

Abstract
With the growing emergence of pan-drug-resistant Acinetobacter baumannii (PDR-Ab) strains in clinical, new strategies for the treatment of PDR-Ab infections are urgently needed. Egg yolk immunoglobulin (IgY) as a convenient and inexpensive antibody has been widely applied to the therapy of infectious diseases. The aim of this study was to produce IgY specific to PDR-Ab and investigate its antibacterial effects in vitro and in vivo. IgYs specific to two PDR-Ab strains were produced by immunizing hens with formaldehyde inactivated PDR-Ab cells and isolated from yolks with a purity of 90% by water dilution, salt precipitations and ultrafiltration. IgYs showed high titers when subjected to an ELISA and inhibited the growth of PDR-Ab in a dose-dependent manner in liquid medium. Scanning electron microscopy assay showed structural modification and aggregation of PDR-Ab treated with specific IgYs. Freshly cultured PDR-Ab cells were nasally inhaled in BALB/c mice to induce acute pneumonia. The infected mice were intraperitoneally injected with specific IgYs using cefoperazone/sulbactam and dexamethasone as positive controls. The IgYs specific to PDR-Ab lowered the mortality of mice with PDR-Ab-induced acute pneumonia, decreased the level of TNF-α and IL-1β in serum and reduced inflammation in lung tissue. Specific IgY has the potential to be used as a new therapeutic approach for the treatment of A. baumannii-induced infections.
Keywords: A. baumannii, Pan-drug-resistance, Egg yolk immunoglobulin, Acute pneumonia
1. Introduction
Acinetobacter baumannii (A. baumannii) is one of the most common nosocomial acinetobacter and belongs to the non-fermentative Gram-negative bacilli. A. baumannii is responsible for many kinds of nosocomial infections, such as hospital acquired sepsis, pneumonia especially ventilator-associated pneumonia (VAP), skin and soft-tissue infections, wound infections, urinary tract infections, secondary meningitis and bloodstream infections [1], [2], [3]. Over the past 10 years, nosocomial outbreaks due to A. baumannii have been reported increasingly. This caused great threat to patients in ICU and increased mortality, hospital stay and medicine costs [3].
With the wide use of broad-spectrum antibacterials, most strains of A. baumannii have been resistant to multiple antimicrobial agents [4]. Infections caused by A. baumannii are difficult to cure because its multiple intrinsic and acquired mechanisms make the bacteria frequently multidrug-resistant (MDR) to antimicrobials [5]. More seriously, pan-drug-resistant A. baumannii (PDR-Ab), defined as resistance to nearly all routinely tested antimicrobials [6], were increasingly reported. Little drug is applicable when face to PDR-Ab infections.
Antibacterial resistance is a ubiquitous and relentless clinical problem that is compounded by a lack of new therapeutic agents [7]. The predicament of new antibacterial development in pharmaceutical industry promotes the innovation of antimicrobial therapy in biomedical field such as the use of antibodies.
Egg yolk immunoglobulin (IgY) is antibody transferred from serum to egg yolk of poultry in order to protect the developing embryo from potential pathogens. Using antigen-specific immunity response, a mass of specific IgY can be obtained from egg yolks of laying hens. Recently, IgY has attracted considerable interest as therapeutic and diagnostic agent. Up to now, specific IgYs have been widely applied to the treatment of infectious diseases caused by various kinds of bacteria, including Escherichia coli [8], [9], Helicobacter pylori [10], Salmonella [11], Staphylococcus aureus [12], [13], Streptococcus mutants [14], Yersinia ruckeri [15]. Infections caused by virus, such as rotavirus [16], coronavirus [17] and enterovirus [18], could also be interfered by specific IgYs. Even so, IgY specific to A. baumannii has not been reported.
Different from traditional antibacterials, specific IgY is environmentally-friendly and does not elicit undesirable side effects, drug resistance or toxic residues [19], [20]. In addition, IgY possesses a large number of advantages over mammalian IgG such as cost-effectiveness, convenience, and high yield [20], [21].
In this study, we prepared two IgYs respectively against two PDR-Ab strains of different sources, and estimate their antibacterial activities in vitro and in vivo. This will provide reference for using IgY in the treatment of PDR-Ab infections.
2. Materials and methods
2.1. Bacteria and animals
A. baumannii ATCC BAA1605, named as Ab-B in this study, was obtained from the American type culture collection (ATCC). Clinically isolated strain of A. baumannii, named as Ab-C, was isolated from the sputum of patients in Dalian Friendship Hospital. These two strains have been identified as PDR-Ab by drug sensitivity test in our previous study [22]. Hens used for immunization were 140-day-old Hy-line laying hens housed at a commercial poultry facility in Dalian, China. Male BALB/c mice, weighing approximately 18–22 g, were obtained from the Animal Care Center of Dalian Medical University (Dalian, China). Before the experiment, the mice had 2–3 days to adapt to their experimental environment [23]. All experiments were performed according to the Experimental Animal Management Law of China and approved by the Animal Ethics Committee of Dalian Medical University.
2.2. Production of IgYs against PDR-Ab
PDR-Ab strains Ab-B and Ab-C were respectively used as antigens for IgY production. Ab-B or Ab-C were cultured in Tryptic Soy Broth (TSB) media for 24 h and collected in PBS at a concentration of 109 cfu/ml and then inactivated with 0.7% (w/v) formaldehyde for 24 h. The inactivated antigen suspensions were emulsified with an equal volume of complete and incomplete Freund’s adjuvant for the first and subsequent booster immunizations, respectively. Hens were immunized according to reported methods [24]. Fifteen Hy-line laying hens were divided into three groups and immunized with the two vaccines and PBS, respectively. Hens were injected intramuscularly at three sites with a total volume of 1.5 ml for the first injection and 2.0 ml for two booster immunizations at 2-week intervals. Eggs were collected daily from the immunized hens after the first injection and were stored at 4 °C before use [13].
2.3. Isolation and purification of IgYs
Egg yolks were diluted with six volumes of distilled water and the pH was adjusted to 5.0 [25]. The suspensions were stored overnight at 4 °C and clarified by centrifugation (10,000 × g). The water soluble fractions (WSF) were collected and then successively precipitated with 60% saturated ammonium sulfate and 14% (w/v) sodium sulfate. The precipitates were resuspended with distilled water and ultrafiltered with a Vivaflow 50 Tangential Flow Ultrafilter (Vivascience, Hannover, Germany) with a 100-kDa cutoff membrane, and lyophilized to obtain the IgY powder.
The purity of IgY obtained from different phases of isolation was assessed by means of sodium dodecyl sulfate polyacryl amide gel electrophoresis (SDS-PAGE). Standard IgY (purity >90%, Promega, Madison, WI) was used as a reference. SDS-PAGE was carried out under non-reducing conditions with an 8% gel on Tris-Glycine System and stained with Coomassie brilliant blue according to the manufacturer’s recommendations [26].
2.4. IgY titer analysis
Titer of IgYs specific to Ab-B (B-IgY) and Ab-C (C-IgY) was determined by enzyme-linked immunosorbent assay (ELISA) conducted on WSF fractions [27]. Microtiter plate was coated with Ab-B or Ab-C at a concentration of 109 cfu/ml. The WSF fractions were firstly diluted 1:100 in PBS, then serially two-fold diluted. WSF fraction (1:2000 dilution) from the eggs of non-immunized hens, serum (1:2000 dilution) from immunized hens and PBS were used as negative, positive and blank controls, respectively. Horseradish peroxidase coupled rabbit anti-chicken IgG (Sigma-Aldrich, St. Louis, USA, 1:30,000 dilution) was used as secondary antibody. The reaction was stopped by 2 M H2SO4 (50 ml/well). All the other reagents were added at a volume of 100 ml/well. Optical density (OD) at 450 nm was read on microplate reader, using 630 nm as a reference wavelength. While ODsample/ODnegative ≥2.1, the corresponding dilution multiple of the sample was determined as IgY titer.
2.5. Growth inhibition of specific IgYs to PDR-Ab
Lyophilized B-IgY and C-IgY powder (titer 12800) was dissolved in TSB liquid medium at concentrations of 5, 10 and 20 mg/ml and nonspecific IgY powder was dissolved in the same medium at a concentration of 10 mg/ml. All solutions were 0.22 μm-filter sterilized, in which freshly cultured Ab-B and Ab-C cells were respectively added at a final concentration of 106 cfu/ml. TSB liquid medium was used as blank control, the medium containing PDR-Ab was used as negative control, and cefoperazone/sulbactam (Pfizer, America) at a concentration of 5 mg/ml was used as positive control. Mixtures of bacteria and IgYs were incubated at 37 °C with shaking at 100 rpm. OD600 nm values of the suspensions were measured by a microplate reader (Thermo Fisher Scientific, Waltham, MA, USA) at 2 h intervals until that of the blank control (i.e., E. coli growth curve) reached stationary phase.
2.6. Scanning electron microscopy (SEM) assay
Lyophilized B-IgY and C-IgY powder (titer 12800) were dissolved in PBS containing 1% BSA to a concentration of 5 mg/ml. Nonspecific IgY at the same concentration was used as control. After the solutions were 0.22 μm-filter sterilized, Ab-B and Ab-C were added at a final concentration of 109 cfu/ml. All suspensions were incubated at 37 °C for 2 h and centrifuged at 4000 rpm for 5 min. The sediments were washed with PBS twice and prepared as specimens for SEM assay and then observed using an electron microscope (FEI, America).
2.7. Death prevention of specific IgYs to mice with PDR-Ab-induced acute pneumonia
Seventy-two mice were divided into six groups. One group was used as blank control, others were modeled as acute pneumonia. Clinically isolated strain Ab-C was used to induce acute pneumonia in mice. Freshly cultured Ab-C cells (109 cfu/ml) were nasally inhaled in BALB/c mice at a dose of 100 μl. Acute pneumonia in mice was determined by status observation and histopathological examination of lung tissue. The powder of specific IgYs (titer 12,800) was dissolved in 0.9% NaCl to 20 mg/ml and sterilized with 0.22 μm filter. 20 mg/ml of cefoperazone/sulbactam and 0.5 mg/ml of dexamethasone (Shuanghe Pharmacy Company, China) were used as positive controls, and 0.9% NaCl was used as negative control. Mice received intraperitoneal injection of the agents at a dose of 250 mg/kg except for dexamethasone at 2.5 mg/kg 12 h before PDR-Ab challenge and then received one more injection at double dose 1 h after infection. The status of the mice was observed and the mortality within 24 h was recorded.
2.8. Effects of specific IgYs on cytokines in PDR-Ab-infected mice
Eighteen BALB/c mice were divided into six groups. One group was used as blank control, others were nasally infected with freshly cultured Ab-C cells (107 cfu/ml) at a dose of 100 μl. The infected mice were treated with IgYs and positive agents as described above. Blood was collected from the mice through retro-orbital puncture at 0, 2, 4 and 6 h of infection. Serum samples were prepared and stored at −80 °C. The levels of tumor necrosis factor-α (TNF-α) and interleukin-1β (IL-1β) in serum were detected by commercial ELISA kits (Jingmei Biotech, Beijing, China).
2.9. Histopathological detection
Six hours after blood collection, the infected mice were killed for histopathological detection. Lung tissue of the mice was fixed in neutral buffered formalin, embedded in paraffin, sectioned at 5 μm and stained with hematoxylin and eosin (H & E). Stained tissue sections were examined microscopically by a scientist who did not know what treatment the experimental animals were assigned to [28].
2.10. Statistical analysis
Statistical analysis was conducted using SPSS 19.0 for Windows (SPSS Inc., Chicago, IL, USA). Data were represented as means ± standard deviation (SD) and levels of significance were evaluated using LSD test. The differences were considered significant at the level of p < 0.05.
3. Results
3.1. Isolation and purification of IgYs
IgYs were isolated from the yolks of immunized hens by water dilution and purified by salt precipitations and ultrafiltration. IgY has a molecular weight of 180 kDa as visualized in Fig. 1 . The purity of IgY increased significantly after sodium sulfate precipitation and ultrafiltration as documented by SDS-PAGE. The electrophoretic pattern of ultrafiltration fractions was in line with that of the standard IgY (purity >90%).
Fig. 1.

SDS-PAGE of the IgY purification process. Lines: 1, molecular weight marker; 2, water soluble fraction of B-IgY; 3, ammonium sulfate salting-out fraction of B-IgY; 4, sodium sulfate salting-out fraction of B-IgY; 5, ultrafiltration fraction of B-IgY; 6, water soluble fraction of C-IgY; 7, ammonium sulfate salting-out fraction of C-IgY; 8, sodium sulfate salting-out fraction of C-IgY; 9, ultrafiltration fraction of C-IgY; 10, standard IgY.
3.2. Titer of IgYs
ELISA revealed the ability of the two specific IgYs binding with A. baumannii and showed the pattern of the immune response by the hens over 75 days. The titer of B-IgY produced by Ab-B-immunized hens increased after 10 days of initial immunization and reach to 12,800 at day 25. The highest titer (25,600) was reached on day 30 and maintained to day 55 (Fig. 2 ). The titer of C-IgY produced by Ab-C-immunized hens increased after 15 days of initial immunization and reach to 12,800 at day 30. The highest titer (25,600) was reached on day 35 and maintained to day 60 (Fig. 2). On the contrary, the titer of IgY produced by hens immunized with PBS was extremely low.
Fig. 2.

Titer of the IgYs specific to Ab-B and Ab-C. Titer of IgY is presented by dilution times of water soluble fraction.
3.3. Growth inhibition of specific IgYs to PDR-Ab
Both of the two IgYs could inhibit the growth of the two antigen strains in liquid medium in a dose-dependent manner at concentrations ranging from 5 mg/ml to 20 mg/ml (Fig. 3 ) while nonspecific IgY did not show significant growth inhibition to the bacteria. However, the inhibition effect of both B-IgY and C-IgY to the clinically isolated strain Ab-C is lower than that to Ab-B. In a whole, the growth of PDR-Ab could be inhibited by specific IgYs with different extent, but could hardly be inhibited by nonspecific IgY.
Fig. 3.

Growth inhibitory activity of the specific IgYs to Ab-B and Ab-C. Ab-B or Ab-C (106 cfu/ml) were respectively cultured with B-IgY or C-gY (5, 10 and 20 mg/ml), nonspecific IgY (10 mg/ml) and cefoperazone/sulbactam (5 mg/ml) in TSB liquid medium. OD600 nm values were measured at 2-h intervals. Data are presented as means ± SD (n = 3).
3.4. SEM assay
SEM assay showed that the morphology of A. baumannii changed greatly after the bacteria were treated by specific IgYs (Fig. 4, Fig. 5 ). Crenation, structural modification and aggregation could be observed on the two PDR-Ab strains treated by specific IgYs. In contrast, bacteria grown with nonspecific IgY or alone were viewed dispersive, smooth and integrally.
Fig. 4.

Scanning electron microscopy of combination of IgYs with Ab-B. A,E. Ab-B grew alone; B,F. Ab-B grew with nonspecific IgY; C,G. Ab-B grew with specific B-IgY; D,H. Ab-B grew with specific C-IgY. Magnification for A–D was 40,000; magnification for E–H was 160,000.
Fig. 5.

Scanning electron microscopy of combination of IgY with Ab-C. A,E. Ab-C grew alone; B,F. Ab-C grew with nonspecific IgY; C,G. Ab-C grew with specific B-IgY; D,H. Ab-C grew with specific C-IgY. Magnification for A–D was 40,000; magnification for E–H was 160,000.
3.5. Death prevention of specific IgYs to mice with PDR-Ab-induced acute pneumonia
Clustering, piloerection, hypothermia, tachypnea could be observed on Ab-C infected mice. Histopathological detection showed inflammatory cell infiltration in lung tissues (Fig. 6 B) while little change was found in liver tissue (Supplementary Fig. S1), suggesting the model in this study was acute pneumonia.
Liver histopathology of A. baumannii-infected mice received different treatments. A. blank group: liver tissue from healthy mice; B. negative group: liver tissue from infected mice which were treated with 0.9% NaCl;C. cefoperazone/sulbactam group: liver tissue from infected mice which were treated with cefoperazone /sulbactam; D. dexamethasone group: liver tissue from infected mice which were treated with dexamethasone;E. B-IgY group: liver tissue from infected mice which were treated with B-IgY; F. C-IgY group: liver tissue from infected mice which were treated with B-IgY.
{kind=link}
Fig. 6.

Lung histopathology of A. baumannii-infected mice received different treatments. A. blank group: lung tissue from normal mice; B. negative group: lung tissue from infected mice which were treated with 0.9% NaCl; C. cefoperazone/sulbactam group: lung tissue from infected mice which were treated with cefoperazone/sulbactam; D. dexamethasone group: lung tissue from infected mice which were treated with dexamethasone; E. B-IgY group: lung tissue from infected mice which were treated with B-IgY; F. C-IgY group: lung tissue from infected mice which were treated with B-IgY.
Mice with acute pneumonia were treated by specific IgYs, antibacterial and anti-inflammatory agent, respectively. As showed in Table 1 , death did not happen in uninfected mice within 24 h. All death of mice in other groups occurred within 12 h of infection. Mortalities of mice in negative control group and dexamethasone-treated group were both 91.7% while that in cefoperazone/sulbactam-treated group was 33.3%. The mortality decreased to 8.3% when the mice were protected by the two specific IgYs individually. The ability of the two specific IgYs to prevent mice from PDR-Ab-induced death was stronger than that of cefoperazone/sulbactam and dexamethasone.
Table 1.
Mortality of Ab-C infected mice after different treatment.
| Treatment | 0 h | 2 h | 4 h | 6 h | 8 h | 10 h | 12 h | 24 h | Mortality |
|---|---|---|---|---|---|---|---|---|---|
| Blank control | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Negative control | 0 | 0 | 1 | 3 | 5 | 2 | 0 | 0 | 11/12(91.7%) |
| Cefoperazone/sulbactam | 0 | 0 | 2 | 2 | 0 | 0 | 0 | 0 | 4/12(33.3%) |
| Dexamethasome | 0 | 2 | 1 | 2 | 6 | 0 | 0 | 0 | 11/12(91.7%) |
| B-IgY | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1/12(8.3%) |
| C-IgY | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1/12(8.3%) |
3.6. Effect of specific IgYs on cytokines in PDR-Ab-infected mice
The levels of pro-inflammatory cytokines TNF-α and IL-1β in serum of mice were determined by ELISA. The levels of TNF-α in all infected mice increased rapidly after PDR-Ab challenge, reached the highest values at 4 h and then decreased (Table 2 ). But the values of specific IgYs-treated mice were significantly lower than those of positive control and negative control mice (p < 0.05). After 6 h of treatment, TNF-α in specific IgYs-treated mice recoverd to normal level. The amounts of TNF-α in cefoperazone/sulbactam or dexamethason-treated mice were still in high level, but were much lower than that in negative control mice (p < 0.05). The levels of IL-1β changed in a similar trend (Table 3 ). These results indicated that specific IgYs could (p < 0.05) decrease the levels of pro-inflammatory cytokines in PDR-Ab -infected mice.
Table 2.
TNF-α levels in serum of infected mice after different treatment (pg/ml).
| Treatment | 0 h | 2 h | 4 h | 6 h |
|---|---|---|---|---|
| Blank control | 69.42 ± 0.49 | 71.53 ± 0.51 | 71.44 ± 0.62 | 70.85 ± 0.15 |
| Negative control | 68.96 ± 0.38 | 462.43 ± 0.80# | 673.41 ± 0.48# | 366.61 ± 0.55# |
| Cefoperazone/sulbactam | 69.72 ± 0.45 | 309.14 ± 0.27*,# | 462.53 ± 0.25*,# | 222.81 ± 0.73*,# |
| Dexamethasome | 69.11 ± 0.32 | 455.95 ± 0.38*,# | 655.64 ± 0.42*,# | 245.01 ± 0.17*,# |
| B-IgY | 70.44 ± 0.96 | 189.42 ± 0.42*,# | 196.47 ± 0.46*,# | 70.31 ± 0.28* |
| C-IgY | 71.48 ± 0.39 | 228.78 ± 0.75*,# | 248.16 ± 0.19*,# | 72.81 ± 0.57* |
Data are presented as means ± SD (n = 3).
p < 0.05 compared with the blank control at the same time.
p < 0.05 compared with the negative control at the same time.
Table 3.
IL-1β levels in serum of infected mice after different treatment (ng/ml).
| Treatment | 0 h | 2 h | 4 h | 6 h |
|---|---|---|---|---|
| Blank control | 284.71 ± 0.44 | 283.22 ± 0.88 | 284.58 ± 0.35 | 285.02 ± 0.78 |
| Negative control | 284.77 ± 0.22 | 298.45 ± 0.62# | 396.24 ± 0.19# | 334.73 ± 0.36# |
| Cefoperazone/sulbactam | 285.09 ± 0.34 | 297.84 ± 0.52# | 353.26 ± 0.75*,# | 296.89 ± 0.52*,# |
| Dexamethasome | 285.66 ± 0.32 | 318.05 ± 0.08*,# | 377.00 ± 0.23*,# | 289.62 ± 0.13* |
| B-IgY | 284.17 ± 0.40 | 300.50 ± 0.16*,# | 322.34 ± 0.50*,# | 285.04 ± 0.69* |
| C-IgY | 285.33 ± 0.22 | 284.81 ± 0.38* | 289.03 ± 0.34* | 284.88 ± 0.11* |
Data are presented as means ± SD (n = 3).
p < 0.05 compared with the blank control at the same time.
p < 0.05 compared with the negative control at the same time.
3.7. Protective effect of specific IgYs on lung tissue of PDR-Ab-infected mice
Protective effect of specific IgYs to PDR-Ab-infected mice was further assessed by histopathological examination on lung tissue sections (Fig. 6). H & E stained section of lung from negative control mice at 24 h showed serious infiltration of neutrophils with filling of the alveolar compared with the normal control lung tissue (Fig. 6A and B). These pathological features are consistent with acute pneumonia. There was a significant attenuation of the above pathological features in the lung tissue of mice treated by cefoperazone/sulbactam or dexamethason (Fig. 6C and D). Mild edema of alveolar was observed on the sections of lungs from specific IgYs-treated mice, but no infiltration of inflammatory cells was viewed (Fig. 6E and F). These results further demonstrated the efficacy of specific IgYs to PDR-Ab infection.
4. Discussion
A. baumannii is a kind of ubiquitous non-fermentative gram-negative bacillus that is commonly associated with aquatic environments [29]. Owing to the high ability of colonization in a warm and humid environment and natural genetic transformation, A. baumannii has currently exceeded other bacteria as the most commonly pathogen in clinical laboratories. Chittawatanarat et al. found that gram negative bacteria were the major pathogens of VAP, among which A. baumannii ranked first [30]. With the aggravation of antibacterial abuse in recent years, A. baumannii is now resistant to multiple antimicrobial agents including β-lactam antibacterial, aminoglycosides, tetracyclines, quinolones and so on. Most existing antibacterials can do nothing to the infection caused by PDR-Ab strains.
In previous study, we obtained seven strains of A. baumannii from respiratory department and ICU of the hospital, which were determined to be PDR-Ab by chemosensitivity test against twelve antimicrobial agents including minocycline, amikacin, ceftriaxone, ceftazidime, cefepime, cefoperazone, piperacillin, ticarcillin, meropenem, trimethoprim-sulfamethoxazole, levofloxacin and ciprofloxacin [22]. The tested strain Ab-C used in this study was one of the above PDR-Ab. The other A. baumannii strain Ab-B used here was also determined to be PDR-Ab by the same test. Generally, carbapenem antibacterial is the first choice for PDR-Ab infection [31]. After the first nosocomial carbapenem-resistant A. baumannii (CR-AB) strain was reported in 1991, CR-AB strains have been shown in increasing documents [32]. It was reported that 74% of A. baumannii were MDR-Ab and 55% of A. baumannii were resistant to imipenem [29]. Several mechanisms are responsible for the resistance of CR-AB, among which production of oxacillinases (OXA enzymes) is important one [29], [33]. We have documented that the PDR-Ab strains used in this study were resistant to meropenem, a new carbapenem antibacterial, and were positive for OXA-23, 51 and 58 enzymes [22]. So, the two MDR-Ab strains are suitable for estimating the activity of new antibacterial agents against A. baumannii.
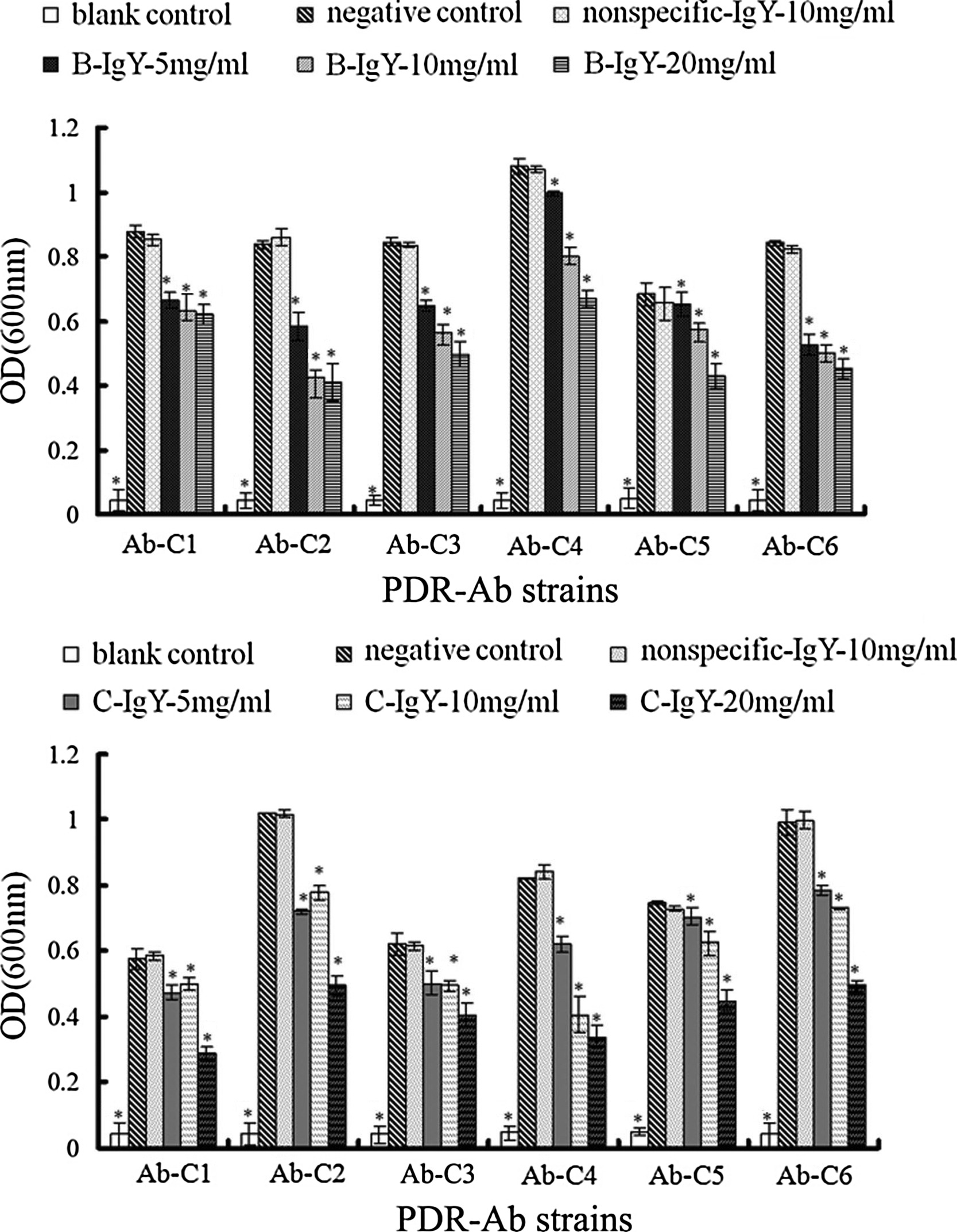
In this study, we use the two PDR-Ab strains obtained from different sources as antigens for producing specific IgYs. Results of bacteriostatic test demonstrated that the two specific IgYs did cross-react with the two PDR-Ab strains. Besides, we did antibacterial test on other 6 strains of MDR-Ab mentioned previously [22], in which the two specific IgYs also showed antibacterial activity (Supplementary Fig. S2). These suggested that IgY against one PDR-Ab strain might be applicable to other A. baumannii strains.
Growth inhibitory activity of the specific IgYs to clinically isolated strains. Six strains of clinically isolated PDR-Ab (106 cfu/ml) were respectively cultured with B-IgY or C-gY (5, 10 and 20 mg/ml), nonspecific IgY (10 mg/ml) in TSB liquid medium for 24 h. Data are presented as means ± SD (n = 3), *p < 0.05 compared with the negative control.
{kind=link}
Being gram-negative bacteria, A. baumannii cause disease by pathogenic factors such as capsule, pili, enzyme, lipid A and carrier. Various investigations have demonstrated that A. baumannii was able to adhere to and invade human epithelial cells and then induce epithelial cells death [34]. Lee et al. found that A. baumannii was capable to adhere to human bronchial epithelial cells [35]. Smani et al. confirmed that fibronectin was a potential host for A. baumannii, and the binding of A. baumannii to immobilized fibronectin was mediated by the outer membrane preparations [34]. In vitro study indicated that the specific IgYs inhibited the growth of PDR-Ab in a dose-dependent manner. The antibacterial activity of the two specific IgYs was comparable to that of cefoperazone/sulbactam. Both B-IgY and C-IgY at 20 mg/ml could significantly inhibit the growth of PDR-Ab within 24 h. On a whole, the inhibitory effect of IgYs on Ab-C was weaker than on Ab-B. Isolated from clinical sample, Ab-C is resistant to antibacterial on a wider and stronger degree. Therefore, we used the Ab-C as a model-building strain for murine acute pneumonia.
In our study, SEM assay indicated that specific IgYs enhanced the agglutination of MDR-Ab. It was reported that IgY could enhance the agglutination of bacteria causing a reduction in CFU rather than actual direct effects on individual bacteria [36]. SEM assay also showed cell crenation and structural modification on the surface of specific IgYs-treated bacteria. These resulted from the combination of IgY with bacteria, and might reduce the attachment of bacteria on mucosa. The similar function of the specific IgY was ever reported about Helicobacter pylori attaching to gastric cancer cells [10], and Salmonella attaching to intestinal cells [37], [38].
In addition to agglutination and adherence-blockade, phagocytosis opsonization is another important mechanism of IgY mediated antibacterial activity [39]. After infection, the quantity of macrophages in local infected tissue increased by recruitment of circulating monocytes and/or in situ proliferation. The activated macrophages with pro-inflammatory phenotype produce various pro-inflammatory mediators, such as TNF-α and IL-1β, which play a central role in the inflammatory process [40], [41], [42], [43]. IgY binding may alter the electron cloud and/or electric field on the bacterial surface [44], and consequently increase vulnerability of the bacteria cells to phagocytosis. Our previous study demonstrated that specific IgY increased the phagocytosis of S. aureus by milk macrophages [45]. Nie et al. revealed that IgY improved the phagocytosis of S. aureus by neutrophils [46]. In this study, though the increase of pro-inflammatory cytokines was observed in all model mice, the levels of TNF-α and IL-1β in specific IgYs-treated mice were much lower and recovered to normal more rapidly than those in other groups. We inferred that IgY might improve the phagocytosis of macrophages to PDR-Ab, as a result, the quantity of macrophage recruited and activated in infected tissue decreased and the pro-inflammatory cytokines produced by macrophages reduced.
We have previously proved the protective effects of specific IgY as a substitution of antibacterial in rats with prevotella intermedia-mediated gingivitis [26] and in mice with lipopolysaccharide-induced endotoxemia [28]. In this study, mice model of Ab-induced acute pneumonia was builded by nasal inhalation of PDR-Ab and used to estimate the efficacy of specific IgYs against A. baumannii. Pneumonia is a common disease of respiratory system. VAP caused by multi-drug resistant Gram-negative pathogens, especially PDR-Ab, threats patients in ICU critically. With few novel antibacterial in late-stage clinical development, clinicians may soon be left with little choice for the treatment of PDR-Ab infections. The present study indicated that IgYs against PDR-Ab could prevent pneumonic mice from death and attenuate inflammation in the infected lungs.
Cefoperazone/sulbactam and dexamethasone were used as positive controls to compare with IgYs in this study. Cefoperazone/sulbactam, a compound preparation of cephalosporin and β-lactamase inhibitor, showed stronger antibacterial activity than specific IgYs in vitro. But its death prevention and anti-inflammatory effects in vivo were much lower than those of specific IgYs. Dexamethasone, an anti-inflammatory glucocorticoid, showed no death prevention effect and a little anti-inflammatory effect in mice with acute pneumonia. But the down-regulation effects of dexamethasone to TNF-α and IL-1β were not comparable with those of specific IgYs. Based on above, specific IgYs are more effective than cefoperazone/sulbactam and dexamethasone against PDR-Ab in vitro and in vivo. Considering its other advantages together, specific IgY may be a promising substitution of the current antibacterials.
In conclusion, specific IgYs produced by immunizing laying hens with PDR-Ab were able to inhibit the growth of A. baumannii in a dose-dependent manner and protect mice from acute pneumonia induced by A. baumannii. The specific IgYs have the potential to be used as a new therapeutic approach for the treatment of PDR-Ab infections.
Conflict of interest
The authors declare that they have no conflict of interest.
Acknowledgement
This work was financially supported by Medical Research Fund of Dalian Municipal Commission of Health and Family Planning, China (2013).
References
- 1.Li S., Li H.T., Qi T.J., Yan X.X., Wang B.L., Guan J.T., Li Y. Comparative transcriptomics analyses of the different growth states of multidrug-resistant Acinetobacter baumannii. Biomed. Pharmacother. 2017;85:564–574. doi: 10.1016/j.biopha.2016.11.065. [DOI] [PubMed] [Google Scholar]
- 2.Endimiani A., Hujer K.M., Hujer A.M., Bertschy I., Rossano A., Koch C., Gerber V., Francey T., Bonomo R.A., Perreten V. Acinetobacter baumannii isolates from pets and horses in Switzerland: molecular characterization and clinical data. J. Antimicrob. Chemother. 2011;66:2248–2254. doi: 10.1093/jac/dkr289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Luo G., Spellberg B., Gebremariam T., Bolaris M., Lee H., Fu Y., French S.W., Ibrahim A.S. Diabetic murine models for Acinetobacter baumannii infection. J. Antimicrob. Chemother. 2012;67:1439–1445. doi: 10.1093/jac/dks050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Shin J.A., Chang Y.S., Kim H.J., Kim S.K., Chang J., Ahn C.M., Byun M.K. Clinical outcomes of tigecycline in the treatment of multidrug-resistant Acinetobacter baumannii infection. Yonsei Med. J. 2012;53:974–984. doi: 10.3349/ymj.2012.53.5.974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Munoz-Price L.S., Weinstein R.A. Acinetobacter infection. N. Engl. J. Med. 2008;358:1271–1281. doi: 10.1056/NEJMra070741. [DOI] [PubMed] [Google Scholar]
- 6.Sopirala M.M., Mangino J.E., Gebreyes W.A., Biller B., Bannerman T., Balada-Llasat J.M., Pancholi P. Synergy testing by Etest microdilution checkerboard, and time-kill methods for pan-drug-resistant Acinetobacter baumannii. Antimicrob. Agents Chemother. 2010;54:4678–4683. doi: 10.1128/AAC.00497-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Smani Y., Pachón-Ibáñez M.E., Pachón J. New molecules and adjuvants in the treatment of infections by Acinetobacter baumannii. Expert. Opin. Pharmacother. 2016;17:1207–1214. doi: 10.1080/14656566.2016.1176144. [DOI] [PubMed] [Google Scholar]
- 8.Amaral J.A., Tino De Franco M., Carneiro-Sampaio M.M., Carbonare S.B. Anti-enteropathogenic Escherichia coli immunoglobulin Y isolated from eggs laid by immunized Leghorn chickens. Res. Vet. Sci. 2002;72:229–234. doi: 10.1053/rvsc.2002.0551. [DOI] [PubMed] [Google Scholar]
- 9.Zhen Y.H., Jin L.J., Li X.Y., Guo J., Li Z., Zhang B.J., Fang R., Xu Y.P. Efficacy of specific egg yolk immunoglobulin (IgY) to bovine mastitis cause by Staphylococus aureus. Vet. Microbiol. 2009;133:317–322. doi: 10.1016/j.vetmic.2008.07.016. [DOI] [PubMed] [Google Scholar]
- 10.Shin J.H., Yang M., Nam S.W., Kim J.T., Myung N.H., Bang W.G., Roe I.H. Use of egg yolk-derived immunoglobulin as an alternative to antibiotic treatment for control of Helicobacter pylori infection. Clin. Diagn. Lab. Immunol. 2002;9:1061–1066. doi: 10.1128/CDLI.9.5.1061-1066.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Yokoyama H., Umeda K., Peralta R.C., Hashi T., Icatlo F.C.J., Kuroki M., Ikemori Y., Kodama Y. Oral passive immunization against experimental salmonellosis in mice using chicken egg yolk antibodies specific for Salmonella enteritidis and S. typhimurium. Vaccine. 1998;16:388–393. doi: 10.1016/s0264-410x(97)80916-4. [DOI] [PubMed] [Google Scholar]
- 12.LeClaire R.D., Hunt R.E., Bavari S. Protection against bacterial superantigen staphylococcal enterotoxin B by passive vaccination. Infect. Immun. 2002;70:2278–2281. doi: 10.1128/IAI.70.5.2278-2281.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Zhen Y.H., Jin L.J., Guo J., Li X.Y., Lu Y.N., Chen J., Xu Y.P. Charaterization of specific egg yolk immunoglobulin (IgY) against mastitis-casusing Escherichia coli. Vet. Microbiol. 2008;130:126–133. doi: 10.1016/j.vetmic.2007.12.014. [DOI] [PubMed] [Google Scholar]
- 14.Hatta H., Tsuda K., Ozeki M., Kim M., Yamamoto T., Otake S., Hirasawa M., Katz J., Childers N.K., Michalek S.M. Passive immunization against dental plaque formation in humans: effect of a mouth rinse containing egg yolk antibodies (IgY) specific to Streptococcus mutans. Caries Res. 1997;31:268–274. doi: 10.1159/000262410. [DOI] [PubMed] [Google Scholar]
- 15.Lee S.B., Mine Y., Stevenson R.M.W. Effects of hen egg yolk immunoglobulin in passive protection of rainbow trout against Yersinia ruckeri. J. Agric. Food Chem. 2000;48:110–115. doi: 10.1021/jf9906073. [DOI] [PubMed] [Google Scholar]
- 16.Hatta H., Tsuda K., Akachi S., Kim M., Yamamoto T., Ebina T. Oral passive immunization effect of anti-human rotavirus IgY and its behavior against proteolytic enzymes. Biosci. Biotechnol. Biochem. 1993;57:1077–1081. doi: 10.1271/bbb.57.1077. [DOI] [PubMed] [Google Scholar]
- 17.Ikemori Y., Ohta M., Umeda K., Icatlo F.C.J., Kuroki M., Yokoyama H., Kodama Y. Passive protection of neonatal calves against bovine coronavirus-induced diarrhea by administration of egg yolk or colostrum antibody powder. Vet. Microbiol. 1997;58:105–111. doi: 10.1016/S0378-1135(97)00144-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Liou J.F., Chang C.W., Tailiu J.J., Yu C.K., Lei H.Y., Chen L.R., Tai C. Passive protection efftect of chicken egg yolk immunoglobulins on enterovirus 71 infected mice. Vaccine. 2010;28:8189–8196. doi: 10.1016/j.vaccine.2010.09.089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.M.A. Coleman, Oral administration of chicken yolk immunoglobulins to lower somatic cell count in the milk of lactating ruminants, US Patent (1996) 5585098.
- 20.Li Y., Wang L.L., Zhen Y.H., Li S.Y., Xu Y.P. Chicken egg yolk antibodies (IgY) as non-antibiotic production enhancers for use in swine production: a review. J. Anim. Sci. Biotechnol. 2015;6:40. doi: 10.1186/s40104-015-0038-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Carlander D., Kollberg H., Wejaker P.E., Larsson A. Peroral immunotherapy with yolk antibodies for the prevention and treatment of enteric infections. Immunol. Res. 2000;21:1–6. doi: 10.1385/IR:21:1:1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Sun L.D., Han Z.M., Qu J., Luo X., Fan T.F., Chen X.L., Guo X., Xu H., Zhen Y.H. The drug resistance of Acinetobacter baumannii and the related resistant genes of pan-drug-resistant strains. Chin. J. Microe. 2015;27:688–6891. [Google Scholar]
- 23.Zhang M., Song Y., Ci X.X., An N., Fan J.W., Cui J.Q., Deng X. Effects of florfenicol on early cytokine responses and survival in murine endotoxemia. Int. Immunopharmacol. 2008;8:982–988. doi: 10.1016/j.intimp.2008.02.015. [DOI] [PubMed] [Google Scholar]
- 24.Sun Y., Yang Y.H., Wang L.F., Lv L., Zhu J., Han W.Q., Wang E.X., Guo X., Zhen Y.H. Highly sensitive detection of cancer antigen human epidermal growth factor receptor 2 using novel chicken egg yolk immunoglobulin. Biologicals. 2015;43:165–170. doi: 10.1016/j.biologicals.2015.03.002. [DOI] [PubMed] [Google Scholar]
- 25.Akita E.M., Nakai S. Immunoglobulins from egg yolk: isolation and purification. J. Food Sci. 1992;57:629–634. [Google Scholar]
- 26.Hou Y.Y., Zhen Y.H., Wang D., Zhu J., Sun D.X., Liu X.T., Wang H.X., Liu Y., Long Y.Y., Shu X.H. Protective effect of an egg yolk-derived immunoglobulin (IgY) against prevotella intermedia mediated gingivitis. J. Appl. Microbiol. 2014;116:1020–1027. doi: 10.1111/jam.12419. [DOI] [PubMed] [Google Scholar]
- 27.Lobbedey L., Schlatterer B. Development and application of an ELISA for the detection of duck antibodies against Riemerella anatipestifer antigens in egg yolk of vaccines and in serum of their offspring. J. Vet. Med. B Infect. Dis. Vet. Public Health. 2003;50:81–85. doi: 10.1046/j.1439-0450.2003.00624.x. [DOI] [PubMed] [Google Scholar]
- 28.Zhen Y.H., Fang R., Ding C., Jin L.J., Li X.Y., Diao Y.P., Shu X.H., Ma X.C., Xu Y.P. Efficacy of specific IgY for treatment of lipopolysaccharide-induced endotoxemia using a mouse model. J. Appl. Microbiol. 2011;111:1524–1532. doi: 10.1111/j.1365-2672.2011.05155.x. [DOI] [PubMed] [Google Scholar]
- 29.Pourhajibagher M., Hashemi F.B., Pourakbari B., Aziemzadeh M., Bahador A. Antimicrobial resistance of Acinetobacter baumannii to imipenem in Iran: a systematic review and meta-analysis. Open Microbiol. J. 2016;10:32–42. doi: 10.2174/1874285801610010032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Chittawatanarat K., Jaipakdee W., Chotirosniramit N., Chandacham K., Jirapongcharoenlap T. Microbiology, resistance patterns, and risk factors of mortality in ventilator-associated bacterial pneumonia in a Northern Thai tertiary-care university based general surgical intensive care unit. Infect. Drug Resist. 2014;7:203–210. doi: 10.2147/IDR.S67267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Tang S.S., Apisarnthanarak A., Hsu L.Y. Mechanisms of β-lactam antimicrobial resistance and epidemiology of major community and healthcare-associated multidrug-resistant bacteria. Adv. Drug Deliv. Rev. 2014;78:3–13. doi: 10.1016/j.addr.2014.08.003. [DOI] [PubMed] [Google Scholar]
- 32.Huang S.T., Chiang M.C., Kuo S.C., Lee Y.T., Chiang T.H., Yang S.P., Yin T., Chen T.L., Fung C.P. Risk factors and clinical outcomes of patients with carbapenem-resistant Acinetobacter baumannii bacteremia. J. Microbiol. Immunol. Infect. 2012;45:356–362. doi: 10.1016/j.jmii.2011.12.009. [DOI] [PubMed] [Google Scholar]
- 33.Olaitan A.O., Berrazeg M., Fagade O.E., Adelowo O.O., Alli J.A., Rolain J.M. Emergence of multidrug-resistant Acinetobacter baumannii producing OXA-23 carbapenemase, Nigeria. Int. J. Infect. Dis. 2013;17:e469–470. doi: 10.1016/j.ijid.2012.12.008. [DOI] [PubMed] [Google Scholar]
- 34.Smani Y., McConnell M.J., Pachón J. Role of fibronectin in the adhesion of Acinetobacter baumannii to host cells. PLoS One. 2012;7:1–6. doi: 10.1371/journal.pone.0033073. e33073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Lee J.C., Koerten H., van den Broek P., Beekhuizen H., Wolterbeek R., van den Barselaar M., van der Reijden T., van der Meer J., van de Gevel J., Dijkshoorn L. Adherence of Acinetobacter baumannii strains to human bronchial epithelial cells. Res. Microbiol. 2006;157:360–366. doi: 10.1016/j.resmic.2005.09.011. [DOI] [PubMed] [Google Scholar]
- 36.Tsubokura K., Berndtson E., Bogstedt A., Kaijse B., Kim M., Ozeki M., Hammarström L. Oral administration of antibodies as prophylaxis and therapy in Campylabacter jejuni-infected chickens. Clin. Exp. Immunol. 1997;108:451–455. doi: 10.1046/j.1365-2249.1997.3901288.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Sugita-Konishi Y., Shibata K., Yun S.S., Hara-Kudo Y., Yamaguchi K., Kumagai S. Immune functions of immunoglobulin Y isolated from egg yolk of hens immunized with various infectious bacteria. Biosci. Biotechnol. Biochem. 1996;60:886–888. doi: 10.1271/bbb.60.886. [DOI] [PubMed] [Google Scholar]
- 38.Sugita-Konishi Y., Ogawa M., Arai S., Kumagai S., Igimi S., Shimizu M. Blockade of Salmonella enteritidis passage across the basolateral barriers of human intestinal epithelial cells by specific antibody. Microbiol. Immunol. 2000;44:473–479. doi: 10.1111/j.1348-0421.2000.tb02522.x. [DOI] [PubMed] [Google Scholar]
- 39.Xu Y.P., Li X.Y., Jin L.J., Zhen Y.H., Lu Y.N., Li S.Y., You J.S., Wang L.H. Application of chicken egg yolk immunoglobulins in the control of terrestrial and aquatic animal diseases: a review. Biotechnol. Adv. 2011;29:860–868. doi: 10.1016/j.biotechadv.2011.07.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Zhang L., Wang C.C. Inflammatory response of macrophages in infection. HBPD Int. 2014;13:138–152. doi: 10.1016/s1499-3872(14)60024-2. [DOI] [PubMed] [Google Scholar]
- 41.Mills K.H., Dunne A. Immune modulation: IL-1, master mediator or initiator of inflammation. Nat. Med. 2009;15:1363–1364. doi: 10.1038/nm1209-1363. [DOI] [PubMed] [Google Scholar]
- 42.Sedger L.M., McDermott M.F. TNF and TNF-receptors: from mediators of cell death and inflammation to therapeutic giants-past, present and future. Cytokine Growth Factor Rev. 2014;25:453–472. doi: 10.1016/j.cytogfr.2014.07.016. [DOI] [PubMed] [Google Scholar]
- 43.Striz I., Brabcova E., Kolesar L., Sekerkova A. Cytokine networking of innate immunity cells: a potential target of therapy. Clin. Sci. 2014;126:593–612. doi: 10.1042/CS20130497. [DOI] [PubMed] [Google Scholar]
- 44.Lee E.N., Sunwoo H.H., Menninen K., Sim J.S. In vitro studies of chicken egg yolk antibody (IgY) against Salmonella enteritidis and Salmonella typhimurium. Poultry Sci. 2002;81:632–641. doi: 10.1093/ps/81.5.632. [DOI] [PubMed] [Google Scholar]
- 45.Zhen Y.H., Jin L.J., Guo J., Li X.Y., Li Z., Fang R., Xu Y.P. Characterization of specific egg yolk immunoglobulin (IgY) against mastitis-causing Staphylococcus aureus. J. Appl. Microbiol. 2008;105:1529–1535. doi: 10.1111/j.1365-2672.2008.03920.x. [DOI] [PubMed] [Google Scholar]
- 46.Nie R., Wu D., Hu G., Zhang J., Yang H., Wen Z. Effect of specific egg yolk immunoglobulins on phagocytosis by neutrophils. Chin. J. Vet Med. 2004;12:23–25. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Liver histopathology of A. baumannii-infected mice received different treatments. A. blank group: liver tissue from healthy mice; B. negative group: liver tissue from infected mice which were treated with 0.9% NaCl;C. cefoperazone/sulbactam group: liver tissue from infected mice which were treated with cefoperazone /sulbactam; D. dexamethasone group: liver tissue from infected mice which were treated with dexamethasone;E. B-IgY group: liver tissue from infected mice which were treated with B-IgY; F. C-IgY group: liver tissue from infected mice which were treated with B-IgY.
Growth inhibitory activity of the specific IgYs to clinically isolated strains. Six strains of clinically isolated PDR-Ab (106 cfu/ml) were respectively cultured with B-IgY or C-gY (5, 10 and 20 mg/ml), nonspecific IgY (10 mg/ml) in TSB liquid medium for 24 h. Data are presented as means ± SD (n = 3), *p < 0.05 compared with the negative control.