

Abstract
Herein we describe the ability of the permissive glycosyltransferase (GT) OleD Loki to convert a diverse set of > 15 histone deacetylase (HDAC) inhibitors (HDACis) into their corresponding hydroxamate glycosyl esters. Representative glycosyl esters were subsequently evaluated in assays for cancer cell line cytotoxicity, chemical and enzymatic stability, and axolotl embryo tail regeneration. Computational substrate docking models were predictive of enzyme-catalyzed turnover and suggest certain HDACis may form unproductive, potentially inhibitory, complexes with GTs.
Keywords: glucosylation, glycosyltransferase, HDAC, histone deacetylase
Graphical Abstract
HDAC inhibitor sweetener: Hydroxamate-based HDAC inhibitors (HDACis) were identified as glycosyltransferase (OleD Loki) substrates, and corresponding hydroxamate glycosyl ester products were evaluated in assays for cancer cell line cytotoxicity, chemical/enzymatic stability, and axolotl (salamander) embryo tail regeneration. Computational docking models suggest certain HDACis to form unproductive, potentially inhibitory, complexes with glycosyltransferases.

Sugar nucleotide-dependent glycosyltransferases (Leloir GTs)[1] catalyze the regio/stereospecific transfer of sugars from activated sugar-nucleotide donors to hydroxy groups,[2] amines,[3] thiols,[4] and activated carbon nucleophiles.[5] GTs are prevalent in nature and contribute to diverse cellular functions including, but not limited to, cellular signaling, molecular recognition, energy/metabolite storage, and drug resistance/detoxification. These proficient biocatalysts have also been used for glycoside synthesis as exemplified by applications of the permissive microbial detoxifying GTs YjiC[6] and OleD.[7] Within this context, OleD acceptor and sugar nucleotide donor permissivity has been further enhanced via directed evolution and structure-based approaches.[7g,j,k] An OleD-based transglycosylation strategy has also been developed[7f,i] that exploits GT-catalyzed reaction reversibility, and the ability of enhanced OleD mutants (OleD ASP, TDP16 and Loki)[7a–f,i–k] to efficiently use simple colorimetric or fluorescent substrates. This transglycosylation platform also enabled a plate-based screen to identify new OleD substrates and improved OleD-catalyzed syntheses via shifting the reaction equilibria toward desired glycoside product.[7a–f]
One such recent transglycosylation screen identified the histone deacetylase inhibitor (HDACi) trichostatin A as a new OleD Loki substrate and subsequently demonstrated the efficient OleD-catalyzed synthesis of the corresponding glucopyranosyl hydroxamate trichostatin C (the only naturally occurring hydroxamate glycoside isolated to date, produced by Streptomyces platensis No. 145 and Streptomyces strain Y-50).[7a,8] Although HDACis have broad clinical utility, their corresponding glycosides have not been extensively studied. To further probe the synthetic utility of OleD, herein we describe the evaluation of 21 structurally diverse hydroxamate-based HDACis as potential OleD substrates. This study revealed OleD Loki to turnover 17 of the hydroxamates tested to their corresponding hydroxamate glycosyl esters. Subsequent bioactivity studies revealed hydroxamate glucosylation to decrease cytotoxicity by 20- to > 3000-fold. The corresponding hydroxamates were also found to be resistant to glycosidase-catalyzed hydrolysis and afford moderate acid and base stability. Cumulatively, this study advances a new OleD catalytic function and the first fundamental stability and bioactivity studies for hydroxamate glycosides.
Using 2-chloro-4-nitrophenyl β-d-glucoside (ClNP β-d-Glc) and the OleD variant Loki,[7a–e] 21 representative hydroxamate-based HDACis were evaluated as putative substrates via a simple colorimetric transglycosylation screen under the control of a catalytic amount of UDP (Figure 1 A). Assays (1 mm hydroxamate, 2 mm ClNP β-d-Glc, 0.1 mm UDP, 25 mm Tris pH 8.0, 5 mm MgCl2, 0.25 μm OleD Loki, 30 μL total volume, 30°C, 1 h, A410) were conducted in triplicate in 384 well plates. Each plate also contained a positive (4-methylumbeliferone; 4-MeUmb)[7a–g,i–k] and negative (DMSO) control. Seventeen preliminary primary hits were identified (ΔA410 > 3 standard deviations above the negative control) in this first-pass screen (Figure 1 B). Crude reactions identified as preliminary primary hits were subsequently analyzed by LC–MS. For all 17 primary hits, the observation of a single major glucoside product by LC–MS served as key validation of the colorimetric screen (Figure S1 and Table S1 in the Supporting Information). Representatives 1–5 (Figure 2) were subsequently selected as models for scale-up and structure elucidation based on turnover and commercial availability.
Figure 1.
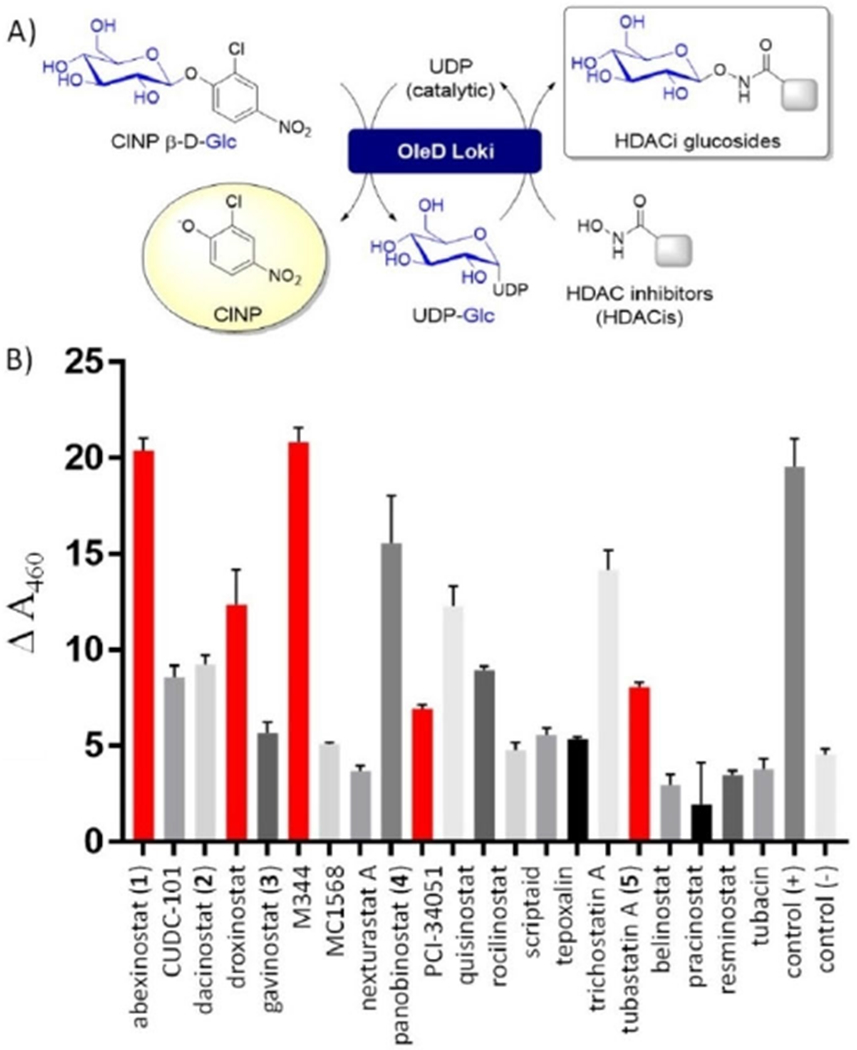
A) Schematic of the colorimetric screen. B) Reaction progress as monitored via ΔA410 using the standard assay format (see panel A; 1 mm hydroxamate, 2 mm CINP β-d-Glc, 0.1 mm UDP, 25 mm Tris pH 8.0, 5 mm MgCl2, 0.25 μm OleD Loki, 30 μL total volume, 30 °C, 1 h). Vehicle alone (no acceptor) served as the negative control and a well-characterized OleD substrate (4-MeUmb) as the positive control. Assays were conducted in triplicate. GT, OleD Loki; ClNP β-d-Glc, 2-chloro-4-nitrophenol β-d-glucose; UDP, uridine diphosphate. The bars in red represent the compounds selected for scale-up and characterization. Hydroxamate structures are illustrated in Figure 2.
Figure 2.

HDACis tested as putative OleD Loki substrates. Compounds in black were identified as hits in the colorimetric screen and subsequently confirmed by LC–MS. Percent conversion (in parentheses) was determined via HPLC peak integration. Compounds in grey lacked turnover.
Scale-up reactions were each accomplished in a total volume of 30 mL (1 mm 1–5, 20 mm Tris·HCl, pH 8.0, 5 mm MgCl2, 2 mm CINP-β-d-glucose, 0.1 mm UDP, 1 μm OleD Loki, 30 °C, 12–24 h) with reaction progress monitored via ΔA410. Upon completion of the reaction, products and residual reactants were captured by solid phase extraction and the resulting mixture purified by semipreparative reversed-phase HPLC (Supporting Information Method B). Glycoside products were collected and subjected to HR-ESI-MS to establish the molecular formula for compounds 1a–5a (Figure 3) as 1a, C27H33N3O10 [m/z 560.2240 (M + H)+]; 2 a, C28H35N3O8 [m/z 542.2489 (M + H)+]; 3a, C30H37N3O9 [m/z 584.2601 (M + H)+]; 4a, C27H33N3O7 [m/z 512.2392 (M + H)+]; and 5a, C26H31N3O7 [m/z 498.2237 (M + H)+], respectively (Table S1). Comparison of the 1a–5a 1D and 2D NMR to that of the corresponding parental hydroxamates (1–5; (Figures S6–S50) revealed 1a–5a signatures consistent atypical glucosides and a lack of the 1–5 hydroxamate hydroxy 1H NMR resonance (δH = 8.5). Evidence of a 1 a–5 a β-O-glucoside derived from the key anomeric 1H (near δH = 4.5, J=6.6–8 Hz) and 13C (δC = 102–106) resonances (Tables S2–S6, Figure S4) typical of β-O-glucosides.[9] As comparators, typical anomeric carbon 13C resonances for C- and N-glycosides range from δC = 70–85[10] and 85–95,[7a,11] respectively. It is also important to note that 1 and 3–5 contain only a single accessible nucleophilic OH (that of the hydroxamate). Consistent with a glycosidic bond comprised of two heteroatoms,[12] HMBC correlations between the sugar and aglycone structure were not observed for 1 a–5 a. Cumulatively, these data provide strong support for the proposed structures and are also consistent with the previously characterized naturally occurring hydroxamate glycoside trichostatin C.[7a,8]
Figure 3.

Representative hydroxamate glycosyl ethers selected for scale-up and further study.
The original trichostatin C structure elucidation studies revealed susceptibility of the naturally occurring hydroxamate glucoside to strong acid (3 n HCl or 40% methanolic HCl).[8b] However, to the best of our knowledge, the lability of hydroxamate glycosides has not been extensively studied. Thus, model hydroxamate glucosides 3 a and 4 a were further evaluated for pH stability and susceptibility to glucosidase-catalyzed hydrolysis. In this analysis, glucosides 3 a, 4 a and commercially available control 4-methylumbelliferyl-glucopyranosides were stable at pH 5, 7.5 and 10 (≤ 2% degradation over 30 days, room temperature; Figure S2). Consistent with the prior trichostatin C precedent, all glucosides were unstable to concentrated H2SO4 (≈50% glucoside hydrolysis within 5 min based on thin layer chromatography, data not shown). This limited study suggested similar pH liabilities among hydroxamate and umbelliferone glucosides. Hydroxamate glucosides 3 a and 4 a were also resistant to α- and β-glucosidase-catalyzed hydrolysis while corresponding controls 4-methylumbelliferyl α-d-glucopyranoside and 4-methylumbelliferyl β-d-glucopyranoside were rapidly hydrolyzed by α- or β-glucosidase, respectively (Figure S3). As glucosidases are typically permissive to substantial aglycon structural diversity,[13] this result may suggest unique electronic and/or steric features contribute to the glycosidase resistance of hydroxamate glycosides.
HDACs catalyze histone lysine deacetylation and function as critical cellular epigenetic modifiers.[14] The agents illustrated in Figure 2 are prototypical HDACis and include three clinically approved anticancer drugs (belinostat, panobinostat and vorinostat). Hydroxamates function as reversible inhibitors by chelating the key HDAC active-site Zn2+ and display potent cancer cell line antiproliferative activities. Yet, while trichostatin A and its hydroxamate glycoside trichostatin C were both reported to increase histone H4 acetylation in B cells and induce erythroleukemia differentiation,[15] the impact of hydroxamate glycosylation on biological activity has not been extensively studied. To address this, compounds 1a–5a and their parental hydroxamates 1–5 were evaluated in standard cancer cell line cytotoxicity assays against both human colorectal (HCT116) and non-small-cell lung (A549) cancer cell lines (Figure 4). Similar to the previously observed decrease in potency invoked via trichostatin A glycosylation, glucosylation of 1–5 led to a 20- to > 3000-fold decrease in potency. Glucoside potency trends mirrored that of the parental HDACis with the glucoside of the pan-HDACi panobinostat (4 A549 IC50 0.8 nm; HCT116 IC50 1.8 nm) identified as the most cytotoxic hydroxamate glycosyl ester (4a A549 IC50 > 1.0 μm; HCT116 IC50 > 1.9 μm).
Figure 4.

Comparative cytotoxicity of representative hydroxamates and hydroxamate glycosyl ethers toward A) human colorectal (HTC116) and B) non-small cell lung (A549 cells) cancer cell lines.
Using a Mexican axolotl (Ambystoma mexicanum) embryo tail regeneration (ETR) assay,[16] HDACis have also been identified as potent inhibitors of tail regeneration. Additional subsequent HDACi-based chemical genetic and microarray studies highlighted the importance of HDAC activity at the time of tail amputation to regulate the initial transcriptional response to injury and regeneration in the axolotl model.[16e] To investigate the impact of hydroxamate glycosyl esters within this context, compounds 1a–5a and their parental hydroxamates 1–5 were evaluated in the ETR assay.[15] Tail-amputated embryos were incubated in microtiter plates in the absence (vehicle control, DMSO) or presence of 10 μm test agent (1–5, 1a–5a) and imaged on day 1 (pre-treatment) and day 7. Consistent with the prior study, parental hydroxamates inhibited tail regeneration (Figure S5). In contrast, hydroxamate glycosides were inactive at the concentration tested, potentially due to reduced potency (consistent with Figure 4) and/or in vivo exposure.
Computational substrate docking models (Figure 5) are also consistent with empirically determined turnover (or lack thereof). Specifically, these models highlight that the representative models for high (abexinostat, Figure 5 A), moderate (tepoxalin, Figure 5 B), and low (MC1568, Figure 5 C) OleD turnover all orient the hydroxamate within close proximity to the key OleD catalytic H19-D110 acid-base pair. Conversely, the predicted high affinity binding mode of a ligand that lacked turnover with OleD (tubacin) revealed a catalytically unproductive conformation (Figure 5D). This computational model may implicate tubacin and/or related pharmacophores as a potential new starting point for GT inhibitor discovery.
Figure 5.

The predicted binding OleD binding models for A) abexinostat (high turnover, Figures 1 and 2, 1), B) tepoxalin (moderate turnover, Figures 1 and 2), C) MC1568 (low turnover, Figures 1 and 2) and D) tubacin (no turnover, Figures 1 and 2) based on the wild-type OleD-erythromycin ligand-bound crystal structure (PDB ID: 2IYF). Yellow dashed lines reflect putative key binding/catalytic contacts (distances in Å), including that of the ligand hydroxamate with the putative OleD active site base (His19 side chain of the active-site H19–D110 acid–base pair).
In summary, this study expands on the prior discovery of trichostatin A as an OleD Loki substrate.[7a] Consistent with the prior comparison of the catalytic competencies for trichostatin A (kcat/KM = 1.4×104 μm−1s−1) to the parental OleD Loki acceptor 4-methylumbelliferone (kcat/KM = 2.2×104 μm−1 s−1), the current study highlights OleD Loki as an efficient and permissive biocatalyst for HDACi glucosylation as a basis for exploring glycoconjugate tumor targeting and/or prodrug strategies.[17] Hydroxamate glycosylation is also expected to circumvent the Lossen rearrangement to a highly reactive isocyanate, a reaction known to contribute to nonspecific alkylation and corresponding off-target toxicity observed by HDACis.[14f] Most hydroxamates also suffer from rapid metabolism and clearance, where UDP-glucuronosyltransferase (UGT)-catalyzed hydroxamate glucuronidation plays a major role.[18] Within this context, the OleD Loki platform may also serve as a potential screen to facilitate the discovery of hydroxamate analogues less susceptible to enzymatic glycosylation. These cumulative factors suggest OleD Loki may add to the repertoire of medicinal chemistry tools to advance HDACis for cancer,[19] immunotherapy,[20] neurological disorders,[21] and/or infectious disease.[22]
Supplementary Material
Acknowledgements
This work was supported in part by US National Institutes of Health (NIH) grants R37 AI52218 (J.S.T.) and R24 OD21479 (S.R.V. and J.S.T.), the National Center for Advancing Translational Sciences (UL1 TR000117 and UL1 TR001998), and the University of Kentucky College of Pharmacy. We thank the College of Pharmacy NMR Center (University of Kentucky) for NMR support.
Footnotes
Conflict of Interest
J.S.T. is a co-founder of Centrose (Madison, WI, USA).
References
- [1].a) Elshahawi SI, Shaaban KA, Kharel MK, Thorson JS, Chem. Soc. Rev 2015, 44, 7591–7697; [DOI] [PMC free article] [PubMed] [Google Scholar]; b) Gloster TM, Curr. Opin. Struct. Biol 2014, 28, 131–141; [DOI] [PMC free article] [PubMed] [Google Scholar]; c) Breton C, Fournel-Gigleux S, Palcic MM, Curr. Opin. Struct. Biol 2012, 22, 540–549; [DOI] [PubMed] [Google Scholar]; d) Singh S, Phillips GN Jr., Thorson JS, Nat. Prod. Rep 2012, 29, 1201–1237; [DOI] [PMC free article] [PubMed] [Google Scholar]; e) Chang A, Singh S, Phillips GN Jr., Thorson JS, Curr. Opin. Biotechnol 2011, 22, 800–808; [DOI] [PMC free article] [PubMed] [Google Scholar]; f) Gantt RW, Peltier-Pain P, Thorson JS, Nat. Prod. Rep 2011, 28, 1811–1853; [DOI] [PubMed] [Google Scholar]; g) Palcic MM, Curr. Opin. Chem. Biol 2011, 15, 226–233; [DOI] [PubMed] [Google Scholar]; h) Roychoudhury R, Pohl NL, Curr. Opin. Chem. Biol 2010, 14, 168–173; [DOI] [PubMed] [Google Scholar]; i) Lairson L, Henrissat B, Davies G, Withers S, Annu. Rev. Biochem 2008, 77, 521–555; [DOI] [PubMed] [Google Scholar]; j) Bowles D, Lim E, Poppenberger B, Vaistij F, Annu. Rev. Plant Biol 2006, 57, 567–597. [DOI] [PubMed] [Google Scholar]
- [2].a) Wu CZ, Jang JH, Woo M, Ahn JS, Kim JS, Hong Y, Appl. Environ. Microbiol 2012, 78, 7680–7686; [DOI] [PMC free article] [PubMed] [Google Scholar]; b) Ahn B, Kim B, Jeon Y, Lee E, Lim Y, Ahn J, J. Microbiol. Biotechnol 2009, 19, 387–390; [DOI] [PubMed] [Google Scholar]; c) Méndez C, Salas J, Ernst Schering Res. Found. Workshop 2005, 51, 127–146; [DOI] [PubMed] [Google Scholar]; d) Lu W, Oberthur M, Leimkuhler C, Tao J, Kahne D, Walsh C, Proc. Natl. Acad. Sci. USA 2004, 101, 4390–4395; [DOI] [PMC free article] [PubMed] [Google Scholar]; e) Rodriguez L, Aguirrezabalaga I, Allende N, Brana A, Mendez C, Salas J, Chem. Biol 2002, 9, 721–729. [DOI] [PubMed] [Google Scholar]
- [3].a) Guo Z, Li J, Qin H, Wang M, Lv X, Li X, Chen Y, Angew. Chem. Int. Ed 2015, 54, 5175–5178; [DOI] [PubMed] [Google Scholar]; Angew. Chem 2015, 127, 5264–5267; [Google Scholar]; b) Gawthorne JA, Tan NY, Bailey UM, Davis MR, Wong LW, Naidu R, Fox KL, Jennings MP, Schulz BL, Biochem. Biophys. Res. Commun 2014, 445, 633–638; [DOI] [PubMed] [Google Scholar]; c) Naegeli A, Michaud G, Schubert M, Lin CW, Lizak C, Darbre T, Reymond JL, Aebi M, J. Biol. Chem 2014, 289, 24521–24532; [DOI] [PMC free article] [PubMed] [Google Scholar]; d) Magarvey NA, Haltli B, He M, Greenstein M, Hucul JA, Antimicrob. Agents Chemother 2006, 50, 2167–2177; [DOI] [PMC free article] [PubMed] [Google Scholar]; e) Gao Q, Zhang C, Blanchard S, Thorson JS, Chem. Biol 2006, 13, 733–743; [DOI] [PubMed] [Google Scholar]; f) Sánchez C, Méndez C, Salas JA, Nat. Prod. Rep 2006, 23, 1007–1045; [DOI] [PubMed] [Google Scholar]; g) Zhang C, Albermann C, Fu X, Peters NR, Chisholm JD, Zhang G, Gilbert EJ, Wang PG, Van Vranken DL, Thorson JS, ChemBioChem 2006, 7, 795–804; [DOI] [PubMed] [Google Scholar]; h) Salas AP, Zhu L, Sáanchez C, Braña AF, Rohr J, Méendez C, Salas JA, Mol. Microbiol 2005, 58, 17–27; [DOI] [PMC free article] [PubMed] [Google Scholar]; i) Hyun CG, Bililign T, Liao J, Thorson JS, ChemBioChem 2003, 4, 114–117; [DOI] [PubMed] [Google Scholar]; j) Sáanchez C, Butovich IA, Braña AF, Rohr J, Méendez C, Salas JA, Chem. Biol 2002, 9, 519–531. [DOI] [PubMed] [Google Scholar]
- [4].a) Calce E, Digilio G, Menchise V, Saviano M, De Luca S, Chem. Eur. J 2018, 24, 6231–6238; [DOI] [PubMed] [Google Scholar]; b) Kopycki J, Wieduwild E, Kohlschmidt J, Brandt W, Stepanova AN, Alonso JM, Pedras MS, Abel S, Grubb CD, Biochem. J 2013, 450, 37–46; [DOI] [PubMed] [Google Scholar]; c) Jahn M, Marles J, Warren R, Withers S, Angew. Chem. Int. Ed 2003, 42, 352–354; [DOI] [PubMed] [Google Scholar]; Angew. Chem 2003, 115, 366–368. [Google Scholar]
- [5].a) Chen D, Chen R, Xie K, Yue T, Zhang X, Ye F, Dai J, Org. Lett 2018, 20, 1634–1637; [DOI] [PubMed] [Google Scholar]; b) Chen D, Sun L, Chen R, Xie K, Yang L, Dai J, Chem. Eur. J 2016, 22, 5873–5877; [DOI] [PubMed] [Google Scholar]; c) Chen D, Chen R, Wang R, Li J, Xie K, Bian C, Sun L, Zhang X, Liu J, Yang L, Ye F, Yu X, Dai J, Angew. Chem. Int. Ed 2015, 54, 12678–12682; [DOI] [PubMed] [Google Scholar]; Angew. Chem 2015, 127, 12869–12873; [Google Scholar]; d) Foshag D, Campbell C, Pawelek PD, Biochim. Biophys. Acta 2014, 1844, 1619–1630; [DOI] [PubMed] [Google Scholar]; e) Gutmann A, Krump C, Bungaruang L, Nidetzky B, Chem. Commun 2014, 50, 5465–5468; [DOI] [PubMed] [Google Scholar]; f) Li L, Wang P, Tang Y, J. Antibiot 2014, 67, 65–70; [DOI] [PubMed] [Google Scholar]; g) Wang F, Zhou M, Singh S, Yennamalli RM, Bingman CA, Thorson JS, Phillips GN Jr., Proteins 2013, 81, 1277–1282; [DOI] [PMC free article] [PubMed] [Google Scholar]; h) Gutmann A, Nidetzky B, Angew. Chem. Int. Ed 2012, 51, 12879–12883; [DOI] [PubMed] [Google Scholar]; Angew. Chem 2012, 124, 13051–13056; [Google Scholar]; i) Härle J, Günther S, Lauinger B, Weber M, Kammerer B, Zechel DL, Luzhetskyy A, Bechthold A, Chem. Biol 2011, 18, 520–530; [DOI] [PubMed] [Google Scholar]; j) Mittler M, Bechthold A, Schulz GE, J. Mol. Biol 2007, 372, 67–76; [DOI] [PubMed] [Google Scholar]; k) Baig I, Kharel M, Kobylyanskyy A, Zhu L, Rebets Y, Ostash B, Luzhetskyy A, Bechthold A, Fedorenko VA, Rohr J, Angew. Chem. Int. Ed 2006, 45, 7842–7846; [DOI] [PMC free article] [PubMed] [Google Scholar]; Angew. Chem 2006, 118, 8006–8010; [Google Scholar]; l) Liu T, Kharel MK, Fischer C, McCormick A, Rohr J, ChemBioChem 2006, 7, 1070–1077; [DOI] [PMC free article] [PubMed] [Google Scholar]; m) Fischbach MA, Lin H, Liu DR, Walsh CT, Proc. Natl. Acad. Sci. USA 2005, 102, 571–576; [DOI] [PMC free article] [PubMed] [Google Scholar]; n) Bililign T, Hyun CG, Williams JS, Czisny AM, Thorson JS, Chem. Biol 2004, 11, 959–969. [DOI] [PubMed] [Google Scholar]
- [6].a) Kim TS, Le TT, Nguyen HT, Cho KW, Sohng JK, Enzyme Microb. Technol 2018, 118, 76–82; [DOI] [PubMed] [Google Scholar]; b) Chu LL, Pandey RP, Lim HN, Jung JH, Thuan NH, Kim TS, Sohng JK, J. Biol. Eng 2017, 11, 15; [DOI] [PMC free article] [PubMed] [Google Scholar]; c) Pandey RP, Parajuli P, Gurung RB, Sohng JK, Enzyme Microb. Technol 2016, 91, 26–33; [DOI] [PubMed] [Google Scholar]; d) Pandey RP, Parajuli P, Shin JY, Lee J, Lee S, Hong YS, Park Y, Kim JS, Sohng JK, Appl. Environ. Microbiol 2014, 80, 7235–7243; [DOI] [PMC free article] [PubMed] [Google Scholar]; e) Pandey RP, Li TF, Yamaguchi T, Park YI, Kim JS, Sohng JK, Appl. Environ. Microbiol 2013, 79, 3516–3521; [DOI] [PMC free article] [PubMed] [Google Scholar]; f) Gurung RB, Kim EH, Oh TJ, Sohng JK, Mol. Cells 2013, 36, 355–361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [7].a) Hughes RR, Shaaban KA, Zhang J, Cao H, Phillips GN, Thorson JS, ChemBioChem 2017, 18, 363–336; [DOI] [PMC free article] [PubMed] [Google Scholar]; b) Zhang J, Hughes R, Saunders M, Elshahawi S, Ponomareva L, Zhang Y, Winchester S, Scott S, Sunkara M, Morris AJ, Prendergast M, Thorson JS, J. Nat. Prod 2017, 80, 12–18; [DOI] [PMC free article] [PubMed] [Google Scholar]; c) Chen Z, Zhang J, Singh S, Pelter-Pain P, Thorson JS, Hinds B, ACS Nano 2014, 8, 8104–8112; [DOI] [PMC free article] [PubMed] [Google Scholar]; d) Zhang J, Singh S, Hughes RR, Zhou M, Sunkara M, Morris AJ, Thorson JS, ChemBioChem 2014, 15, 647–652; [DOI] [PMC free article] [PubMed] [Google Scholar]; e) Gantt RW, Peltier-Pain P, Singh S, Zhou M, Thorson JS, Proc. Natl. Acad. Sci. USA 2013, 110, 7648–7653; [DOI] [PMC free article] [PubMed] [Google Scholar]; f) Gantt RW, Peltier-Pain P, Cournoyer WJ, Thorson JS, Nat. Chem. Biol 2011, 7, 685–691; [DOI] [PMC free article] [PubMed] [Google Scholar]; g) WIlliams GJ, Yang J, Zhang C, Thorson JS, ACS Chem. Biol 2011, 6, 95–100; [DOI] [PMC free article] [PubMed] [Google Scholar]; h) Zhou M, Thorson JS, Org. Lett 2011, 13, 2786–2788; [DOI] [PMC free article] [PubMed] [Google Scholar]; i) Gantt RW, Goff RD, Williams GJ, Thorson JS, Angew. Chem. Int. Ed 2008, 47, 8889–8892; [DOI] [PMC free article] [PubMed] [Google Scholar]; Angew. Chem 2008, 120, 9021–9024; [Google Scholar]; j) WIlliams GJ, Goff R, Thorson JS, Chem. Biol 2008, 15, 393–401; [DOI] [PMC free article] [PubMed] [Google Scholar]; k) Williams GJ, Thorson JS, Nat. Protoc 2008, 3, 357–362; [DOI] [PubMed] [Google Scholar]; l) Zhang C, Bitto E, Goff RD, Singh S, Bingman CA, Griffith BR, Albermann C, Phillips GN Jr., Thorson JS, Chem. Biol 2008, 15, 842–853; [DOI] [PMC free article] [PubMed] [Google Scholar]; m) Zhang C, Moretti R, Jiang J, Thorson JS, ChemBioChem 2008, 9, 2506–2514; [DOI] [PMC free article] [PubMed] [Google Scholar]; n) Williams GJ, Zhang C, Thorson JS, Nat. Chem. Biol 2007, 3, 657–662; [DOI] [PubMed] [Google Scholar]; o) Zhang C, Fu Q, Albermann C, Li L, Thorson JS, ChemBioChem 2007, 8, 385–390; [DOI] [PubMed] [Google Scholar]; p) Zhang C, Albermann C, Fu X, Thorson JS, J. Am. Chem. Soc 2006, 128, 16420–16421; [DOI] [PubMed] [Google Scholar]; q) Zhang C, Griffith BR, Fu Q, Albermann C, Fu X, Lee IK, Li L, Thorson JS, Science 2006, 313, 1291–1294; [DOI] [PubMed] [Google Scholar]; r) Minami A, Uchida R, Eguchi T, Kakinuma K, J. Am. Chem. Soc 2005, 127, 6148–6149; [DOI] [PubMed] [Google Scholar]; s) Quirós LM, Aguirrezabalaga I, Olano C, Méendez C, Salas JA, Mol. Microbiol 1998, 28, 1177–1185; [DOI] [PubMed] [Google Scholar]; t) Hernández C, Olano C, Méendez C, Salas JA, Gene 1993, 134, 139–140. [DOI] [PubMed] [Google Scholar]
- [8].a) Yoshida M, Iwamoto Y, Uozumi T, Beppu T, Agric. Biol. Chem 1985, 49, 563–565; [Google Scholar]; b) Tsuji N, Kobayashi M, J. Antibiot 1978, 31, 939–944. [DOI] [PubMed] [Google Scholar]
- [9].a) Zhou M, Hamza A, Zhan C, Thorson JS, J. Nat. Prod 2013, 76, 279–286; [DOI] [PMC free article] [PubMed] [Google Scholar]; b) Zhou M, Hou Y, Hamza A, Pain C, Zhan C, Bugni T, Thorson JS, Org. Lett 2012, 14, 5424–5427; [DOI] [PMC free article] [PubMed] [Google Scholar]; c) Svehlikova V, Bennett R, Mellon F, Needs P, Piacente S, Kroon P, Bao Y, Phytochemistry 2004, 65, 2323–2332. [DOI] [PubMed] [Google Scholar]
- [10].a) Wen L, Zhao Y, Jiang Y, Yu L, Zeng X, Yang J, Tian M, Liu H, Yang B, Free Radical Biol. Med 2017, 110, 92–101; [DOI] [PubMed] [Google Scholar]; b) Sasaki N, Nishizaki Y, Yamada E, Tatsuzawa F, Nakatsuka T, Takahashi H, Nishihara M, FEBS Lett 2015, 589, 182–187; [DOI] [PubMed] [Google Scholar]; c) Shaaban K, Ahmed KT, Leggas M, Rohr J, J. Nat. Prod 2012, 75, 1383–1392; [DOI] [PMC free article] [PubMed] [Google Scholar]; d) Lin C, Kuo S, Chung M, Ko F, Teng C, J. Nat. Prod 1997, 60, 851–853. [DOI] [PubMed] [Google Scholar]
- [11].a) Xie K, Chen R, Chen D, Li J, Wang R, Yang L, Dai J, Adv. Synth. Catal 2017, 359, 603–608; [Google Scholar]; b) Shaaban KA, Elshahawi SI, Wang X, Horn J, Kharel M, Leggas M, Thorson JS, J. Nat. Prod 2015, 78, 1723–1729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [12].Goff RD, Thorson JS, MedChemComm 2014, 5, 1036–1047; and references therein. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [13].a) Singh G, Verma AK, Kumar V, 3Biotech 2016, 6, 3; [DOI] [PMC free article] [PubMed] [Google Scholar]; b) Hancock S, Vaughan M, Withers S, Curr. Opin. Chem. Biol 2006, 10, 509–519; [DOI] [PubMed] [Google Scholar]; c) Berrin J, Czjzek M, Kroon P, McLauchlan W, Puigserver A, Williamson G, Juge N, Biochem. J 2003, 373, 41–48; [DOI] [PMC free article] [PubMed] [Google Scholar]; d) Davies G, Henrissat B, Structure 1995, 3, 853–859; [DOI] [PubMed] [Google Scholar]; e) Daniels L, Coyle P, Chiao Y, Glew R, Labow R, J. Biol. Chem 1981, 256, 13004–13013. [PubMed] [Google Scholar]
- [14].a) Porter N, Christianson D, Curr. Opin. Struct. Biol 2019, 59, 9–18; [DOI] [PMC free article] [PubMed] [Google Scholar]; b) Li Y, Wang F, Chen X, Wang J, Zhao Y, Li Y, He B, Curr. Top. Med. Chem 2019, 19, 223–241; [DOI] [PubMed] [Google Scholar]; c) Sagwan R, Rajan R, Mandal PK, Eur. J. Med. Chem 2018, 158, 620–706; [DOI] [PubMed] [Google Scholar]; d) Porter N, Christianson D, ACS Chem. Biol 2017, 12, 2281–2286; [DOI] [PMC free article] [PubMed] [Google Scholar]; e) Bieliauskas A, Weerasinghe S, Negmeldin A, Pflum MK, Arch. Pharm 2016, 349, 373–382; [DOI] [PMC free article] [PubMed] [Google Scholar]; f) Shen S, Kozikowski A, ChemMedChem 2016, 11, 15–21; [DOI] [PMC free article] [PubMed] [Google Scholar]; g) Wu R, Lu Z, Cao Z, Zhang Y, J. Am. Chem. Soc 2011, 133, 6110–6113; [DOI] [PMC free article] [PubMed] [Google Scholar]; h) Bieliauskas AV, Pflum MK, Chem. Soc. Rev 2008, 37, 1402–1413; [DOI] [PMC free article] [PubMed] [Google Scholar]; i) Xu W, Parmigiani R, Marks P, Oncogene 2007, 26, 5541–5552; [DOI] [PubMed] [Google Scholar]; j) Vanhaecke T, Papeleu P, Elaut G, Rogiers V, Curr. Med. Chem 2004, 11, 1629–1643. [DOI] [PubMed] [Google Scholar]
- [15].a) Kohge T, Gohda E, Okamura T, Yamamoto I, Biochem. Pharmacol 1998, 56, 1359–1364; [DOI] [PubMed] [Google Scholar]; b) Yoshida M, Nomura S, Beppu T, Cancer Res 1987, 47, 3688–3691. [PubMed] [Google Scholar]
- [16].a) Zhang Y, Ye Q, Ponomareva LV, Cao Y, Liu Y, Cui Z, Van Lanen SG, Voss SR, She Q-B, Thorson JS, Chem. Sci 2019, 10, 7641–7648; [DOI] [PMC free article] [PubMed] [Google Scholar]; b) Wang X, Abbas M, Zhang Y, Elshahawi SI, Ponomareva LV, Cui Z, Van Lanen SG, Sajid I, Voss S…R, Shaaban KA, Thorson JS, J. Nat. Prod 2019, 82, 1686–1693; [DOI] [PMC free article] [PubMed] [Google Scholar]; c) Voss SR, Ponomareva LV, Dwaraka VB, Pardue KE, Baddar NWAH, Rodgers AK, Woodcock MR, Qiu Q, Crowner A, Blichmann D, Khatri S, Thorson JS, Sci. Rep 2019, 9, 6751; [DOI] [PMC free article] [PubMed] [Google Scholar]; d) Wang X, Zhang Y, Ponomareva LV, Qiu Q, Woodcock R, Elshahawi SI, Chen X, Zhou Z, Hatcher BE, Hower JC, Zhan CG, Parkin S, Kharel MK, Voss SR, Shaaban KA, Thorson JS, Angew. Chem. Int. Ed 2017, 56, 2994–2998; [DOI] [PMC free article] [PubMed] [Google Scholar]; Angew. Chem 2017, 129, 3040–3044; [Google Scholar]; e) Ponomareva LV, Athippozhy A, Thorson JS, Voss SR, Comp. Biochem. Physiol. Part C 2015, 178, 128–135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [17].a) Rais R, Vavar J, Tichy T, Dash R, Gadiano A, Tenora L, Monincova L, Barinka C, Alt J, Zimmermann S, Slusher C, Wu Y, Wozniak K, Majer P, Tsukamoto T, Slusher B, J. Med. Chem 2017, 60, 7799–7809; [DOI] [PMC free article] [PubMed] [Google Scholar]; b) McClure J, Zhang C, Inks E, Peterson Y, Li J, Chou CJ, J. Med. Chem 2016, 59, 9942–9959; [DOI] [PMC free article] [PubMed] [Google Scholar]; c) Moradi SV, Hussein WM, Varamine P, Simerska P, Toth I, Chem. Sci 2016, 7, 2492–2500; [DOI] [PMC free article] [PubMed] [Google Scholar]; d) Jones EM, Polt R, Front. Chem 2015, 3, 40; [DOI] [PMC free article] [PubMed] [Google Scholar]; e) Calvaresi EC, Hergenrother PJ, Chem. Sci 2013, 4, 2319–2333; [DOI] [PMC free article] [PubMed] [Google Scholar]; f) Thomas M, Rivault F, Tranoy-Opalinski I, Roche J, Gesson JP, Papot S, Bioorg. Med. Chem. Lett 2007, 17, 983–986. [DOI] [PubMed] [Google Scholar]
- [18].a) Hermant P, Bosc D, Piveteau C, Gealageas R, Lam B, Ronco C, Roigant M, Tolojanahary H, Jean L, Renard P, Lemdani M, Bourotte M, Herledan A, Bedard C, Biela A, Leroux F, Deprez B, Deprez-Poulain R, J. Med. Chem 2017, 60, 9067–9089; [DOI] [PubMed] [Google Scholar]; b) Wang L, Ramirez J, Yeo W, Chan M, Thuya W, Lau J, Wan S, Wong A, Zee Y, Lim R, Lee S, Ho P, Lee H, Chan A, Ansher S, Ratain M, Goh B, PLoS One 2013, 8, e54522; [DOI] [PMC free article] [PubMed] [Google Scholar]; c) Kang SP, Ramirez J, House L, Zhang W, Mirkov S, Liu W, Haverfield E, Ratain MJ, Pharmacogenet. Genomics 2010, 20, 638–641; [DOI] [PMC free article] [PubMed] [Google Scholar]; d) Balliet RM, Chen G, Gallagher CJ, Dellinger RW, Sun D, Lazarus P, Cancer Res 2009, 69, 2912–2918; [DOI] [PMC free article] [PubMed] [Google Scholar]; e) Mulder GJ, Meerman JH, Environ. Health Perspect 1983, 49, 27–32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [19].a) Hull E, Montgomery M, Leyva K, Biomed. Res. Int 2016, 8797206; [DOI] [PMC free article] [PubMed] [Google Scholar]; b) Falkenberg K, Johnstone R, Nat. Rev. Drug Discovery 2014, 13, 673–691; [DOI] [PubMed] [Google Scholar]; c) Sweet M, Shakespear M, Kamal N, Fairlie D, Immunol. Cell Biol 2012, 90, 14–22; [DOI] [PubMed] [Google Scholar]; d) Shakespear M, Halili M, Irvine K, Fairlie D, Sweet M, Trends Immunol 2011, 32, 335–343. [DOI] [PubMed] [Google Scholar]
- [20].a) Bensaid D, Blondy T, Deshayes S, Dehame V, Bertrand P, Gregoire M, Errami M, Blanguart C, Clin. Epigenet 2018, 79; [DOI] [PMC free article] [PubMed] [Google Scholar]; b) Conte M, De Palma R, Altucci L, Int. J. Biochem. Cell Biol 2018, 98, 65–74; [DOI] [PubMed] [Google Scholar]; c) Booth L, Roberts JL, Poklepovic A, Kirkwood J, Dent P, Oncotarget 2017, 8, 83155–83170; [DOI] [PMC free article] [PubMed] [Google Scholar]; d) McCaw T, Randall T, Forero A, Buchsbaum D, Immunotherapy 2017, 9, 1359–1372; [DOI] [PMC free article] [PubMed] [Google Scholar]; e) Kroesen M, Gielen P, Brok IC, Armandari I, Hoogerbrugge PM, Adema GJ, Oncotarget 2014, 5, 6558–6572; [DOI] [PMC free article] [PubMed] [Google Scholar]; f) Shen L, Pili R, Oncoimmunol 2012, 1, 948–950. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [21].a) Ceccacci E, Minucci S, Br. J. Cancer 2016, 114, 605–611; [DOI] [PMC free article] [PubMed] [Google Scholar]; b) Ravasio R, Ceccacci E, Minucci S, Curr. Opin. Genet. Dev 2016, 36, 92–99; [DOI] [PubMed] [Google Scholar]; c) Gryder B, Sodji Q, Oyelere A, Future Med. Chem 2012, 4, 505–524; [DOI] [PMC free article] [PubMed] [Google Scholar]; d) Khan O, La Thangue N, Immunol. Cell Biol 2012, 90, 85–94; [DOI] [PubMed] [Google Scholar]; e) Wagner J, Hackanson B, Lubbert M, Jung M, Clin. Epigenet 2010, 1, 117–136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [22].Archin N, Kirchherr J, Sung J, Clutton G, Sholtis K, Xu Y, Allard B, Stuelke E, Kashuba A, Kuruc J, Eron J, Gay C, Goonetilleke N, Margolis D, J. Clin. Invest 2017, 127, 3126–3135. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.