising tool for the study of sociality. Animal Behaviour. 2008;75(2):333–344. [Google Scholar]
- Whitehead H. Analyzing animal societies: quantitative methods for vertebrate social analysis. Chicago: The University of Chicago Press; 2008. [Google Scholar]
- Wilcox Alana, Willis Craig K. R. Energetic benefits of enhanced summer roosting habitat for little brown bats (Myotis lucifugus) recovering from white-nose syndrome. Conservation Physiology. 2016;4(1):cov070. doi: 10.1093/conphys/cov070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilcox A, Warnecke L, Turner JM, McGuire LP, Jameson JW, Misra V, Bollinger TC, Willis CKR. Behaviour of hibernating little brown bats experimentally inoculated with the pathogen that causes white-nose syndrome. Anim Behav. 2014;88:157–164. doi: 10.1016/j.anbehav.2013.11.026. [DOI] [Google Scholar]
- Wilder AP, Frick WF, Langwig KE, Kunz TH. Risk factors associated with mortality from white-nose syndrome among hibernating bat colonies. Biol Lett. 2011;7:950–953. doi: 10.1098/rsbl.2011.0355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilkinson GS. Reciprocal altruism in bats and other mammals. Ethol Sociobiol. 1987;9:85–100. doi: 10.1016/0162-3095(88)90015-5. [DOI] [Google Scholar]
- Willis CKR. Conservation physiology and conservation pathogens: white-nose syndrome and integrative biology for host-pathogen systems. Integr Comp Biol. 2015;55:631–641. doi: 10.1093/icb/icv099. [DOI] [PubMed] [Google Scholar]
- Willis CKR, Brigham RM. Roost switching, roost sharing and social cohesion: forest-dwelling big brown bats, Eptesicus fuscus, conform to the fission-fusion model. Anim Behav. 2004;68:495–505. doi: 10.1016/j.anbehav.2003.08.028. [DOI] [Google Scholar]
- Willis CKR, Brigham RM. Social thermoregulation exerts more influence than microclimate on forest roost preferences by a cavity-dwelling bat. Behav Ecol Sociobiol. 2007;62:97–108. doi: 10.1007/s00265-007-0442-y. [DOI] [Google Scholar]
- Willis CKR, Kolar KA, Karst AL, Kalcounis-Rueppell MC, Brigham RM. Medium- and long-term reuse of trembling aspen cavities as roosts by big brown bats (Eptesicus fuscus) Acta Chiropterol. 2003;5:85–90. doi: 10.3161/001.005.0107. [DOI] [Google Scholar]
- Willis CKR, Menzies AK, Boyles JG, Wojciechowski MS. Evaporative water loss is a plausible explanation for mortality of bats from white-nose syndrome. Integr Comp Biol. 2011;51:364–373. doi: 10.1093/icb/icr076. [DOI] [PubMed] [Google Scholar]
- Wilson EO. Sociobiology: the new synthesis. Cambridge, Massachusetts: Belknap Press; 1975. [Google Scholar]
- Wolf M, Krause J. Why personality differences matter for social functioning and social structure. Trend Ecol Evol. 2014;29:306–308. doi: 10.1016/j.tree.2014.03.008. [DOI] [PubMed] [Google Scholar]
- Wolf M, Weissing FJ. Animal personalities: consequences for ecology and evolution. Trends Ecol Evol. 2012;27:452–461. doi: 10.1016/j.tree.2012.05.001. [DOI] [PubMed] [Google Scholar]
- Zhang L, Parsons S, Daszak P, Wei L, Zhu G, Zhang S. Variation in the abundance of ectoparasitic mites of flat-headed bats. J Mammal. 2010;91:136–143. doi: 10.1644/08-MAMM-A-306R2.1.. [DOI] [Google Scholar]
- Zuk M, McKean KA. Sex differences in parasite infections: patterns and processes. Int J Parasitol. 1996;26:1009–1024. doi: 10.1016/S0020-7519(96)80001-4. [DOI] [PubMed] [Google Scholar]
- Zukal J, Bandouchova H, Brichta J, et al. White-nose syndrome without borders: Pseudogymnoascus destructans infection tolerated in Europe and Palearctic Asia but not in North America. Sci Rep. 2016;6:19829. doi: 10.1038/srep19829. [DOI] [PMC free article] [PubMed] [Google Scholar]