

Abstract
Innate, non-specific, resistance mechanisms are important barriers to pathogens, particularly delaying virus multiplication at the onset of infections. These innate defense mechanisms include a series of mechanical barriers, pre-existing inhibitory molecules, and cellular responses with antimicrobial activity. The antiviral activities of these innate inhibitors reside in a variety of partly characterized substances. This review presents the innate antiviral inhibitors in cell cultures, urine, serum, the gastrointestinal tract, the nervous system, tissues of crustaceans, and saliva. Medical adaptation of the innate antiviral defense mechanisms may be useful for prevention and treatment of viral infections.
Keywords: Antiviral defenses, Innate antivirals
1. Introduction
Innate, non-specific, resistance mechanisms are important barriers to pathogens, particularly in delaying virus multiplication at the onset of infections and prior to development of the specific immune response (Dianzani and Baron, 1996). The reactive, non-specific defenses (e.g. interferon, cytokines, and activated cells) are not included because they are not innate since they are produced in response to infection (Baron, 1963). The innate defenses include a series of mechanical barriers, pre-existing molecules, and cellular responses with antimicrobial activity.
Prior to the recognition of non-specific defenses, scientists believed that viral infections occurred in nonimmune hosts. They believed that the body's specific immune responses (antibody and immune cells) were sufficient to overcome the invading viruses starting at about day 5 (Baron, 1963). However, recognition that earlier host defenses existed was hinted at by a key finding — that the specific immune responses began several days after virus multiplication was brought under control (Baron, 1963) (Fig. 1 ). These early defenses were determined to be non-specific defenses — both innate and reactive.
Fig. 1.
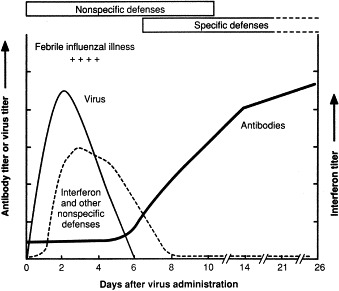
Production of virus, interferon, and antibody during experimental infection of humans with influenza wild-type virus. Non-specific defenses include anatomic barriers, inhibitors, phagocytosis, fever, inflammation and IFN. Specific defenses include antibody and cell-mediated immunity. Data from a study by B. Murphy et al., National Institutes of Health (pers. commun.).
Subsequent studies confirmed that most viral infections are initially limited by the early non-specific defenses, which restrict the initial virus multiplication to manageable levels and initiate recovery. Recovery is then completed by a combination of these early non-specific defenses and the subsequent virus-specific immune defenses. The early non-specific defenses enable the host to cope with the peak accumulation of viruses that, if presented as the initial infecting dose, could be lethal. Although non-specific and immune defenses operate together to control viral infections, this review considers mainly innate, non-specific, inhibitory substances.
The role of the innate inhibitors as host defenses was suggested by their presence in vivo, their significant antiviral activity as titered in tissue culture, and by the direct correlation between the degree of virulence of some viruses and their degree of resistance to certain inhibitors. Such correlations have been made in studies of the serum and mucus inhibitors of influenza viruses during experimental infections (Kriznova and Rathova, 1969). In addition, the presence of these inhibitors in vivo may explain the relatively high dose of virus required to initiate infection in vivo, compared with the low dose needed in cell cultures.
Normal body fluids and tissues have been reported to possess inhibitory activities against discrete viruses, individual virus groups, or a broad spectrum of viruses (Karzon, 1956, Kriznova and Rathova, 1969, Kitamura et al., 1973, Shortridge and Ho, 1974, Welsh et al., 1976, Thiry et al., 1978, Suribaldi et al., 1979, Gerna et al., 1980, Singh and Baron, 2000). Since the antiviral activities against single or small groups of viruses have been poorly characterized, there has not been enough information to fully understand their role as host defense mechanisms. More recently, molecules possessing broad antiviral activity in the body have been better characterized and also may be considered as potentially effective defenses against viral infections. Below we will focus this review mainly on the properties of these broadly antiviral innate substances using the background literature of the more narrowly acting inhibitors.
Throughout this review, we refer to antiviral titer or activity as units/ml (U/ml). One unit/ml of antiviral activity was defined as the reciprocal of the highest dilution of the inhibitor preparation showing 50% reduction of plaques compared to medium control. In this kind of assay, 64 U of antiviral activity means that a sample, after 64-fold dilution, can still reduce 30–40 plaque forming units of viruses by 50% in comparison with the medium control.
Cytotoxicity was excluded as a mediator of virus inhibition in our laboratories by control experiments that measured cell killing by Trypan Blue in the presence of the inhibitors and also experiments that measured cell proliferation in the presence of the inhibitors.
2. Cell cultures: inhibitor with broad antiviral activity (CVI)
A broadly active inhibitor of viruses (vaccinia, polio 1, and vesicular stomatitis) was found in the culture fluid from several, but not all, types of normal human and mouse cells in culture (Baron and McKerlie, 1981). Virus plaque-inhibiting activity appeared in culture fluids within a few hours after incubation of cultures with fresh medium. Peak inhibitory activity occurred within 24 h. Blockade of cellular ribonucleic acid or protein synthesis decreased the appearance of the inhibitor. Inhibition of virus required the simultaneous presence of inhibitor, virus, and cells, suggesting the reversible nature of the inhibition (Hughes et al., 1981). The degree of inhibitory activity depended on the animal species of origin of the inhibitor, the cell type used for the assay, and the virus type used for challenge. No cell species barrier against inhibitory action was found. Strong inhibition of multicycle yields of vesicular stomatitis virus and Sindbis virus was caused even by low doses of the inhibitor. Characterization of CVI with respect to molecular size and stability to heat and a number of chemical reagents and enzymes indicate that the antiviral activity of CVI is (a) associated with a large molecule (approximately 4 million Da), (b) stable at 100°C, and (c) resistant to the action of RNase, DNase, sulfhydral reagents, protein denaturants, and extraction by organic solvents (Sullivan et al., 1987). These characteristics are specific for CVI (Singh and Baron, 2000). Similar inhibitors occur in certain tissue extracts and body fluids (described later).
3. Urine: broad antiviral activity
Broad antiviral activity was detected in urine while screening physiologic body fluids for the presence of viral inhibitors (Coppenhaver et al., 1984, Kumar et al., 1984). Moderate levels (2–48 U/ml) of anti-vaccinia activity were consistently found in sterile filtered urine from normal donors (Baron et al., 1989a, Baron et al., 1989b). Similar amounts of antiviral activity were seen against vesicular stomatitis and herpes simplex 1 viruses. The active moiety exhibited marked pH and heat stability. Further studies, presented below, indicated that this activity is attributable to the ammonium content of urine (Baron et al., 1989a, Baron et al., 1989b).
Ammonium salts are known to exert fairly broad antiviral activity, partly by accumulating in the lysosomes and reducing the acidity of the local environment, thereby inhibiting the activity of lysosomal enzymes and impairing the uncoating process during virus infection (Marsh and Helenius, 1980). Antiviral activity in urine can be accounted for by the ammonium ion concentration. The basis for this conclusion is twofold. First, the physical characteristics and biological mechanisms of the urine inhibitor are similar to those of authentic NH4 +. Second, the quantity of ammonium salts normally present in urine is sufficient to explain the level of antiviral activity in this physiologic fluid. The ammonium ion concentration in other body fluids is much lower than that in urine, and their content of NH4 + does not generally inhibit viruses (Tietz, 1983). In addition, ammonium salts are distinct from the previously reported antiviral mucoprotein in urine (Tamm and Horsfall, 1952).
It was also determined that other simple inorganic amines can suppress plaque formation of herpes simplex type 1 virus, Semliki Forest virus, respiratory syncitial virus, rhinovirus 13, and influenza A virus. Similar results have been reported for ammonium chloride and other lysomotropic amines by other investigators (Jensen et al., 1961, Banfield and Kisch, 1973, Norkin and Einck, 1978, Helenius et al., 1980, Coombs et al., 1981). Although ammonium ions significantly inhibit several types of viruses, their breadth of antiviral activity is somewhat less than the inhibitors found in the plasma, gastrointestinal tract, and nervous system, as described later.
During natural viral infections, virus shedding in the urinary tract is known to be less than in the respiratory or gastrointestinal tracts (Utz, 1964). One of the factors that could explain this decreased virus infection of the urinary tract may be the content of ammonium ions in urine.
4. Serum: innate inhibitors
Human blood plasma has been reported early on to possess non-specific antiviral activity. This activity is reported to be due to several pre-existing naturally occurring molecules that are either active against individual or a family of viruses (Singh and Baron, 2000). These molecules, however, have not been adequately studied to reveal their molecular structures and mechanisms of action, presumably because of their low titer. Therefore, their possible role against viremia is only partly understood. Recently, two naturally occurring non-specific broad-spectrum antiviral agents, UTIβ and high-density lipoprotein (HDL), have been described (Singh et al., 1992, Singh et al., 1993). They are active against DNA and enveloped and nonenveloped RNA viruses. One of them, UTIβ, possesses substantial antiviral activity of 40 U/ml of serum. In comparison, HDL activity is 4 U/ml. Since the pre-existing antiviral molecules in serum appear to be the only circulating defense mechanisms available at the onset of viral infection, they may be of significance in restricting viremia.
As background, the narrowly acting antiviral activities are reported to reside in a variety of uncharacterized or partly characterized substances. Most of these inhibitors have been distinguished from antibody, interferon and complement. Many of these innate antiviral substances have generally been reported to act only against a single virus, such as coronavirus, Newcastle disease virus, variola virus, Sendai virus and vesicular stomatitis virus (Karzon, 1956, Kitamura et al., 1973, Thiry et al., 1978, Suribaldi et al., 1979, Gerna et al., 1980). Some are active against individual virus families, including myxoviruses, togaviruses, and retroviruses (Kriznova and Rathova, 1969, Shortridge and Ho, 1974, Welsh et al., 1976). The mechanisms of action of these viral inhibitors, when known, vary. Some of these act by irreversible neutralization of infectivity, as in the case with coronavirus and Sendai virus inhibitors (Karzon, 1956, Kitamura et al., 1973, Suribaldi et al., 1979, Gerna et al., 1980). The myxo- and togavirus inhibitors appear to prevent hemaglutination but not infectivity (Kriznova and Rathova, 1969, Shortridge and Ho, 1974). The inhibitor of vesicular stomatitis virus acts by penetration of the viral envelope and inactivation of viral RNA (Thiry et al., 1978). The retrovirus inhibitor works via complement mediated lysis of the virally infected cells (Welsh et al., 1976).
Only four broad-spectrum viral inhibitors have been reported in human serum. Interferon and TNF occur in response to infections, inflammation, and cancer (Baron et al., 1987, Sambhi et al., 1991, Czarniecki, 1993, Wallace et al., 1994). The other two are constitutive. They are the above-mentioned UTIβ and high-density lipoprotein (HDL) (Baron et al., 1989a, Baron et al., 1989b, Singh et al., 1992, Singh et al., 1999). The broad antiviral activity of UTIβ is shown in Table 1 . Heparin, which may be used medically to prevent clotting, has narrow antiviral activity but deserves to be mentioned because of its potent activity against HIV-1 (Nahmias and Kibrick, 1964, Baba et al., 1988, Rider, 1997). Below we will consider the properties of the constitutive broadly active inhibitors, i.e. UTIβ and HDL.
Table 1.
Broad antiviral activity of UTIβ
| Virus | Antiviral titer (U/ml)a |
|---|---|
| DNA viruses | |
| Herpesviruses | |
| Herpes simplex (HSV-1) | 36 |
| Poxviruses | |
| Vaccina | 24 |
| RNA viruses | |
| Picornaviruses | |
| Polio | 132 |
| Mengo | 36 |
| Paramyxoviruses | |
| Newcastle disease (NDV) | 144 |
| Alphaviruses | |
| Semliki Forest | 48 |
| Sindbis | 96 |
| Flaviviruses | |
| Banzi | 64 |
| Bunyaviruses | |
| Bunyamwera | 96 |
| Rhabdoviruses | |
| Vesicular stomatitis (VSV) | 96 |
Differences ≥3-fold are significant by Student's t-test.
4.1. Molecular properties of UTIβ and HDL
4.1.1. UTIβ
As determined by enzymatic inactivation, UTIβ is a glycoprotein. It has a molecular weight of approximately 60 000±10 000 Da based on HPLC size exclusion chromatography (Fig. 2 ). Its antiviral activity of 40 plaque inhibitory units/ml, is stable at pH 2 through 10 and at 80°C for up to 10 min. Mild oxidation by sodium periodate (NaIO4) and glycolysis by a mixture of glycosidases destroys its antiviral activity. Proteinase digestion degrades the inhibitor into small products of <1000 Da, which retain broad antiviral activity. This activity of the small components has increased heat stability (120°C for 15 min) and is still inactivated by glycosidases. Thus the antiviral activity appears to reside mainly in a separable oligosaccharide moiety of the glycoprotein, UTIβ (Singh et al., 1992).
Fig. 2.
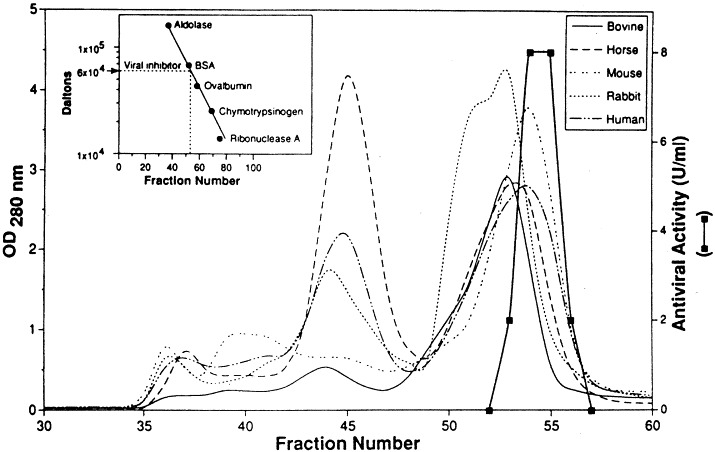
Estimation of the molecular size of the UTIβ by size exclusion chromatography. Sera from different mammalian species were individually loaded onto a column (2.5×90 cm) of Sephacryl S-200. The column was equilibrated with phosphate buffered saline. Four-milliliter fractions were collected: the antiviral activity (depicted by the heavy line) in individual fractions was examined by the standard plaque reduction assay. The inset shows the calibration of the column. Reprinted with permission from the J. Biol. Regul. Homeost. Agents, 1993, 7: 9.
4.1.2. HDL
Human serum HDL is a lipoprotein of 300 kDa and occurs in the serum in the range of 30–80 mg/dl. Its antiviral activity in serum is about four virus plaque-inhibitory units per ml. The antiviral activity seems to reside in its protein component, apolipoprotein A-1. Apolipoprotein A-1 has been reported to inhibit HIV (Owens et al., 1990) and HSV1 (Srinivas et al., 1990). HDL is also broadly active against DNA, and enveloped and non-enveloped viruses. Thus, the main broad-spectrum viral inhibitors in serum are UTIβ and, to a lesser degree, HDL (Singh et al., 1999).
4.2. Mechanisms of antiviral action of UTIβ and HDL
Preincubation of virus with UTIβ or HDL did not result in reduction of infectivity, indicating that they do not irreversibly neutralize viral infectivity. Similarly, preincubation of cells with HDL and UTIβ, unlike interferon, does not induce an antiviral state in the cells. Inhibition experiments at 4 and 37°C revealed that UTIβ inhibits virus by preventing virus attachment. At 4°C, the cell membrane is physiologically inert, and virus replication at 4°C does not proceed beyond initial attachment to the target cell. Comparable titers at 4 and 37°C, therefore, imply that an antiviral substance inhibits the attachment of virus to target cells. Significantly higher titers at 37 than at 4°C imply that the inhibitor acts at a post-attachment stage.
This type of experiment showed that HDL, unlike UTIβ, acts against most viruses at a post-attachment stage. Further investigations of the mechanism of viral inhibition by HDL indicated that HDL inhibits at an early stage of the multiplication cycle, between 0 and 1 h of initiation of infection, most likely by preventing penetration of cell surface by virus (Singh et al., 1999).
4.3. Natural defensive role of narrowly acting inhibitors in serum
A possible defensive role of the narrowly active serum inhibitors has been reviewed by Kriznova and Rathova (1969). The mechanisms of action of these inhibitors are competitive inhibition of virus attachment to cells, and neutralization of viral infectivity by a mechanism unlike classical antibody. The various approaches undertaken to elucidate the possible role of these serum inhibitors in natural resistance were grouped into three categories. In the first category an effort was made to find a correlation between the susceptibility of experimental animals to a particular virus, and the natural inhibitory activity of their sera (Bang et al., 1950, Smorodintsev, 1957, Smorodintsev, 1960). The correlations showed that resistance of the experimental animals to several strains of influenza virus correlated with the presence of innate serum inhibitors against these viral strains.
In the second category, efforts were made to measure the effect of suppression or deletion of serum inhibitors on the susceptibility of the experimental animals to virus infection. In several experiments, it was shown that mice and hamsters fed ethionine (an ethyl analogue of methoinine) had appreciably reduced inhibitory activity in serum against influenza A1 and Newcastle disease viruses, compared to untreated controls. The ethionine-treated hamsters were more susceptible to influenza A1 virus as indicated by higher titers of virus in their lungs (Borecky et al., 1961, Borecky et al., 1962a, Borecky et al., 1962b). These observations point to a relationship in vivo between the level of serum inhibitors and influenza virus multiplication. Thus, they support a defensive role for the narrowly active, non-specific viral inhibitors.
The third category assessed the effect of administering inhibitors at different times during viral infection. Experimental animals were treated with an inhibitor preparation either before or immediately after virus inoculation. Using the gamma serum inhibitor, multiplication of the inhibitor-sensitive strain of influenza A2 virus in mouse lungs was inhibited. This effect was seen only when the inhibitor was administered at a time before the virus had time to penetrate susceptible cells (Davoli and Bartolomei-Corsi, 1959, Cohen, 1960, Casazza et al., 1964, Link et al., 1964, Link et al., 1965). Thus, gamma inhibitor given intranasally 4–6 h post-infection or later was not protective. However, high doses of inhibitor given i.p., even at longer intervals after infection, showed an appreciable protective effect (Casazza et al., 1964). It appears, overall, that gamma inhibitor can function as a defense mechanism under certain conditions.
5. Gastrointestinal tract viral inhibitors
Here we will consider the properties of the broadly active inhibitors associated with the murine and bovine gastrointestinal (GI) tracts (Baron et al., 1986, Singh and Baron, 2000), including size, structure, active chemical moiety and mechanisms of action. As considered elsewhere in this review, there are other broadly active viral inhibitors in several body tissues and fluids. The GI inhibitors are distinct from these other inhibitors as determined by their properties and mechanisms of antiviral action, as described below. Studies in humans have not yet been done.
The GI inhibitory preparations studied were (a) a commercial porcine mucosal extract, (b) the pharmaceutical, neuramide, and (c) mouse intestinal inhibitors eluted from the lumen or GI tissues. The porcine mucosal preparation (National Formulary XII specifications) was obtained from American Laboratories, Inc., Omaha, NE. Neuramide is a partially purified extract of gastric mucosa, prepared for treating viral infection in humans (Antonelli et al., 1986). It was obtained from DIFA-Cooper, Milan, Italy. The mouse intestinal inhibitor preparations were made from either the luminal fluids or eluates of pre-rinsed minced intestine from ICR female mice (Baron et al., 1986, Singh and Baron, 2000).
5.1. Molecular size
The intestinal samples were tested on a variety of size exclusion columns. The antiviral activity consistently emerged in the column volume for most supports, indicating that the active moiety was of very small size. On the Synchropak GPC peptide HPLC columns, on which it is possible to resolve small peptides, the neuramide, porcine mucosal, and mouse intestinal inhibitors all emerged with an apparent molecular size of <1000 Da. This is consistent with the results of dialysis experiments, where the antiviral activity was retained only by a 100 Da MWCO membrane. Based on the GPC Peptide column results, a tentative molecular weight of 600±400 Da was assigned to the antiviral moiety in these preparations.
5.2. Antiviral spectrum
The antiviral activity of all three gastrointestinal inhibitor preparations was broad in that five virus types were inhibited. The titers of the inhibitors were substantial, ranging from 24 to 64 U/ml. Broad antiviral activity was confirmed using partly purified fractions. Thus, all three gastrointestinal extracts contained significant titers of broadly antiviral substance(s).
5.3. Chemical composition required for antiviral activity
To determine the structural composition required for antiviral activity, the extracts were treated to degrade proteins, break carbohydrate linkages, oxidize carbohydrates, or extract lipids and then the residual antiviral activity was measured. Treatment of neuramide, porcine mucosal and mouse intestine preparations, with either proteinase K, carbohydrases, sodium periodate, or extraction by lipid solvents (butanol and ethyl ether), did not affect their antiviral activity. Since the carbohydrases do not degrade all possible sugar linkages, the inhibitors were also treated with NaIO4 as a general oxidizing agent without inactivation of the antiviral activity. These findings with the intestinal preparations suggest that ordinary protein, carbohydrate, and lipid moieties may not be required for the antiviral activity of GI inhibitors.
5.4. Mechanism of action
To determine whether the intestinal preparations inactivated viruses directly, viruses were pre-treated with the inhibitor and then assayed for residual infectivity. Pretreatment of Sindbis virus, vesicular stomatitis virus and mengovirus with neuramide, porcine mucosal extract, and mouse intestine inhibitors did not reduce their infectivity. This indicated that the inhibitors do not bind irreversibly to virions to permanently neutralize their infectivity.
To determine whether the inhibitors acted during attachment, or later in the replication cycle, the inhibitory titers were compared at 4 and 37°C. The titers of neuramide and porcine mucosal inhibitors against three viruses at 37°C were significantly higher than those at 4°C. This indicates that neuramide and porcine mucosal inhibitors may act by blocking viral replication at a post-attachment stage. The mouse intestine inhibitor, on the other hand, is active at 4°C, and, therefore, inhibits virus replication by preventing attachment of virions to the target cells. These possible mechanisms of action must eventually be confirmed using fully purified inhibitors.
To determine if cells acquired resistance to virus infection after being pre-treated with inhibitor, cells were incubated with the inhibitor preparations (mouse intestine, neuramide and porcine mucosa) for 24 h and then washed before virus challenge. The pre-treated cells did not exhibit any resistance to subsequent infection, indicating the inhibitors do not induce a durable antiviral state in the target cells.
The resistance to degradation by the GI inhibitors could be of significance in vivo, since to remain active in the GI tract the inhibitors must resist the action of the various lipolytic, proteolytic and glycolytic activities of the digestive process. Enzyme-resistant peptides and carbohydrates have been reported previously (Witas et al., 1983, Alpers, 1986, Zijlstra et al., 1996, Day et al., 1998, Podolsky, 2000, Shen and Xu, 2000). Thus, the structure of the GI inhibitors remains undetermined.
Consistent with a possible defensive role for the GI tract inhibitors is their naturally high titer, broad antiviral activity and location in the GI tract and its lumen. To definitely assign defensive roles, future studies should show that (a) administration of these inhibitors is protective in vivo and (b) deletion of the inhibitors enhances infections.
6. Nervous system viral inhibitor (NS)
The blood–brain barrier largely excludes most of the body's host defenses, including systemic antibodies, immune cells, and cytokines, from the central nervous system (CNS) (Baron, 1963, Cathala and Baron, 1970, Griffin, 1991). Innate antiviral molecules, already within the brain, may be candidate defenses against CNS infections, and thus, nervous system tissue extracts have been studied for their ability to inhibit virus infections (Baron, 1963, Cathala and Baron, 1970, Griffin, 1991, Singh et al., 1995, Singh et al., 1999, Baron et al., 1998).
6.1. Antiviral activity
The NS inhibitor in lamb brain tissue exhibited broad antiviral activity against DNA viruses and enveloped and non-enveloped RNA viruses (Singh et al., 1995, Baron et al., 1998). Human, bovine, ovine, porcine, lapine, murine and piscine brains all possessed similar levels of antiviral activity. In humans, this NS inhibitor is present in the brain, spinal cord, white and gray matter, and sciatic nerve, but not in the cerebrospinal fluid. Furthermore, the NS inhibitor can diffuse extracellularly from prerinsed brain tissue in vitro. The antiviral activity was not due to cell toxicity as no difference in Vero and CER cell growth over 48 h was detected in the absence or presence of up to 100 U of the inhibitor. Furthermore, transferrin and low-density lipoprotein, used as non-specific protein controls, were not antiviral.
6.2. Molecular size
Using sucrose-density gradient centrifugation (Sullivan et al., 1987, Singh et al., 1995), high levels of antiviral activity were detected in the 40 and 45% sucrose bands (Fig. 3 ). These results indicated that the antiviral factor from brain extracts was at least 4000 kDa.
Fig. 3.
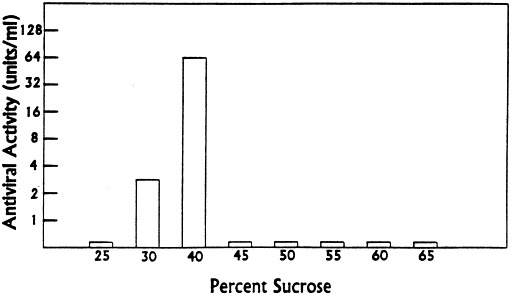
Sedimentation rate of virus inhibitor from lamb brain tissue in a discontinuous sucrose gradient. Sucrose solutions in PBS, 5 mM MgC12, were used to form step gradients. Equivalent volumes of brain inhibitor preparation (0.5 ml) and 50% sucrose were mixed, layered on the discontinuous gradient, and centrifuged at 31 000 rev./min in a SW-55 rotor for 90 min. Collected fractions were examined for antiviral activity against Sindbis virus after extensive dialysis. Reprinted with permission from Antiviral Res., 1995, 27:380.
Lipid extraction reduced the size of the antiviral components without inactivating the inhibitory activity. The antiviral activity of the lipid-extracted material from both mouse and lamb brains was reduced to a molecular weight of less than 5000 Da. Equivalent sizing results were obtained using Superdex 200 and Superdex 75 FPLC sizing columns. Also analyzed was the size of the antiviral material released by butanol–ether extraction using GPC peptide HPLC columns. Again, the antiviral activity emerged near the lower limit of the resolving power of the columns, with a calculated molecular weight of 650±300 Da. It appears that the butanol–ether extraction of the high molecular weight antiviral material from brain tissue releases a low molecular weight moiety (≤1000 Da) that possesses broad antiviral activity.
6.3. Chemical composition and thermostability
Thermostability studies showed that the native brain tissue antiviral activity was stable at 100°C for at least 12 h, but not at 120°C. The inhibitory activity was also stable following treatment with 0.01 M DTT, n-butanol and ethyl ether. Viral inhibitory activity of the crude brain extract was abolished by proteolysis, enzymatic glycolysis, or periodate oxidation. This implies that the native inhibitor molecule contains both protein and carbohydrate structure, which are necessary to maintain antiviral activity. However, the lipid-extracted small inhibitor molecule was resistant to proteolysis and glycolysis and was more heat resistant (stable at 120°C for 15 min) than the native inhibitor. Taken together, the evidence indicates that the natural inhibitor moiety in brain, is composed of lipid, protein, and carbohydrate, and disruption of its structure by proteolysis or glycolysis inactivates its inhibitory activity.
6.4. Mechanism of action
Pretreatment of viruses with inhibitor for 2 h did not affect their infectivity, indicating that the inhibitor neither bound irreversibly to virions nor did it permanently neutralize infectivity. To determine whether the inhibitor acted during attachment or later in the replication cycle, inhibitory titers at 4 and 37°C were compared. In additional experiments, virus yields were determined after timed addition of an inhibitor preparation to cell monolayers. Taken together, the results indicated inhibition at the attachment stage of the virus multiplication.
Pretreatment of cells did not induce any resistance to subsequent infection with viruses, indicating that unlike IFN, the inhibitory material does not induce a durable antiviral state in the target cells.
6.5. Effect of virus infection on inhibitor levels in vivo
The possible regulation of the NS antiviral activity levels in neural tissues in response to an ongoing infection was assessed using an encephalitic flavivirus infection of mice. Inhibitor titers were determined over 7 days of infection with the encephalitic Banzi virus (Singh et al., 1995). This timing was chosen since BZV can be detected in mouse brain from day 3 p.i. (Singh et al., 1989). No change in inhibitory titer was seen throughout this lethal infection (Fig. 4 ), indicating that the inhibitor, unlike IFN or antibody, is not specifically induced or decreased in response to viral infection.
Fig. 4.
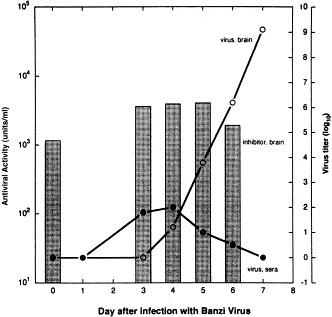
Level of virus inhibitor in mouse brain extracts is unchanged during encephalitis. Three-week-old female outbred (ICR) mice were infected intraperitoneally with 3 LD75 of BZV. Brains were harvested at days 3–6 postinfection, and homogenized. Virus inhibitory levels in the homogenates were titered against Sindbis virus, and are depicted in the bar graph (left Y axis). The rise in the virus titers in the brains and sera of infected mice (taken from Singh et al., 1989) are superimposed to illustrate the course of the BZV infections (○, virus load in brain; •, virus load per ml serum). Reprinted with permission from Antiviral Res., 1995, 27:384.
6.6. Pathogenesis studies
The possibility that this constitutive antiviral inhibitor in the nervous system could affect pathogenesis was studied by determining (Baron et al., 1998) whether the inhibitor was: (a) widely distributed in the human nervous system and released extracellularly, (b) present in effective concentrations and (c) protective in vivo (see below).
6.6.1. Distribution in the nervous system
It was found that the NS inhibitor is widely distributed in the human white matter, gray matter, spinal cord and peripheral sciatic nerve tissues, but not in cerebrospinal fluid (which excludes large molecules). These inhibitory activities from the human nervous system had the same properties as the inhibitor in other mammalian brains (Singh et al., 1995). These properties are 4000 kDa size, complex structure of lipid and essential protein and carbohydrate (presumably an aggregation into micellar structure), competitive inhibition of virus attachment, and broad antiviral activity. Thus, the NS inhibitor is located in the nervous system tissues that can be infected by viruses.
Importantly, the inhibitory activity diffused from prewashed minced brain into the extracellular fluid. This extracellular space is used by many viruses to spread. Thus the inhibitor is in the appropriate extracellular neural compartments to interdict virus attachment to cells.
6.6.2. Concentration of the NS inhibitor
The antiviral titer of the inhibitor in the nervous system ranged from 48 to 188 U/g of the tissue. By comparison, the titers of other host defenses such as antibody and interferon, as measured by the same type of assay, are in the same range (Baron, 1996). Thus the concentration of the NS inhibitor appears to be sufficient to play a defensive role.
6.6.3. Protective action in vivo
A putative host defense should protect in vivo as well as in vitro. In vivo protection studies demonstrated that subcutaneous administration of the NS inhibitor with an alphavirus or a picornavirus resulted in both prolongation of survival and in protection against death. These findings show that the NS inhibitor can be protective in vivo against viruses which can infect the nervous system.
Overall, a natural defensive role for the broadly antiviral NS inhibitor is suggested by its constitutively high concentration, wide distribution in tissues of the nervous system, presence in extracellular fluid and its protection of mice (Baron et al., 1998).
7. Viral inhibitors in tissues of crustaceans (Pan et al., 2000)
7.1. Overview of the crustacean inhibitors
Tissue extracts from invertebrate crustaceans, crab, shrimp and crayfish are broadly antiviral against a variety of viruses including DNA and enveloped and non-enveloped RNA viruses. Characterization of the inhibitor(s) indicated a 440-kDa molecular size for the main inhibitor. The antiviral activity of the inhibitor preparation was reduced by lipid extraction with butanol, but was unaffected by treatments with DTT, urea, proteinase K and carbohydrases. The inhibitor preparation does not directly neutralize virus infectivity. The inhibitor appears to act during the virus attachment stage. In addition, the crab inhibitor preparation also induces a durable antiviral state in Vero cells against subsequent infection of Sindbis and vaccinia viruses that was not due to interferon. The non-specific, broadly antiviral substances that are present in crustaceans may be host defense substances and may contribute to survival of crustaceans, despite continuing exposure to extremely high dose of viruses (Fuhrman, 1999).
Viruses are the most common biological agents in the sea. They number ten billion per liter and probably infect many species (Fuhrman, 1999). Viral infections are common in crustaceans. For instance, penaeid shrimp are infected by at least 20 viruses (Johnson et al., 1989, Johnson et al., 1999), while blue crabs are infected naturally or experimentally by rotavirus, enterovirus, Newcastle disease virus, and poliovirus (McCumber and Clem, 1977, Seidel et al., 1983). In comparison with vertebrates, invertebrates do not produce specific antibodies (Schapiro, 1975), and therefore may rely on innate defense for protection against viruses (Schnapp et al., 1996). Little is known about possible innate antiviral substances in invertebrates. Thus, crustaceans, such as crabs, shrimp and crayfish, are appropriate invertebrate subjects to study innate immunity against viruses (Pan et al., 2000).
7.2. Antiviral spectrum
The inhibitor extract from crabs is broadly inhibitory for the six types of DNA and RNA viruses tested. The titers of the inhibitor varied moderately with the different viruses. The inhibitory activity of tissue extracts from two other crustaceans, shrimp and crawfish, were also found to be broadly active against the same viruses with titers comparable to that of crab inhibitor preparations. No inhibitory effect of the inhibitor preparation was detected on either cell-free HIV or HIV-infected MT-2 cells.
7.3. Molecular size
The crab inhibitor preparation was fractionated by size-exclusion chromatography on a Superdex 200 26/60 FPLC column, and individual fractions were tested for the presence of antiviral activity. The antiviral peak (44 U/ml) corresponded to a molecular mass of approximately 440 kDa. In addition, a secondary peak of relatively low antiviral activity (17 U/ml) was found in fractions corresponding to a molecular size of about 15 kDa.
7.4. Possible structure as determined by enzymatic and chemical treatments
Lipid extraction with butanol significantly decreased the antiviral activity in the residual aqueous phase of the inhibitor preparation. Lipid extraction also resulted in the reduction of the size of the antiviral substance as evidenced by the filterability of the residual antiviral activity through a 10 K MWCO ultrafiltration membrane (Filtron, Northborough, MA). In contrast, neither insoluble proteinase K, nor a cocktail of glycosidases, significantly inactivated the antiviral activity of the inhibitor preparation. In addition, a sulfhydryl reagent (DTT) and a chaotropic reagent (urea) were used to treat the inhibitor preparation, and no loss of antiviral activity was found. Thus, the inhibitor is resistant to many chemical degradations.
7.5. Mode of action
The crab inhibitor preparation did not irreversibly inactivate viruses. To determine whether the inhibitor preparation acts during virus attachment or thereafter, antiviral activity at 4 and 37°C was compared, and the equal activity at 4 and 37°C indicated that one action of the crab inhibitor is to prevent virus attachment to target cells.
To determine whether the inhibitor preparation also induced durable antiviral activity in host cells, the inhibitor preparation was preincubated with Vero cells overnight and then washed before virus challenge. The inhibitor preparation-pre-treated Vero cells were resistant to subsequent infections by Sindbis and vaccinia viruses, indicating that the crab inhibitor preparation also induces a durable antiviral state in Vero cells against subsequent infection by Sindbis and vaccinia viruses. In contrast, vesicular stomatitis virus was not inhibited by the same pretreatment of cells, indicating a reversible and probably different mechanism of inhibition and the possibility of two different inhibitors.
8. Oral resistance to HIV: innate salivary defenses
8.1. Overview of the innate salivary defense against HIV
The oral cavity is unique among mucosal surfaces in having a defensive barrier to transmission of HIV and probably to the related human T-cell leukemia virus (Mohor, 1973, Phillips and Bourinbaiar, 1992, Milman and Sharma, 1994, Bomsel, 1997, Zacharopoulos and Phillips, 1997). The main defense mechanism appears to be inactivation of the transmitting leukocytes by the hypotonicity of saliva. Additional protection may be contributed by the other less active salivary inhibitors (Coppenhaver et al., 1984, Fultz, 1986, Fox et al., 1988, Fox et al., 1989, Archibald and Cole, 1990, Yeh et al., 1992, Bergey et al., 1994, McNeely et al., 1995, McNeely et al., 1997, Wahl and Orenstein, 1997, Crombie et al., 1998). This salivary defense appears to explain the rarity of casual HIV transmission orally. However, the normally effective oral barrier can be overcome by the solutes in seminal fluid, milk, and colostrums (Mayer and DeGruttola, 1987, Lifson et al., 1990, Quatro et al., 1990, Lane et al., 1991, Ryder et al., 1991, Dunn et al., 1992, Keet et al., 1992, Bertolli et al., 1996, Black, 1996, Berrey and Shea, 1997, Cao et al., 1997).
Of special importance for medical application is that the salivary defense has directed attention to the HIV-infected leukocyte as the main mucosal transmitting agent. Consequently, anticellular solutions that are more potent than saliva's hypotonicity could be applied topically at the vulnerable mucosal sites (e.g. vagina and rectum) to inactivate infected leukocytes in seminal fluid. To prevent the vaginal and rectal transmission, studies of candidate topical anticellular substances (Baron, 2000), other than the reportedly ineffective nonoxynol-9 (Mohor, 1973, Pauza et al., 1993, Pauza et al., 1994), are being done. These candidate preventives include disinfectants for blood products, bile detergents, and over-the-counter (OTC) commercial vaginal products, which are effective in vitro. Some of these OTC products are already established to be safe for use by FDA standards.
8.2. Incidence of oral transmission
When the HIV epidemic began, an initial concern was that HIV might be transmitted casually from the oral secretions of the millions of HIV carriers during kissing, dental treatment, biting, and aerosolization. Later it became clear that casual transmission is actually a rare event, even when infectious HIV is shed into the oral cavity by bleeding and exudation (Ho et al., 1985, Levy and Greenspan, 1988, Rogers et al., 1990, Gooch et al., 1993, Moore et al., 1993, Richman and Rickman, 1993, Yeung et al., 1993, Piazza et al., 1994). Saliva of infected individuals usually contains only non-infectious components of HIV, including viral DNA, indicating that the saliva may cause a breakdown of infected leukocytes. The infectivity of these salivas in vitro is only 1%, although all carriers have infectious virus in their blood (Goto et al., 1991, Barr et al., 1992, Moore et al., 1993, Coppenhaver et al., 1994, Qureshi et al., 1995, Melvin et al., 1997, Baron et al., 1999). In comparison, in seminal fluid the percentage of patients shedding HIV, mainly as infected leukocytes, is much higher — approximately 20% (Milman and Sharma, 1994, Vernazza et al., 1996, Zhu et al., 1996). Thus the rarity of casual oral transmission correlates with the low infectivity of saliva compared to the high infectivity of seminal fluid, which is associated with 27% vaginal transmission (European Study Group, 1989, Johnson et al., 1989, Lazzarin et al., 1991, European Study Group on Heterosexual Transmission of HIV, 1992, Royce et al., 1997). It is, therefore, important to determine the salivary mechanisms of protection.
8.3. Innate salivary inhibitors
There are many inhibitors of HIV in saliva (Coppenhaver et al., 1984, Fultz, 1986, Fox et al., 1988, Fox et al., 1989, Archibald and Cole, 1990, Yeh et al., 1992, Bergey et al., 1994, McNeely et al., 1995, McNeely et al., 1997, Wahl and Orenstein, 1997, Crombie et al., 1998, Shugars, 1999, Baron et al., 1999, Baron et al., 2000). An important question is ‘which of the many reported salivary inhibitors of HIV best accounts for saliva's protection against HIV-infected leukocytes?’ These salivary inhibitors include HIV-1-specific antibodies, lysozyme, peroxidases, complement, cystatins, lactoferrin, defensins, mucins, amylase, statherin, proline-rich peptides, thrombospondin-1, histatins, Rnases (Lee-Huang et al., 1999) and secretory leukocyte protease inhibitor (Coppenhaver et al., 1984, Fultz, 1986, Fox et al., 1988, Fox et al., 1989, Archibald and Cole, 1990, Sun et al., 1990, Yeh et al., 1992, Robinovitch et al., 1993, Bergey et al., 1994, McNeely et al., 1995, McNeely et al., 1997, Wahl et al., 1997, Shugars, 1999).
The existence of an undiscovered salivary inhibitor was suggested because these known salivary inhibitors did not inhibit HIV sufficiently to explain the rarity of oral transmission (Coppenhaver et al., 1984, Fultz, 1986, Fox et al., 1988, Fox et al., 1989, Archibald and Cole, 1990, Sun et al., 1990, Yeh et al., 1992, Robinovitch et al., 1993, Bergey et al., 1994, McNeely et al., 1995, McNeely et al., 1997, Wahl et al., 1997, Shugars, 1999). A new inhibitor also was suggested by the unique hypotonicity of salivas, i.e. one-seventh the tonicity of normal interstitial fluids (Ben-Aryeh et al., 1990, Edgar, 1992). This hypotonicity was shown to disrupt the crucial infected leukocytes and thereby render them incapable of supporting virus multiplication and cell-to-cell transmission of HIV (Fig. 5 ). Specifically, there was a 10 000-fold inhibition of the multiplication of HIV by the hypotonicity of saliva compared to the only 3- to 5-fold inhibition by the macromolecular inhibitors in saliva (Baron et al., 1999, Baron et al., 2000). Also, reconstitution of the isotonicity of saliva reversed its inhibition (Baron et al., 1999). Thus, hypotonic disruption appears to be a major mechanism by which saliva kills the transmitting mononuclear leukocytes and thereby prevents their attachment to mucosal epithelial cells and their production of infectious HIV.
Fig. 5.
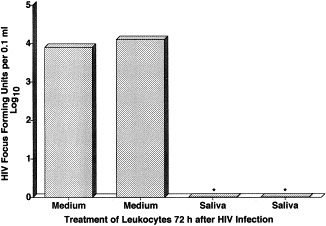
Saliva interrupts the multiplication of human immunodeficiency virus (HIV) in infected human mononuclear leukocytes. *P<0.05 by Student's t-test. Reprinted with permission from Arch. Intern. Med., 1999, 159:306.
Unexplained, however, was the paradox that oral transmission of HIV does occur epidemiologically if infected seminal fluid (Mayer and DeGruttola, 1987, Lifson et al., 1990, Quatro et al., 1990, Lane et al., 1991, Keet et al., 1992, Berrey and Shea, 1997) or milk (Ryder et al., 1991, Dunn et al., 1992, Bertolli et al., 1996, Black, 1996, Cao et al., 1997) is deposited orally, despite the presence of saliva. The mechanisms by which the carrier's seminal fluid or milk may overcome protection by the recipient's saliva was studied (Baron et al., 2000). It was hypothesized that reconstitution of salts in hypotonic saliva by orally deposited isotonic seminal fluid, milk, or colostrum would reverse saliva's ability to inactivate its virus-infected leukocytes (Girgis et al., 1980, Kavanagh, 1985, Mehl, 1986, Levy, 1988, Vonesch et al., 1992, Buranasin et al., 1993, Goldman and Goldblum, 1996). Experimentally, the volumes of seminal fluid, milk, or colostrum deposited in the mouth were found to contain sufficient solutes to overcome saliva's normal hypotonic protection against HIV-infected leukocytes and thereby appear to explain the paradoxically successful oral transmission by these fluids (Fig. 6 ). This conclusion was confirmed by the reversal of salivary protection by adding salts to saliva and by dialysis with culture medium.
Fig. 6.
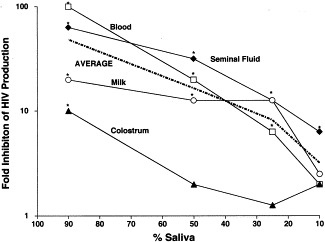
Dilution in blood, milk, colostrums, and seminal fluid prevents saliva's inhibition of HIV multiplication in human PBL. *P<0.05 Student's t-test. Reprinted with permission from J. Infect. Dis., 2000, 181:500.
9. Conclusion
Throughout the history of medicine, the most effective antimicrobial applications have come from the understanding and adaptation of natural defense mechanisms. Examples are: (1) the natural separation of the host from the microbes was adapted to sanitation and quarantine; (2) the host's immune response was adapted to vaccines and passive immunization; and (3) the microbial molecules for self-protection were adapted as antibiotics. Similar application of innate antiviral defenses, which are overviewed in Table 2 , may result in new medically useful antiviral substances and strategies.
Table 2.
Comparison of broadly active antiviral activities in cell cultures and body tissues and fluidsa
| Origin | Cell culture CVI | Serum |
Urine NH4+ | GI | NS | Crustacean | |
|---|---|---|---|---|---|---|---|
| HDL | UTIβ | ||||||
| Antiviral titer (U/ml) | 24 | 4 | 48 | 12 | 32 | 96 | 48 |
| Molecular size (kDa) | 4000 | 300 | 60 | 0.03 | 0.6 | 4000 | 440 |
| Chemical structure to sustain antiviral activity | Protein/carbohydrate | Protein | Carbohydrate | NH4+ | Undetermined | Protein/carbohydrate | Lipid |
| Mechanism of inhibition during virus multiplication cycle | Attachment | Penetration | Attachment | Uncoating | Attachment/postattachment | Attachment | Attachment/postattachment |
GI, gastrointestinal tract inhibitor; NS, nervous system inhibitor.
Acknowledgements
Studies from our laboratory have been funded, in part, by the NIH, the MacArthur Foundation, Cooper Vision Laboratories, BioFilm, Inc., and royalties through the University of Texas. The authors wish to recognize the editorial and organization contributions of Laurie Mitchell and Lyska Morrison.
References
- Alpers D.H. Uptake and fate of absorbed amino acids and peptides in the mammalian intestine. Fed. Proc. 1986;45:2261–2267. [PubMed] [Google Scholar]
- Antonelli G., Dianzani F., Coppenhaver D.H., Baron S., Calandra P., Folchitto G. An influenza virus inhibitor that acts late in the replication cycle. Antimicrob. Agents Chemother. 1986;29:49–51. doi: 10.1128/aac.29.1.49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Archibald D.W., Cole G.A. In vitro inhibition of HIV-1 infectivity by human salivas. AIDS Res. Hum. Retroviruses. 1990;6:1425–1432. doi: 10.1089/aid.1990.6.1425. [DOI] [PubMed] [Google Scholar]
- Baba M., Pauwels R., Balzarini J., Arnout J., Desmyter J., De Clercq E. Mechanism of inhibitory effect of dextran sulfate and heparin on replication of human immunodeficiency virus in vitro. Proc. Natl. Acad. Sci. USA. 1988;85:6132–6136. doi: 10.1073/pnas.85.16.6132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Banfield W.J., Kisch A.L. The effect of chloroquine on herpesvirus infection in vitro and in vivo. Proc. Soc. Exp. Biol. Med. 1973;142:1018–1022. doi: 10.3181/00379727-142-37166. [DOI] [PubMed] [Google Scholar]
- Bang F.B., Foard M., Karzon D.T. The determination and significance of substances neutralizing Newcastle disease virus in human serum. Johns Hopkins Hosp. Bull. 1950;87:130–143. [PubMed] [Google Scholar]
- Baron J.L., Li J.L., McKerlie M.L., Shbot J.M., Coppenhaver D.H. A new subtype of a natural viral inhibitor (CVI) that is stable in the gastrointestinal tract. Microb. Pathog. 1986;1:241–247. doi: 10.1016/0882-4010(86)90048-3. [DOI] [PubMed] [Google Scholar]
- Baron S. Mechanism of recovery from viral infection. Adv. Virus Res. 1963;10:39–64. doi: 10.1016/s0065-3527(08)60696-x. [DOI] [PubMed] [Google Scholar]
- Baron, S., Poast, J., Nguyen, D., Cloyd, M.W. 2000. Sexual transmission of HIV: Prevention by commercial lubricants in vitro, submitted.
- Baron S., McKerlie L. Broadly active inhibitor of viruses spontaneously produced by many cell types in culture. Infect. Immun. 1981;32:449–453. doi: 10.1128/iai.32.2.449-453.1981. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baron S., Dianzani F., Stanton G.J., Fleischmann W.R., Jr . The Interferon System: A Current Review to 1987. University of Texas Press; Austin, TX: 1987. [Google Scholar]
- Baron S., Sabados J., McKerlie M.L., Coppenhaver D.H. Antiviral activity in urine is attributable to ammonium salts. J. Biol. Regul. Homeost. Agents. 1989;3:67–70. [PubMed] [Google Scholar]
- Baron S., Niesel D., Singh I.P., McKerlie L., Poast J., Chopra A., Dianzani F., Coppenhaver D.H. Recently described innate broad spectrum inhibitors. Microb. Pathog. 1989;7:237–247. doi: 10.1016/0882-4010(89)90042-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baron S., Chopra A.K., Copenhaver D.H., Gelman B.B., Poast J., Singh I.P. A host defense role for a natural antiviral substance in the nervous system. J. Neuroimmunol. 1998;85:168–173. doi: 10.1016/s0165-5728(98)00015-0. [DOI] [PubMed] [Google Scholar]
- Baron S., Poast J., Cloyd M.W. Why is HIV rarely transmitted by oral secretions? Saliva can disrupt orally shed, infected leukocytes. Arch. Intern. Med. 1999;159:303–310. doi: 10.1001/archinte.159.3.303. [DOI] [PubMed] [Google Scholar]
- Baron S., Poast J., Richardson C.J., Nguyen D., Cloyd M. Oral transmission of human immunodeficiency virus by infected seminal fluid and milk: A novel mechanism. J. Infect. Dis. 2000;181:498–504. doi: 10.1086/315251. [DOI] [PubMed] [Google Scholar]
- Barr C.E., Miller L.K., Lopez M.R., Croxson T.S., Schwartz S.A., Denman H., Jandorek R. Recovery of infectious HIV-1 from whole saliva. J. Am. Dent. Assoc. 1992;123:36–37. doi: 10.14219/jada.archive.1992.0038. [DOI] [PubMed] [Google Scholar]
- Ben-Aryeh H., Fisher M., Szargel R., Laufer D. Composition of whole unstimulated saliva of healthy children: Changes with age. Arch. Oral Biol. 1990;35:929–931. doi: 10.1016/0003-9969(90)90075-l. [DOI] [PubMed] [Google Scholar]
- Bergey E.J., Cho M.I., Blumberg B.M., Hammarskjold M.L., Rekosh D., Epstein L.G., Levine M.J. Interaction of HIV-1 and human salivary mucins. J. Acquir. Immune Defic. Syndr. 1994;7:995–1002. [PubMed] [Google Scholar]
- Berrey M.M., Shea T. Oral sex and HIV transmission. J. Acquir. Immune Defic. Syndr. Hum. Retrovirol. 1997;14:475–477. doi: 10.1097/00042560-199704150-00012. [DOI] [PubMed] [Google Scholar]
- Bertolli J.S., Louis M.E., Simonds R.J., Nieburg P., Kamenga M., Brown C.M.T., Quinn T., Ou C.Y. Estimating the timing of mother-to-child transmission of human immunodeficiency virus in a breast-feeding population in Kinshasa, Zaire. J. Infect. Dis. 1996;174:722–726. doi: 10.1093/infdis/174.4.722. [DOI] [PubMed] [Google Scholar]
- Black R.F. Transmission of HIV-1 in the breast-feeding process. J. Am. Diet. Assoc. 1996;96:267–274. doi: 10.1016/S0002-8223(96)00079-X. [DOI] [PubMed] [Google Scholar]
- Bomsel M. Transcytosis of infectious human immunodeficiency virus across a tight human epithelial cell line barrier. Nat. Med. 1997;3:42–47. doi: 10.1038/nm0197-42. [DOI] [PubMed] [Google Scholar]
- Borecky L., Kociskova D., Hana L. An attempt to affect the level of serum inhibitors of myxoviruses in vivo. I. Effect of ethionine administration on the level of inhibitors in guinea-pig sera. Acta Virol. 1961;5:236–244. [Google Scholar]
- Borecky L., Rathova V., Kociskova D., Hana L. An attempt to affect the level of inhibitors of myxoviruses in vivo. II. Effect of ethionine administration on virus multiplication. Acta Virol. 1962;6:97–104. [Google Scholar]
- Borecky L., Rathova V., Kociskova D., Hana L. Effect of d-1 ethionine on some factors of nonspecific resistance to myxoviruses. J. Hyg. Epidemiol. 1962;6:65–70. [PubMed] [Google Scholar]
- Buranasin P., Kunakorn M., Petchclai B., Raksakait K., Wichukchiuda N., Jirapinyo M., Thongcharoen P. Detection of human immunodeficiency virus type 1 (HIV-1) proviral DNA in breast milk and colostrum of seropositive mothers. J. Med. Assoc. Thailand. 1993;76:41–45. [PubMed] [Google Scholar]
- Cao, Y., Krogstad, P., Korber, B.T., Koup, R.A., Muldoon, M., Macken, C., Song, J.-L., Jin, Z., Zhao, J.-Q., Clapp, S., Chen, I.S.Y., Ho, D.D., Ammann, A.J., and The Ariel Project Investigators, 1997. Maternal HIV-1 viral load and vertical transmission of infection: The Ariel Project for the prevention of HIV transmission from mother to infant. Nat. Med. 3, 549–552. [DOI] [PubMed]
- Casazza A.M., Di Marco A., Ghione M., Zanella A. Biological properties of horse serum inhibitor against influenza A2 viruses. Gen. Microbiol. 1964;12:1–14. [Google Scholar]
- Cathala F., Baron S. Interferon in rabbit brain, cerebrospinal fluid, and serum following administration of polyinosinic polycytidylic acid. J. Immunol. 1970;104:1355–1358. [PubMed] [Google Scholar]
- Cohen A. Protection of mice against Asian influenza virus infection by a normal horse serum inhibitor. Lancet. 1960;11:791–794. doi: 10.1016/s0140-6736(60)91859-6. [DOI] [PubMed] [Google Scholar]
- Coombs K., Mann E., Edwards J., Brown D.T. Effects of chloroquine and cytochalasin B on the infection of cells by Sindbis virus and vesicular stomatitis virus. J. Virol. 1981;37:1060–1065. doi: 10.1128/jvi.37.3.1060-1065.1981. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coppenhaver D.H., Baron J.L., McKerlie M.L., Sabados J., Baron S. Size and stability of a naturally occurring virus inhibitor. Antimicrob. Agents Chemother. 1984;25:646–649. doi: 10.1128/aac.25.5.646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coppenhaver D.H., Sriyuktasuth-Woo P., Baron S. Correlation of nonspecific antiviral activity with the ability to isolate infectious HIV-1 from saliva. New Engl. J. Med. 1994;330:1314–1315. doi: 10.1056/NEJM199405053301815. [DOI] [PubMed] [Google Scholar]
- Crombie R., Silverstein R.L., MacLow C., Pearce S.F.A., Nachman R.L., Laurence J. Identification of a CD36-related thrombospondin 1-binding domain in HIV-1 envelope glycoprotein gp120: relationship to HIV-1-specific inhibitory factors in human saliva. J. Exp. Med. 1998;187:25–35. doi: 10.1084/jem.187.1.25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Czarniecki C.W. The role of tumor necrosis factor in viral disease. Antiviral Res. 1993;22:223–258. doi: 10.1016/0166-3542(93)90035-h. [DOI] [PubMed] [Google Scholar]
- Davoli R., Bartolomei-Corsi O. Nonspecific inhibition of haemagglutination by influenza viruses. VI. Haemaglutination-inhibition neutralization of infectivity if influenza viruses of type A2 by normal guinea-pig serum activated by heat. Sperimentale. 1959;109:32–42. [Google Scholar]
- Day A.J., DuPont M.S., Ridley S., Rhodes M., Rhodes M.J., Morgan M.R., Williamson G. Deglycosylation of flavonoid and isoflavonoid glycosides by human small intestine and liver beta-glucosidase activity. FEBS Lett. 1998;436:71–75. doi: 10.1016/s0014-5793(98)01101-6. [DOI] [PubMed] [Google Scholar]
- Dianzani F., Baron S. Nonspecific Defenses. In: Baron S., editor. Medical Microbiology. The University of Texas Medical Branch at Galveston; Galveston, TX: 1996. pp. 631–640. [Google Scholar]
- Dunn D.T., Newell M.L., Ades A.E., Peckham C.S. Risk of human immunodeficiency virus type 1 transmission through breastfeeding. Lancet. 1992;340:585–588. doi: 10.1016/0140-6736(92)92115-v. [DOI] [PubMed] [Google Scholar]
- Edgar, W.M., 1992. Saliva: its secretion, composition and functions. Br. Dent. J., 305–312. [DOI] [PubMed]
- European Study Group, 1989. Risk factors for male to female transmission of HIV. BMJ 298, 411–415. [DOI] [PMC free article] [PubMed]
- European Study Group on Heterosexual Transmission of HIV, 1992. Comparison of female to male and male to female transmission of HIV in 563 stable couples. BMJ 304, 809–813. [DOI] [PMC free article] [PubMed]
- Fox, P.C., Wolff, A., Yeh, C.-K., Atkinson, J.C., Baum, B.J., 1988. Saliva inhibits HIV-1 infectivity. J. Am. Dent. Assoc. 116. [DOI] [PubMed]
- Fox P.C., Wolff A., Yeh C.-K., Atkinson J.C., Baum B.J. Salivary inhibition of HIV-1 infectivity: functional properties and distribution in men, women, and children. J. Am. Dent. Assoc. 1989;118:709–711. doi: 10.14219/jada.archive.1989.0165. [DOI] [PubMed] [Google Scholar]
- Fuhrman J.A. Marine viruses and their biogeochemical and ecological effects. Nature. 1999;399:541–548. doi: 10.1038/21119. [DOI] [PubMed] [Google Scholar]
- Fultz P.N. Components of saliva inactivate human immunodeficiency virus. Lancet. 1986;2:1215. doi: 10.1016/s0140-6736(86)92218-x. [DOI] [PubMed] [Google Scholar]
- Gerna G., Cattaneo E., Cereda P.M., Revelo M.G., Achilli G. Human coronavirus OC43 serum inhibitor and neutralizing antibody by a new plaque-reduction assay. Proc. Soc. Exp. Biol. Med. 1980;163:360–366. doi: 10.3181/00379727-163-40778. [DOI] [PubMed] [Google Scholar]
- Girgis S.M., Hafiez A.A., Mittawy B., Hamza K.H. Electrolyte concentration in semen. Andrologia. 1980;12:323–327. doi: 10.1111/j.1439-0272.1980.tb00639.x. [DOI] [PubMed] [Google Scholar]
- Goldman A.S., Goldblum R.M. Transfer of maternal leukocytes to the infant by human milk. In: Olding L.B., editor. Reproductive Immunology. Springer; Berlin: 1996. pp. 205–213. [DOI] [PubMed] [Google Scholar]
- Gooch B., Marianos D., Ciesielski C. Lack of evidence for patient-to-patient transmission of HIV in a dental practice. J. Am. Dent. Assoc. 1993;124:38–44. doi: 10.14219/jada.archive.1993.0006. [DOI] [PubMed] [Google Scholar]
- Goto Y., Yeh C.K., Notkins A.L., Prabhakar B.S. Detection of proviral sequences in saliva of patients infected with human immunodeficiency virus type 1. AIDS Res. Hum. Retroviruses. 1991;7:343–347. doi: 10.1089/aid.1991.7.343. [DOI] [PubMed] [Google Scholar]
- Griffin D.E. Therapy of viral infections of the central nervous system. A mini-review. Antiviral Res. 1991;15:1–10. doi: 10.1016/0166-3542(91)90036-q. [DOI] [PubMed] [Google Scholar]
- Helenius A., Kartenbeck J., Simons K., Fries E. On the entry of Semliki forest virus into BHK-21 cells. J. Cell Biol. 1980;84:404–420. doi: 10.1083/jcb.84.2.404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ho D.D., Byington R.E., Schooley R.T., Flynn T., Rota T.R., Hirsch M.S. Infrequency of isolation of HTLV-III virus from saliva in AIDS. New Engl. J. Med. 1985;313:1606. doi: 10.1056/NEJM198512193132512. [DOI] [PubMed] [Google Scholar]
- Hughes T.K., Blalock J.E., McKerlie M.L., Baron S. Cell-produced viral inhibitor: possible mechanism of action and chemical composition. Infect. Immun. 1981;32:454–457. doi: 10.1128/iai.32.2.454-457.1981. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jensen E.M., Force E.E., Unger J.B. Inhibitory effect of ammonium ions on influenza virus in tissue culture. Proc. Soc. Exp. Biol. Med. 1961;107:447–451. doi: 10.3181/00379727-107-26653. [DOI] [PubMed] [Google Scholar]
- Johnson A.M., Petherick A., Davidson S.J., Brettle R., Hooker M., Howard L., McLean K.A., Osborne L.E., Robertson R., Sonnex C. Transmission of HIV to heterosexual partners of infected men and women. AIDS. 1989;3:367–372. doi: 10.1097/00002030-198906000-00005. [DOI] [PubMed] [Google Scholar]
- Johnson M.C., Maxwell J.M., Loh P.C., Leong J.A. Molecular characterization of the glycoproteins from two warm water rhabdoviruses: snakehead rhabdovirus (SHRV) and rhabdovirus of penaeid shrimp (RPS)/spring viremia of carp virus (SVCV) Virus Res. 1999;64:95–106. doi: 10.1016/s0168-1702(99)00071-4. [DOI] [PubMed] [Google Scholar]
- Karzon D.T. Non-specific viral inactivating substance (VIS) in human and mammalian sera. J. Immunol. 1956;76:454–463. [PubMed] [Google Scholar]
- Kavanagh J. Sodium, potassium, calcium, magnesium, zinc, citrate and chloride content of human prostatic and seminal fluid. J. Reprod. Fertil. 1985;75:35–41. doi: 10.1530/jrf.0.0750035. [DOI] [PubMed] [Google Scholar]
- Keet I.P., Albrecht van Lent N., Sandfort T.G., Countinho R.A., van Griensven G.J. Orogenital sex and the transmission of HIV among homosexual men. AIDS. 1992;6:223–226. doi: 10.1097/00002030-199202000-00014. [DOI] [PubMed] [Google Scholar]
- Kitamura T., Tanaka Y., Sugane M. Studies on a heat-labile variola virus inhibitor in normal sera. II. Further characterization of the inhibitor and its activity. Intervirology. 1973;1:288–296. doi: 10.1159/000148857. [DOI] [PubMed] [Google Scholar]
- Kriznova O., Rathova V. Serum inhibitors of myxoviruses. Curr. Top. Microbiol. Immunol. 1969;47:125–151. doi: 10.1007/978-3-642-46160-6_6. [DOI] [PubMed] [Google Scholar]
- Kumar S., McKerlie M.L., Albrecht T.B., Goldman A.S., Baron S. A broadly active viral inhibitor in human and animal organ extracts and body fluids (41918) Proc. Soc. Exp. Biol. Med. 1984;177:104–111. doi: 10.3181/00379727-177-41918. [DOI] [PubMed] [Google Scholar]
- Lane H.C., Holmberg S.D., Jaffe H.W. HIV seroconversion and oral intercourse (letter) Am. J. Public Health. 1991;81:658. doi: 10.2105/ajph.81.5.658. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lazzarin A., Saracco A., Musicco M., Nicolosi A. Man-to-woman sexual transmission of the human immunodeficiency virus. Risk factors related to sexual behavior, man's infectiousness, and woman's susceptibility. Italian Study Group on HIV Heterosexual Transmission. Arch. Intern. Med. 1991;151:2411–2416. [PubMed] [Google Scholar]
- Lee-Huang S., Huang P.L., Sun Y., Huang P.L., Kung H.-F., Blithe D.L., Chen H.-C. Lysozyme and RNases as anti-HIV components in β-core preparation of human chorionic gonadotropin. Proc. Natl. Acad. Sci. USA. 1999;96:2678–2681. doi: 10.1073/pnas.96.6.2678. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Levy J.A. The transmission of AIDS: the case of the infected cell. JAMA. 1988;260:3586–3587. [PubMed] [Google Scholar]
- Levy J., Greenspan D. HIV in saliva. Lancet. 1988;2:1248. doi: 10.1016/s0140-6736(88)90835-5. [DOI] [PubMed] [Google Scholar]
- Lifson A.R., O'Malley P.M., Hessol N.A., Buchbinder S.P., Cannon L., Ritherford G.W. HIV seroconversion in two homosexual men after receptive oral intercourse with ejaculation: implications for counseling concerning safe sexual practices. Am. J. Public Health. 1990;80:1509–1511. doi: 10.2105/ajph.80.12.1509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Link F., Blaskovic D., Raus J. On the therapeutic effect of gamma inhibitor in mice infected with an inhibitor-sensitive unadapted A2 influenza virus. Acta Virol. 1965;9:553. [PubMed] [Google Scholar]
- Link F., Szanto J., Kriznova O. A quantitative assay of the in vivo protective effect of gamma inhibitor against inhibitor sensitive A2 influenza virus. Acta Virol. 1964;8:71–75. [PubMed] [Google Scholar]
- Marsh M., Helenius A. Adsorptive endocytosis of Semliki Forest virus. J. Mol. Biol. 1980;142:439–454. doi: 10.1016/0022-2836(80)90281-8. [DOI] [PubMed] [Google Scholar]
- Mayer K.H., DeGruttola V. Human immunodeficiency virus and oral intercourse. Ann. Intern. Med. 1987;107:428–429. doi: 10.7326/0003-4819-107-2-428_2. [DOI] [PubMed] [Google Scholar]
- McCumber L.J., Clem L.W. Recognition of viruses and xenogeneic proteins by the blue crab, Callinectes sapidus. I. Clearance and organ concentration. Dev. Comp. Immunol. 1977;1:5–14. doi: 10.1016/s0145-305x(77)80045-1. [DOI] [PubMed] [Google Scholar]
- McNeely T.B., Dealy M., Dripps D.J., Orenstein J.M., Eisenberg S.P., Wahl S.M. Secretory leukocyte protease inhibitor: a human saliva protein exhibiting anti-human immunodeficiency virus 1 activity in vitro. J. Clin. Invest. 1995;96:456–464. doi: 10.1172/JCI118056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McNeely T., Shugars D.M.R., Tucker C., Eisenberg S., Wahl S.M. Inhibition of human immunodeficiency virus type 1 infectivity by secretory leukocyte protease inhibitor occurs prior to viral reverse transcription. Blood. 1997;90:1141–1149. [PubMed] [Google Scholar]
- Mehl M. Change in sodium and potassium concentration in human seminal plasma infected experimentally with ureaplasma urealyticum. Andrologia. 1986;18:224–229. [PubMed] [Google Scholar]
- Melvin A.J., Tamura G.S., House J.K., Hobson A.C., Cone R.W., Frenkel L.M., Burchett S. Lack of detection of human immunodeficiency virus type 1 in the saliva of infected children and adolescents. Arch. Pediatr. Adolesc. Med. 1997;151:228–232. doi: 10.1001/archpedi.1997.02170400014003. [DOI] [PubMed] [Google Scholar]
- Milman G., Sharma O. Mechanisms of HIV/SIV mucosal transmission. AIDS Res. Hum. Retroviruses. 1994;10:1305–1312. doi: 10.1089/aid.1994.10.1305. [DOI] [PubMed] [Google Scholar]
- Mohor J.A. The possible induction of cellular hypersensitivity associated with ingestion of colostrum. J. Pediatr. 1973;82:1062–1064. doi: 10.1016/s0022-3476(73)80448-2. [DOI] [PubMed] [Google Scholar]
- Moore B.E., Flaitz C.M., Coppenhaver D.H., Nichols C.M., Kalmaz G.D., Bessman J.D., Cloyd M.W., Lynch D.P., Prabhakar B.S., Baron S. HIV recovery from saliva before and after dental treatment: Inhibitors may have a critical role in viral inactivation. J. Am. Dent. Assoc. 1993;124:67–74. doi: 10.14219/jada.archive.1993.0197. [DOI] [PubMed] [Google Scholar]
- Nahmias A.J., Kibrick S. Inhibitory effect of heparin on herpes simplex virus. J. Bacteriol. 1964;87:1060–1066. doi: 10.1128/jb.87.5.1060-1066.1964. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Norkin L.C., Einck K.H. Cell killing by Simian virus 40: protective effect of chloroquine. Antimicrob. Agents Chemother. 1978;14:930–932. doi: 10.1128/aac.14.6.930. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Owens B.L., Ananthramaiah G.M., Kahlon J.B., Srinivas R.V., Compans R.W., Segrest J.P. Apoliprotein A-1 and its amphipathic helix peptide analogues inhibit human immunodeficiency virus-induced syncitium formation. J. Clin. Invest. 1990;4:1142–1150. doi: 10.1172/JCI114819. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pan J., Kurosky A., Xu B., Chopra A., Coppenhaver D.H., Singh I.P., Baron S. Broad antiviral activity in tissues of crustaceans. Antiviral Res. 2000;48:39–47. doi: 10.1016/s0166-3542(00)00117-0. [DOI] [PubMed] [Google Scholar]
- Pauza C.D., Emau P., Salvato M.S., Trivedi P., MacKenzie D., Malkovsky M., Uno H., Schultz K. Pathogenesis of SIVmac251 after a traumatic inoculation of the rectal mucosa in rhesus monkeys. J. Med. Primatol. 1993;22:154–161. [PubMed] [Google Scholar]
- Pauza C.D., Malkovsky M., Salvato M.S. SIV transmission across the mucosal barrier. AIDS Res. Hum. Retroviruses. 1994;10:S7–S10. [PubMed] [Google Scholar]
- Phillips D.M., Bourinbaiar A.S. Mechanism of HIV spread from lymphocytes to epithelia. Virology. 1992;186:261–273. doi: 10.1016/0042-6822(92)90080-9. [DOI] [PubMed] [Google Scholar]
- Piazza M., Chirianni A., Picciotto L., Cataldo P.T., Borgia G., Orlando R. Blood in saliva of patients with acquired immunodeficiency syndrome: possible implication in sexual transmission of the disease. J. Med. Virol. 1994;42:38–41. doi: 10.1002/jmv.1890420108. [DOI] [PubMed] [Google Scholar]
- Podolsky D.K. Mechanisms of regulatory peptide action in the gastrointestinal tract: trefoil peptides [In Process Citation] J. Gastroenterol. 2000;35:69–74. [PubMed] [Google Scholar]
- Quatro M., Germinario C., Troiano T. HIV transmission by fellatio. Eur. J. Epidemiol. 1990;6:339–340. doi: 10.1007/BF00150446. [DOI] [PubMed] [Google Scholar]
- Qureshi M.N., Barr C.E., Seshamma T., Reidy J., Pomerantz R.J., Bagasra O. Infection of oral mucosal cells by human immunodeficiency virus type 1 in seropositive persons. J. Infect. Dis. 1995;171:190–193. doi: 10.1093/infdis/171.1.190. [DOI] [PubMed] [Google Scholar]
- Richman K.M., Rickman L.S. The potential for transmission of human immunodeficiency virus through human bites. J. Acquir. Immune Defic. Syndr. 1993;6:402–406. [PubMed] [Google Scholar]
- Rider C.C. The potential for heparin and its derivatives in the therapy and prevention of HIV-1 infection. Glycoconj. J. 1997;14:639–642. doi: 10.1023/a:1018596728605. [DOI] [PubMed] [Google Scholar]
- Robinovitch M.R., Iversen J.M., Resnick L. Anti-infectivity activity of human salivary secretions toward human immunodeficiency virus. Crit. Rev. Oral Biol. Med. 1993;4:455–459. doi: 10.1177/10454411930040032801. [DOI] [PubMed] [Google Scholar]
- Rogers, M.F., White, C.R., Sanders, R., et al., 1990. Lack of transmission of human immunodeficiency virus from infected children to their household contacts. Pediatrics 85. [PubMed]
- Royce R.A., Sena A., Cates W., Jr, Cohen M.S. Sexual transmission of HIV. New Engl. J. Med. 1997;336:1072–1078. doi: 10.1056/NEJM199704103361507. [DOI] [PubMed] [Google Scholar]
- Ryder R.W., Manzila T., Baende E., Kabagabo U., Behets F., Batter V., Paquot E., Binyingo E., Heyward W.L. Evidence from Zaire that breast-feeding by HIV-1-seropositive mothers is not a major route for perinatal HIV-1 transmission but does decrease morbidity. AIDS. 1991;5:709–714. doi: 10.1097/00002030-199106000-00010. [DOI] [PubMed] [Google Scholar]
- Sambhi S.K., Kohonen-Corish M.R., Ramshaw I.A. Local production of tumor necrosis factory encoded by recombinant vaccinia virus is effective in controlling virus replication in vivo. Proc. Natl. Acad. Sci. USA. 1991;88:4025–4029. doi: 10.1073/pnas.88.9.4025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schapiro H.C. Immunity in decapod crustaceans. Am. Zool. 1975;15:13–19. [Google Scholar]
- Schnapp D., Kemp G.D., Smith V.J. Purification and characterization of a proline-rich antibacterial peptide, with sequence similarity to bactenecin-7, from the haemocytes of the shore crab, Carcinus maenas. Eur. J. Biochem. 1996;240:532–539. doi: 10.1111/j.1432-1033.1996.0532h.x. [DOI] [PubMed] [Google Scholar]
- Seidel K.M., Goyal S.M., Rao V.C., Melnick J.L. Concentration of rotavirus and enteroviruses from blue crabs (Callinectes sapidus) Appl. Environ. Microbiol. 1983;46:1293–1296. doi: 10.1128/aem.46.6.1293-1296.1983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shen W.H., Xu R.J. Stability of insulin-like growth factor I in the gastrointestinal lumen in neonatal pigs. J. Pediatr. Gastroenterol. Nutr. 2000;30:299–304. doi: 10.1097/00005176-200003000-00016. [DOI] [PubMed] [Google Scholar]
- Shortridge K.F., Ho W.K. Human serum lipoproteins as inhibitors of haemagglutination for selected togaviruses. J. Gen. Virol. 1974;23:113–116. doi: 10.1099/0022-1317-23-1-113. [DOI] [PubMed] [Google Scholar]
- Shugars D.C. Endogenous mucosal antiviral factors of the oral cavity. J. Infect. Dis. 1999;179:S431–S435. doi: 10.1086/314799. [DOI] [PubMed] [Google Scholar]
- Singh, I.P., Coppenhaver, D.H., Chopra, A.K., Baron, S., 2000. Innate gastrointestinal immunity: Characterization of broadly active viral inhibitors. Antiviral Res., submitted. [DOI] [PMC free article] [PubMed]
- Singh I.P., Coppenhaver D.H., Sarzotti M., Sriyuktasuth P., Poast J., Levy H.B., Baron S. Postinfection therapy of arbovirus infections in mice. Antimicrob. Agents Chemother. 1989;33:2126–2131. doi: 10.1128/aac.33.12.2126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Singh I.P., Chopra A.K., Coppenhaver D.H., Smith E., Poast J., Baron S. Vertebrate brains contain a broadly active antiviral substance. Antiviral Res. 1995;27:375–388. doi: 10.1016/0166-3542(95)00021-d. [DOI] [PubMed] [Google Scholar]
- Singh I.P., Coppenhaver D.H., Chopra A.K., Baron S. Further characterization of a broad-spectrum antiviral substance in human serum. Viral Immunol. 1992;5:293–303. doi: 10.1089/vim.1992.5.293. [DOI] [PubMed] [Google Scholar]
- Singh I.P., Coppenhaver D.H., Chopra A.K., Baron S. Generalized occurrence of the broadly antiviral substance UTIβ in mammalian sera. J. Biol. Regul. Homeost. Agents. 1993;7:7–14. [PubMed] [Google Scholar]
- Singh I.P., Chopra A.K., Coppenhaver D.H., Ananatharamaiah G.M., Baron S. Lipoproteins account for part of the broad non-specific antiviral activity of human serum. Antiviral Res. 1999;42:211–218. doi: 10.1016/s0166-3542(99)00032-7. [DOI] [PubMed] [Google Scholar]
- Smorodintsev A.A. Factors of natural resistance and specific immunity to viruses. Virology. 1957;3:299–321. doi: 10.1016/0042-6822(57)90095-8. [DOI] [PubMed] [Google Scholar]
- Smorodintsev A.A. Basic mechanism of nonspecific resistance to viruses in animals and man. Adv. Virus Res. 1960;7:327–376. [Google Scholar]
- Srinivas R.V., Birkedal B., Owens R.J., Anantharamaiah G.M., Segrest J.P., Compans R.W. Antiviral effects of apolipoprotein A-I and its synthetic amphipathic peptide analogs. Virology. 1990;176:48–57. doi: 10.1016/0042-6822(90)90229-k. [DOI] [PubMed] [Google Scholar]
- Sullivan M.L., Niesel D.W., Coppenhaver D.H., Sabados J., Baron S. Characterization of an antiviral agent from primary murine fibroblast cultures: murine tissue culture CVI. J. Biol. Regul. Homeost. Agents. 1987;1:126–132. [PubMed] [Google Scholar]
- Sun D., Archibald D.W., Furth P.A. Variation of secretory antibodies in parotid saliva to human immunodeficiency virus type 1 with HIV-1 disease stage. AIDS Res. Hum. Retroviruses. 1990;6:933–941. doi: 10.1089/aid.1990.6.933. [DOI] [PubMed] [Google Scholar]
- Suribaldi L., Seganti L., Orise N., Stasia A., Valenti P. Microbiologics. 1979;2:121–128. [Google Scholar]
- Tamm I., Horsfall F.L., Jr A mucoprotein derived from human urine which reacts with influenza, mumps, and Newcastle disease. J. Exp. Med. 1952;95:71–97. doi: 10.1084/jem.95.1.71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thiry L., Cogniaux-Le Clerc J., Content J., Tack L. Factors which influence inactivation of vesicular stomatitis virus by fresh human serum. Virology. 1978;87:384–393. doi: 10.1016/0042-6822(78)90142-3. [DOI] [PubMed] [Google Scholar]
- Tietz N.W. Vol. 52. Saunders; Philadelphia, PA: 1983. (Clinical Guide to Laboratory Tests). [Google Scholar]
- Utz J.P. Viuria in man. Prog. Med. Virol. 1964;6:71–81. [PubMed] [Google Scholar]
- Vernazza P.L., Eron J.J., Fiscus S.A. Sensitive method for the detection of infectious HIV in semen of seropositive individuals. J. Virol. Methods. 1996;56:33–40. doi: 10.1016/0166-0934(95)01899-9. [DOI] [PubMed] [Google Scholar]
- Vonesch N., Sturchio E., Humani A.C., Fei P.C., Cosenza D., Caprilli F., Pezzella M. Detection of HIV-1 genome in leukocytes of human colostrum from anti-HIV-1 seropositive mothers. AIDS Res. Hum. Retroviruses. 1992;8:1283–1287. doi: 10.1089/aid.1992.8.1283. [DOI] [PubMed] [Google Scholar]
- Wahl S.M., Orenstein J.M. Immune stimulation and HIV-1 viral replication. J. Leukocyte Biol. 1997;62:67–71. doi: 10.1002/jlb.62.1.67. [DOI] [PubMed] [Google Scholar]
- Wahl S.M., Worley P., Jin W., McNeely T.B., Eisenberg S., Fasching C., Orenstein J.M., Janoff E.N. Anatomic dissociation between HIV-1 and its endogenous inhibitor in mucosal tissues. Am. J. Pathol. 1997;150:1275–1284. [PMC free article] [PubMed] [Google Scholar]
- Wallace M.R., Woelfl I., Bowler W.A., Olson P.E., Murray N.B., Brodine S.K., Goldfield E.C., 3rd, Arvin A.M. Tumor necrosis factor, interleukin-2, and interferon-gamma in adult varcella. J. Med. Virol. 1994;43:69–71. doi: 10.1002/jmv.1890430113. [DOI] [PubMed] [Google Scholar]
- Welsh R.M., Jr, Jensen F.C., Cooper N.R., Oldstone M.B. Inactivation of lysis of oncornaviruses by human serum. Virology. 1976;74:432–440. doi: 10.1016/0042-6822(76)90349-4. [DOI] [PubMed] [Google Scholar]
- Witas H., Sarosiek J., Aono M., Murty V.L., Slomiany A., Slomiany B.L. Lipids associated with rat small-intestinal mucus glycoprotein. Carbohydr. Res. 1983;120:67–76. doi: 10.1016/0008-6215(83)88007-0. [DOI] [PubMed] [Google Scholar]
- Yeh C.K., Handelman B., Fox P.C., Baum B.J. Further studies of salivary inhibition of HIV-1 infectivity. J. Acquir. Immune Defic. Syndr. 1992;5:898–903. [PubMed] [Google Scholar]
- Yeung S.C.H., Kazazi F., Randle C.G.M. Patients infected with human immunodeficiency virus type 1 have low levels of virus in saliva even in the presence of periodontal disease. J. Infect. Dis. 1993;167:803–809. doi: 10.1093/infdis/167.4.803. [DOI] [PubMed] [Google Scholar]
- Zacharopoulos V.R., Phillips D.M. Cell-mediated HTLV-1 infection of a cervix-derived epithelial cell line. Microb. Pathog. 1997;23:225–233. doi: 10.1006/mpat.1997.0151. [DOI] [PubMed] [Google Scholar]
- Zhu T., Wang N., Carr A., Nam D.S., Moor-Jankowski R., Cooper D.A., Ho D.D. Genetic characterization of human immunodeficiency virus type 1 in blood and genital secretions: evidence for viral compartmentalization and selection during sexual transmission. J. Virol. 1996;70:3098–3107. doi: 10.1128/jvi.70.5.3098-3107.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zijlstra A.I., Offner G.D., Afdhal N.H., van Overveld M., Tytgat G.N., Groen A.K. The pronase resistance of cholesterol-nucleating glycoproteins in human bile. Gastroenterology. 1996;110:1926–1935. doi: 10.1053/gast.1996.v110.pm8964420. [DOI] [PubMed] [Google Scholar]