

Abstract
Manuka honey, a wound treatment used to eradicate bacteria, resolve inflammation, and promote wound healing, is a current focus in the tissue engineering community as a tissue template additive. However, Manuka honey’s effect on neutrophils during the inflammation-resolving phase has yet to be examined. This study investigates the effect of 0.5% and 3% Manuka honey on the release of cytokines, chemokines, and matrix-degrading enzymes from a dHL-60 neutrophil model in the presence of anti-inflammatory stimuli (TGF-β, IL-4, IL-4 +IL-13). We hypothesized that Manuka honey would reduce the output of pro-inflammatory signals and increase the release of anti-inflammatory signals. The results of this study indicate that 0.5% honey significantly increases the release of CXCL8/IL-8, CCL2/MCP-1, CCL4/MIP-1β, CCL20/MIP-3α, IL-4, IL-1ra, and FGF-13 while reducing Proteinase 3 release in the anti-inflammatory-stimulated models. However, 3% honey significantly increased the release of TNF-α and CXCL8/IL-8 while reducing the release of all other analytes. We replicated a subset of the most notable findings in primary human neutrophils, and the consistent results indicate that the HL-60 data are relevant to the performance of primary cells. These findings demonstrate the variable effects of Manuka honey on the release of cytokines, chemokines, and matrix-degrading enzymes of this model of neutrophil anti-inflammatory activity. This study reinforces the importance of tailoring the concentration of Manuka honey in a wound or tissue template to elicit the desired effects during the inflammation-resolving phase of wound healing. Future in vivo investigation should be undertaken to translate these results to a physiologically-relevant wound environment.
Keywords: Tissue Engineering, Cytokine, Inflammation
Graphical Abstract
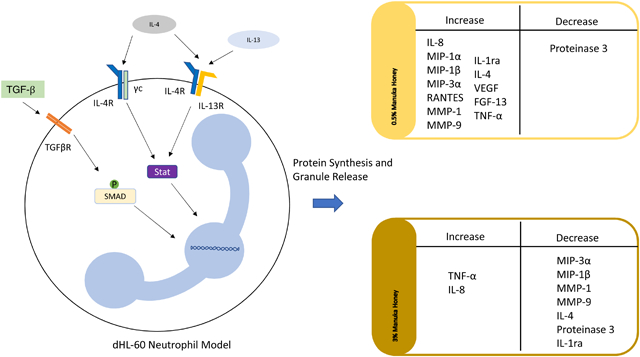
Summary Sentence:
Manuka honey modulates the cytokine release of a dHL-60 neutrophil model and primary human neutrophils under anti-inflammatory stimulation conditions.
1. Introduction
Manuka honey, a variety of honey indigenous to New Zealand, has been widely researched as a wound treatment due to its extraordinary wound-healing properties [1–4]. These properties include reducing bacterial infection, an effect attributed to its methylglyoxal and hydrogen peroxide content, as well as accelerating wound closure [1, 4–9]. Manuka honey’s high sugar concentration creates an osmotic effect within the wound, causing a flow of liquid and nutrients from the deeper subcutaneous tissue into the wound and flushing debris and bacteria out of the wound bed [1, 10]. This high sugar concentration also gives the honey a viscous quality, creating a protective covering over the wound which hydrates the area and prevents additional bacterial deposition [1]. The sugars also provide a nutrient source for proliferating cells such as endothelial cells and fibroblasts [1]. Manuka honey’s acidic pH increases wound oxygenation as well as fibroblast and macrophage activity [10, 11]. The flavonoids within the honey react with free radicals, reducing tissue damage within the wound [10, 11]. These flavonoids have been quantified via HPLC-MS analysis, and are also theorized to improve the antioxidant response of cells within the wound [4]. Major relevant anti-inflammatory constituents include the antioxidant flavonoids pinobanksin, pinocembrin, and chrysin, as well as the myeloperoxidase-inhibiting compounds methyl syringate and leptosin [12]. These components are thought to cross the cell membrane and scavenge free radicals within the cytosol, reducing the activation of inflammatory pathways and activating antioxidant pathways within the cell [4]. Anti-inflammatory properties of Manuka honey have been corroborated in several animal models and clinical trials [6, 13–17]. Other honey varieties, such as buckwheat honey, tualang honey, Ulmo honey, and Capparis spinosa honey have been shown to have similar antioxidant and antibacterial properties beneficial in wound healing [18–22]. However, Manuka honey’s unique methylglyoxal content is a potent tool against antibiotic-resistant bacteria [22–24]. We have chosen to focus this research on Manuka honey because it is the most widely-used commercially available in wound healing products [25, 26]. A few analyses, however, have demonstrated that high concentrations of Manuka honey can have a counterproductive cytotoxic effect [27–29]. We have previously reported a cytotoxic limit of 5% v/v Manuka honey, and as such, use concentrations below this limit in this current work [30]. Our previous work indicated that 0.5% and 3% honey were effective at reducing activation of the NF-κB pathway via IκBα phosphorylation in a dose-dependent manner. Therefore, we have chosen to focus on these honey concentrations in this research. Timepoints of 3 hours and 24 hours were chosen to focus on the events within the first day after application of Manuka honey to a wound, as this is the timeframe when neutrophil behavior would be most relevant.
Within a period of minutes to hours after the occurrence of injury, neutrophils swarm the wound site and begin orchestrating the inflammatory response within the wound [31, 32]. These cells kill bacteria through phagocytosis and reactive oxygen species release and produce a variety of cytokines, chemokines, and matrix-degrading enzymes. These mediators recruit more neutrophils and other immune cells, activate inflammatory pathways within those newly recruited cells, and break down the ECM within the wound site to clear debris and free vascular endothelial cells for angiogenesis [33–35]. This initial acute inflammatory phase is necessary for the destruction of bacteria and removal of dead cells and damaged ECM from the wound. However, if this inflammation persists and becomes chronic, it can prevent the wound from healing [36–38]. In addition to their pro-inflammatory role, however, neutrophils also have the capability to release anti-inflammatory mediators, decreasing inflammatory behaviors in the surrounding immune cells, reducing further immune cell recruitment, and promoting wound healing [39–41]. This anti-inflammatory activity contributes to the resolution of acute inflammation within the wound, preventing the wound from being stuck in a phase of chronic inflammation and allowing it to heal.
In this study, the effect of Manuka honey on neutrophil anti-inflammatory activity is investigated using a dHL-60 model of a neutrophil. The HL-60 cell line is a promyelocytic progenitor that can be differentiated to a neutrophil-like phenotype using DMSO [42–45]. The use of this model eliminates the donor-to-donor variability found in the primary neutrophil cytokine response, and it has been previously validated as a model of neutrophil cytokine release [46, 47]. We have investigated the effects of Manuka honey on this model, and reported reduced superoxide release, chemotaxis, and activation of the inflammatory transcription factor NF-κB in the presence of Manuka honey concentrations below the cytotoxic limit of 5% v/v [30]. In addition, we have investigated Manuka honey’s effect on the dHL-60 cytokine release profile in the presence of several inflammatory stimulators, and found a dose-dependent effect of reducing several inflammatory mediators and ECM-degrading components [48].
This study investigates the dHL-60 release of cytokine, chemokine, and matrix-degrading enzymes under anti-inflammatory stimuli in the presence of Manuka honey. Three anti-inflammatory stimuli were used: TGF-β, IL-4, and the combination of IL-4 and IL-13. TGF-β activates the SMAD/RSMAD pathway, while IL-4 activates the Akt/PKB pathway, and, in concert with IL-13, the Jak/STAT pathway [49–51]. These signals have been demonstrated to reduce neutrophil superoxide production and activate neutrophil release of the anti-inflammatory factor IL-1ra [52, 53]. Due to the hurdles of working with primary human neutrophils in culture, such as their patient-to-patient variability and limited lifespan outside the body, it was decided to do the bulk of the experimentation using the dHL-60 neutrophil model system, and use primary human neutrophils to confirm several of the most interesting findings from that system. The dHL-60s were preconditioned with these stimulators (TGF-β, IL-4, and IL-4 + IL-13) for 72 hours prior to the addition of Manuka honey to allow for the activation of their respective pathways, and then cultured with both their respective stimulators in the absence of honey or the presence of 0.5% or 3% v/v Manuka honey for 3 or 24 hours. The supernatant of these cultures was then assayed for several pro-inflammatory signals, anti-inflammatory signals, matrix degrading enzymes, angiogenic growth factors, and chemoattractants. To validate these findings, primary human neutrophils were cultured with 0, 10, 100, or 1000 ng/mL IL-4 in the absence of honey or presence of 0.5% or 3% v/v Manuka honey for 3 or 24 hours. Supernatant collected from these samples was assayed for MMP-9, VEGF, and IL-1ra, and these results were compared to the cytokine production of the dHL-60 neutrophil model in the absence of honey or presence of 0.5% or 3% v/v Manuka honey. These experiments reveal the impact of Manuka honey on neutrophil cytokine output under anti-inflammatory stimulation, informing future honey-based therapies and biomaterials.
2. Materials and Methods
2.1. HL-60 culture and differentiation
The HL-60 cells were obtained from ATCC (CCL240) (Manasses, VA, USA) and cultured in RPMI 1640 with 10% v/v non-heat-inactivated FBS, 1% v/v penicillin/streptomycin, and 1% L-glutamine (hereafter referred to as culture media), all obtained from Hyclone (Logan, UT, USA). Cells were cultured at a density ranging from 2 × 105 to 5 × 105 cells per mL. These cells were maintained at 37 °C in a 5% CO2 incubator. Throughout the culture period, medium was changed every 3–4 days and cells were passaged at a cell density of 5 × 105 cells/mL. Cells were discarded after reaching a passage number of 30. HL-60s were differentiated to a neutrophil-like phenotype using a previously validated procedure, adding 1.25% DMSO (Sigma Aldrich, St. Louis, MO, USA) to the culture medium for 6 days and replenishing medium/DMSO on the third day [42, 54]. The morphology of these differentiated cells was observed by permeabilizing the cells with 0.17 mM Triton X-100 (Fisher Scientific, Hampton, NH, USA) for 5 minutes, then fixing in 10% buffered formalin (Fisher Scientific, Hampton, NH, USA) and staining with DAPI (NucBlue Fixed Cell Stain ReadyProbes reagent) for 5 minutes at stock concentration and phalloidin-conjugated Alexa Fluor 488 (ActinGreen 488 Ready Probes reagent) (both from Invitrogen, Carlsbad, CA, USA) for 30 minutes also at stock concentration according to the manufacturer’s protocols. Imaging was conducted using an Olympus microscope (model BX34F) with attached Olympus DP73 digital color camera and Olympus U-HGLGPS fluorescent light source (Olympus, Shinjuku, Tokyo, Japan). The percentage of differentiated cells (kidney-shaped nucleus) was calculated to be 69%, comparable to the percentage range of 60–90% reported in literature [54–56].
2.2. Cytokine release experiment
First, dHL-60s were pre-conditioned for 72 hours with their respective stimuli. Cells were maintained in T-75 culture flasks (Thermo Scientific, Rochester, NY, USA) and either not stimulated (control) or stimulated with 2 ng/mL TGF-β, 10 ng/mL IL-4, or both 10 ng/mL IL-4 and 10 ng/mL IL-13. Concentrations of these analytes were taken from prior published studies [30, 52]. After the 72-hour pre-conditioning period, the dHL-60s were seeded in a 96-well culture plate at 400,000 cells per well (2.67 million cells per mL) in 150 μL of culture media with their respective stimuli and in the presence of 0%, 0.5%, and 3% Manuka honey (UMF 12+, Manuka Guard, Monterey, CA, USA) alongside honey/medium blanks which contained no cells. Cells were cultured for 3 or 24 hours and then centrifuged, and supernatants were removed and saved. These supernatants were analyzed using a multiplexed magnetic bead immunoassay (R&D Systems, Minneapolis, MN, USA) on a MAGPIX® reader (Luminex, Austin, TX, USA). Additionally, a control group cultured for 24 hours without Manuka honey was run again and its values were compared against the values from the first kit run to determine reproducibility.
2.3. Primary human neutrophil experiments
Neutrophils were isolated from healthy donor blood obtained from Tennessee Blood Services as specified by protocols approved by the University of Tennessee Institutional Review Board as previously described [57]. After the neutrophils were isolated, they were resuspended in RPMI 1640 with 1% L-glutamine, 5% penicillin/streptomycin, and 5% human serum isolated from the same donor blood as the neutrophils (hereafter referred to as RPMI+) [58]. Neutrophils were seeded into a 96-well culture plate at 400,000 cells per well (2.67 million cells per mL) in 150 μL of RPMI+ with the respective concentration of IL-4 and Manuka honey and cultured for 3 or 24 hours. Immediately following the culture period, plates were placed on ice for 10 minutes to inhibit the further stimulation of the neutrophils. Samples were pipetted into microcentrifuge tubes and centrifuged at 300 × g for 5 minutes to separate out the cells, and the supernatants were frozen and saved for analysis. 20 μL of trypsin (Gibco, Dublin, Ireland) was added to each well, and the culture plates were placed back in the incubator for 5 minutes to lift any remaining adherent cells. After the incubation period, 20 μL of RPMI+ was added to each well to inactivate the trypsin, and then the trypsin/media was removed from each well and added to labeled microcentrifuge tubes, which were then centrifuged at 300 × g for 5 minutes. Supernatants were discarded, and cells were resuspended in 75 μL RPMI+ with 75 μL trypan blue (Gibco). The numbers of viable and non-viable cells in each sample were counted with a Countess II FL Automated Cell Counter (Invitrogen) [30]. Two unique experiments were performed with cells isolated from two unique same-gender (male) and age-matched (between 20 and 40 years of age) donors, and the results were pooled.
2.4. Statistical Analysis
Groups were compared using non-parametric methods due to the data distributions. The results of the control group between the two assay runs were compared using the Wilcoxon rank sum test for each analyte. To be conservative in our identification of statistically significant differences, values below the Min LOD were treated as though they were equal to the Min LOD during the statistical analysis. Using an α=0.05, there were no significant differences found between the dHL-60 assay runs, which indicates reproducibility and allows for comparisons to be made between the assay runs. Stimulation groups at each honey level (0%, 0.5%, and 3%) were compared using the Kruskal Wallis test with a Bonferroni p-value adjustment to account for multiple comparisons (α=0.05). To control the family-wise error rate (FWER), the current anti-inflammatory data were analyzed a priori alongside pro-inflammatory stimulation groups, results of which have been reported in a previous publication. 50
3. Results
3.1. dHL-60 experiments
The release results of each analyte at each level of honey after 3 hours of culture are displayed in Figure 1, grouped into boxes by trend. In general, 3% honey decreased the levels of all analytes except TNF-α and IL-8, while 0.5% honey increased the release of both pro-inflammatory and anti-inflammatory analytes compared to the non-honey controls. Box A contains the results for TNF-α, the only analyte which had the highest release in the presence of 3% honey in all groups except the TGF-β-stimulated group. The presence of 0.5% honey also increased TNF-α release relative to the respective non-honey groups in the cells stimulated with TGF-β, IL-4, and IL-4 + IL-13. These data indicate that TGF-β has a statistically-significant suppressive effect on TNF-α release at 3% honey, but at 0.5% honey it significantly increases TNF-α release relative to the non-stimulated control. In contrast, stimulation by IL-4 and IL-4 + Il-13 increased TNF-α release at 0.5% honey relative to the non-stimulated cells (statistically significant in the IL-4 + IL-13 group). However, at 3% honey there was only a non-significant trend of increased TNF-α in the IL-4 + Il-13 group relative to the non-control cells, with no difference in the IL-4 group from the control.
Figure 1.
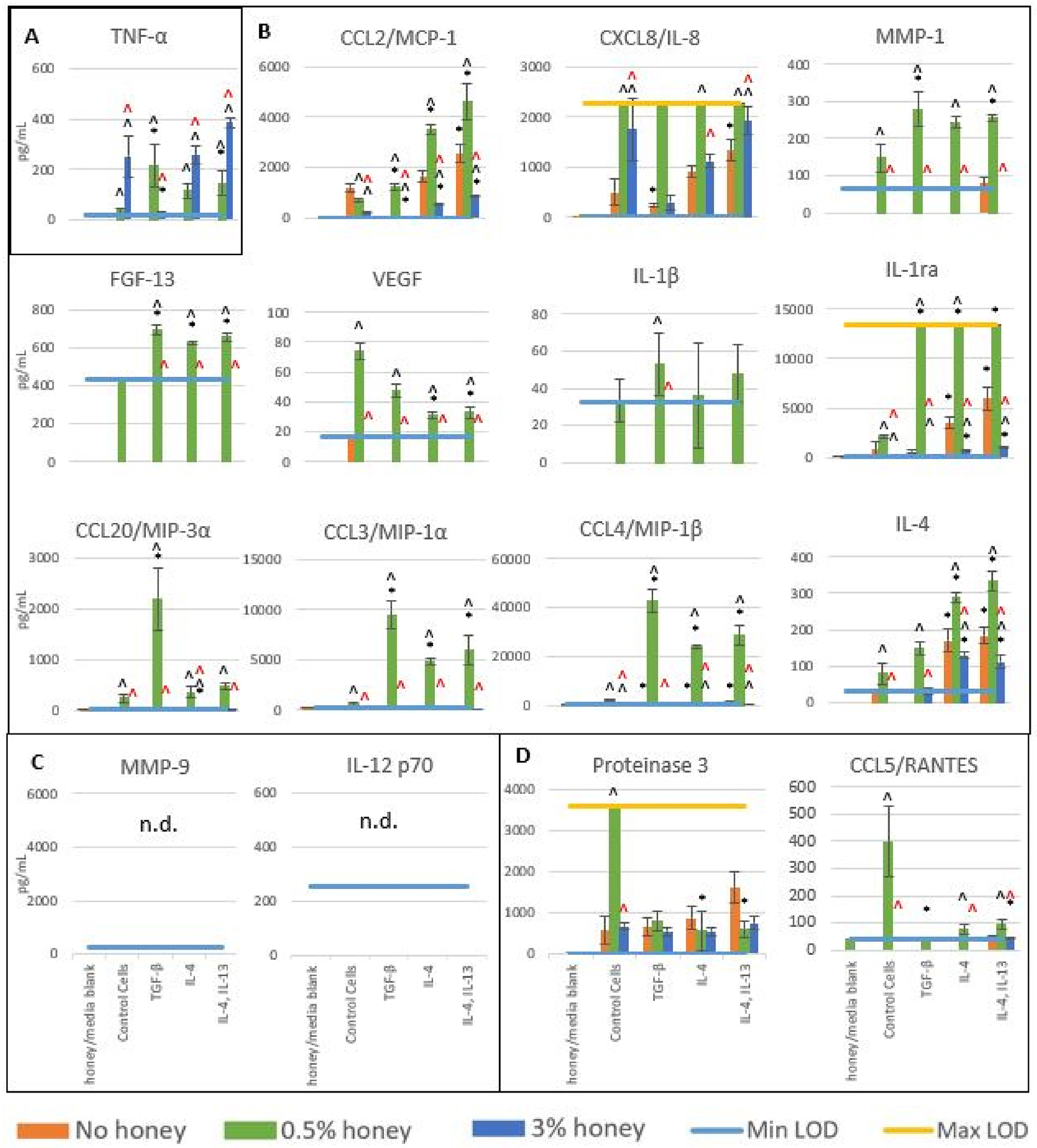
Release levels of cytokines, chemokines, and matrix-degrading enzymes in the absence of honey or presence of 0.5% or 3% honey at 3 hours. Bars indicate mean ± standard deviation. * indicates a statistically significant difference from the non-stimulated control at the respective honey level, ^ indicates a statistically significant difference from the non-honey group at that stimulation type, and “^” indicates a statistically significant difference from the 0.5% honey group at that stimulation type. Values below Min LOD are shown as 0. The Kruskal-Wallis test was used with a Bonferroni multiple comparisons adjustment to establish significant differences between stimulation types at each honey level and between honey levels at each stimulation type (α = 0.05). Analytes are grouped by trend (A-D). Sample size = 3 independent replicates.
Box B contains analytes whose release was highest when in the presence of 0.5% honey. This trend was the strongest in the CCL3/MIP-1α, CCL4/MIP-1β, and CCL20/MIP-3α results, where release was minimal in all groups at 0.5% and 3% honey but significantly increased at 0.5% honey in all stimulated groups relative to the non-honey control. All three of these analytes had the greatest release in the TGF-β-stimulated group at 0.5% honey. In the CCL20/MIP-3α data, the TGF-β-stimulated group was about six times higher than the other groups at 0.5% honey, while in the CCL3/MIP-1α and CCL4/MIP-1β data the IL-4 and IL-4 + IL-13 groups had a little over half of the release of the TGF-β-stimulated group at 0.5% honey. Release of CXCL8/IL-8 and IL-1ra exceeded the Max LOD of the assay in the experimental groups cultured in 0.5% honey. CXCL8/IL-8 release was also significantly increased by 3% honey in the control group and IL-4 + IL-13 group relative to their non-honey release, while IL-1ra release was minimal in all groups in the presence of 3% honey. Significant release of IL-4 and CCL2/MCP-1 in the absence of honey occurred primarily in the IL-4 and the IL-4 + IL-13 groups, with negligible release in the TGF-β-stimulated cells. Although 0.5% honey significantly increased release of these analytes and 3% honey significantly decreased their release relative to non-honey controls, the trend of increased release in the IL-4 and IL-4 + IL-13 groups over the TGF-β group persisted at each honey level. Release of MMP-1, FGF-13, VEGF, and IL-1β only exceeded the Min LOD in the presence of 0.5% honey, with the release of FGF-13 and IL-1β relatively equivalent across all stimulated groups close to Min LOD. VEGF release was highest in the control cells at 0.5% honey, with levels decreased in each stimulated group relative to the control (significant in the IL-4 and IL-4 + IL-13 groups). In contrast, MMP-1 levels at 0.5% honey were higher in each stimulated group than in the non-stimulated control cells (significant in the TGF-β and IL-4 + Il-13 groups).
Box C displays the release of MMP-9 and IL-12 p70, neither of which were above the Min LOD of the assay in any group. Box D contains release results for Proteinase 3 and CCL5/RANTES, both of which had the highest release in the control cells at 0.5% honey. As the release of both of these analytes is significantly increased by 0.5% honey in the control cells relative to non-honey samples, but not in any of the stimulation groups, it is likely that however the honey is activating Proteinase 3 and CCL5/RANTES release, this mechanism is suppressed by each of the anti-inflammatory stimulators. Because this stimulation was done for 72 hours prior to the addition of the honey to the cells, it is likely that by the time the honey is added, the expression mechanisms for Proteinase 3 and CCL5/RANTES are suppressed.
Figure 2 displays the release results for each of these analytes at 24 hours, grouped by trend. Similar to the results seen at 3 hours, 3% honey decreased the release of all analytes except TNF-α and IL-8, which were increased relative to the non-honey samples. Likewise, the presence of 0.5% honey increased the results of most analytes relative to non-honey samples, including both pro-inflammatory and anti-inflammatory factors. As in Figure 1, TNF-α is grouped by itself in Box A and had significantly higher release in the presence of 3% honey than 0.5% honey or the absence of honey. At 3% honey, release was significantly less in the TGF-β-stimulated group than the control or the IL-4 and IL-4 + IL-13 groups. Similar to the results shown in Figure 1, these results suggest that TGF-β has a significant suppressive effect on the release of TNF-α at 3% honey, putting the cells in a state that is resistant to the secretion of TNF-α. Box B contains the release results for CXCL8/IL-8. As release exceeded the Max LOD in all groups at 0.5% and 3% honey, and in the IL-4-stimulated non-honey group, distinctions cannot be made between these groups. However, these results do indicate that both 0.5% and 3% honey increase CXCL8/IL-8 release relative to the non-honey groups in the control, TGF-β, and IL-4 + IL-13 groups. IL-4 stimulation also increased IL-8 release in the non-honey group relative to the non-stimulated control.
Figure 2.
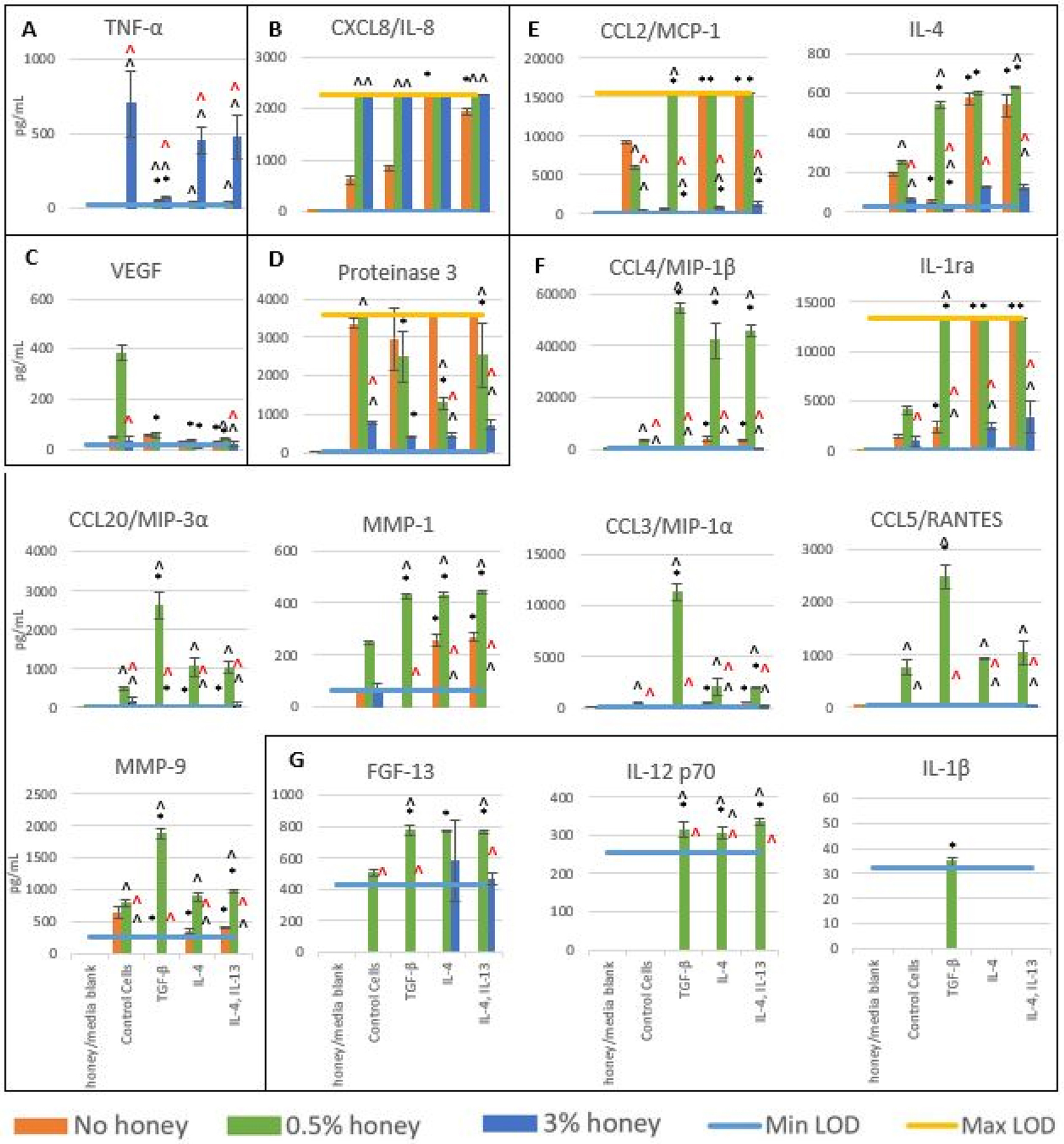
Release levels of cytokines, chemokines, and matrix-degrading enzymes in the absence of honey or presence of 0.5% or 3% honey at 24 hours. Bars indicate mean ± standard deviation. * indicates a statistically significant difference from the non-stimulated control at the respective honey level, ^ indicates a statistically significant difference from the non-honey group at that stimulation type, and “^” indicates a statistically significant difference from the 0.5% honey group at that stimulation type. Values below Min LOD are shown as 0. The Kruskal-Wallis test was used with a Bonferroni multiple comparisons adjustment to establish significant differences between stimulation types at each honey level and between honey levels at each stimulation type (α = 0.05). Analytes are grouped by trend (A-F). Sample size = 3 independent replicates.
VEGF release results are contained within Box C. These results indicate that 0.5% honey causes a non-statistically significant increase in VEGF release in the control cells relative to the non-honey samples (p=0.09), but this increase is not present in any of the stimulated groups. This trend is similar to that seen in the VEGF results in Figure 1, which suggests that the anti-inflammatory stimulators have a suppressive effect on the release of VEGF at 0.5% honey. Box D displays the release results for Proteinase 3. Although release exceeded the Max LOD for the 0.5% honey control cells and the non-honey IL-4 and IL-4 + IL-13 groups, some conclusions can still be drawn from these results. The presence of 0.5% honey significantly decreased release in the IL-4 and IL-4 + IL-13 groups, while 3% honey decreased release in all groups relative to the non-honey samples (significant in the control, IL-4, and IL-4 + IL-13 groups). These data suggest that the anti-inflammatory stimulation reverses the effect of 0.5% honey on Proteinase 3 release, but does not change the effect of 3% honey on the release of this analyte.
Box E contains the release results for analytes which had the highest release in the presence of 0.5% honey. The release of MMP-9, CCL5/RANTES, CCL3/MIP-1α, CCL20/MIP-3α, and MIP-1β were negligible in the non-honey and 3% honey groups, but had a statistically significant increase in release at 0.5% honey in most groups relative to the non-honey samples. The release profiles of MIP-1β and MMP-1 were similar in that there was elevated secretion from all three stimulation groups at 0.5% honey relative to the non-honey samples. Release of MMP-9, MIP-1α, CCL5/RANTES, and MIP-3α was highest in the TGF-β-stimulated group, suggesting that TGF-β enhances the effect of 0.5% honey on the release of these analytes. Release of CCL2/MCP-1, IL-4, IL-1ra, and MMP-1 was significantly increased in the non-honey samples by IL-4 and IL-4 + IL-13 stimulation relative to the non-stimulation controls. In particular, the CCL2/MCP-1, IL-4, and IL-1ra release profiles were similar in that their non-stimulated groups had low release, the TGF-β-stimulated group had elevated release only in the presence of 0.5% honey, the IL-4 and IL-4, IL-13 groups had elevated release at 0% and 0.5% honey, and 3% honey reduced all levels close to the Min LOD. TGF-β stimulation had little effect on IL-1ra and MMP-1 release in the absence of honey, and decreased CCL2/MCP-1, IL-4, and MMP-9 release relative to the non-stimulated control cells. 3% honey suppressed the release of all of the analytes in Box F, limiting CCL2/MCP-1, IL-4, and IL-1ra levels and keeping MIP-1β, CCL20/MIP-3α, MMP-1, CCL3/MIP-1α, CCL5/RANTES, and MMP-9 release below the Min LOD of the assay.
Box F displays the release results of FGF-13, IL-12 p70, and IL-1β, all of which were released at levels close to or below the Min LOD of the assay. The presence of 0.5% honey significantly increased release of FGF-13 and IL-12 p70 in the TGF-β, IL-4, and IL-4 + IL-13 groups relative to the control cells and the non-honey groups. IL-1β release was above the Min LOD only in the TGF-β-stimulated sample at 0.5% honey, and even this level was barely above the Min LOD, indicating that this release is negligible. The presence of 3% honey kept the release of IL-12 p70 and IL-1β below the Min LOD, and release of FGF-13 was only measurable in the IL-4 and IL-4 + IL-13 groups at this honey concentration, indicating that 3% honey suppresses the release of FGF-13, IL-12 p70, and IL-1β.
3.2. Primary human neutrophil experiments
Figure 3 displays the results of the primary human neutrophil experiments at both 3 and 24 hours of culture. The IL-1ra release results demonstrate that primary human neutrophils exhibit a dose-dependent release of Il-1ra in response to IL-4, which is significantly heightened in the presence of 0.5% honey but suppressed by the presence of 3% honey at both 3 and 24 hours. The VEGF results indicate that at 3 hours, both 0.5% and 3% honey significantly increase VEGF release in the 100 and 1000 ng/mL IL-4 groups (and in the 0 and 10 ng/mL IL-4 groups at 0.5% honey), but this release is still relatively low. At 24 hours, 0.5% honey causes a more substantial VEGF release (significant in the 10, 100, and 1000 ng/mL IL-4 groups), relative to both the non-honey control and 3% honey samples. The MMP-9 release results reveal that both 0.5% and 3% honey increased MMP-9 output (significant in all groups except 10 ng/mL IL-4 at 24 hours).
Figure 3.
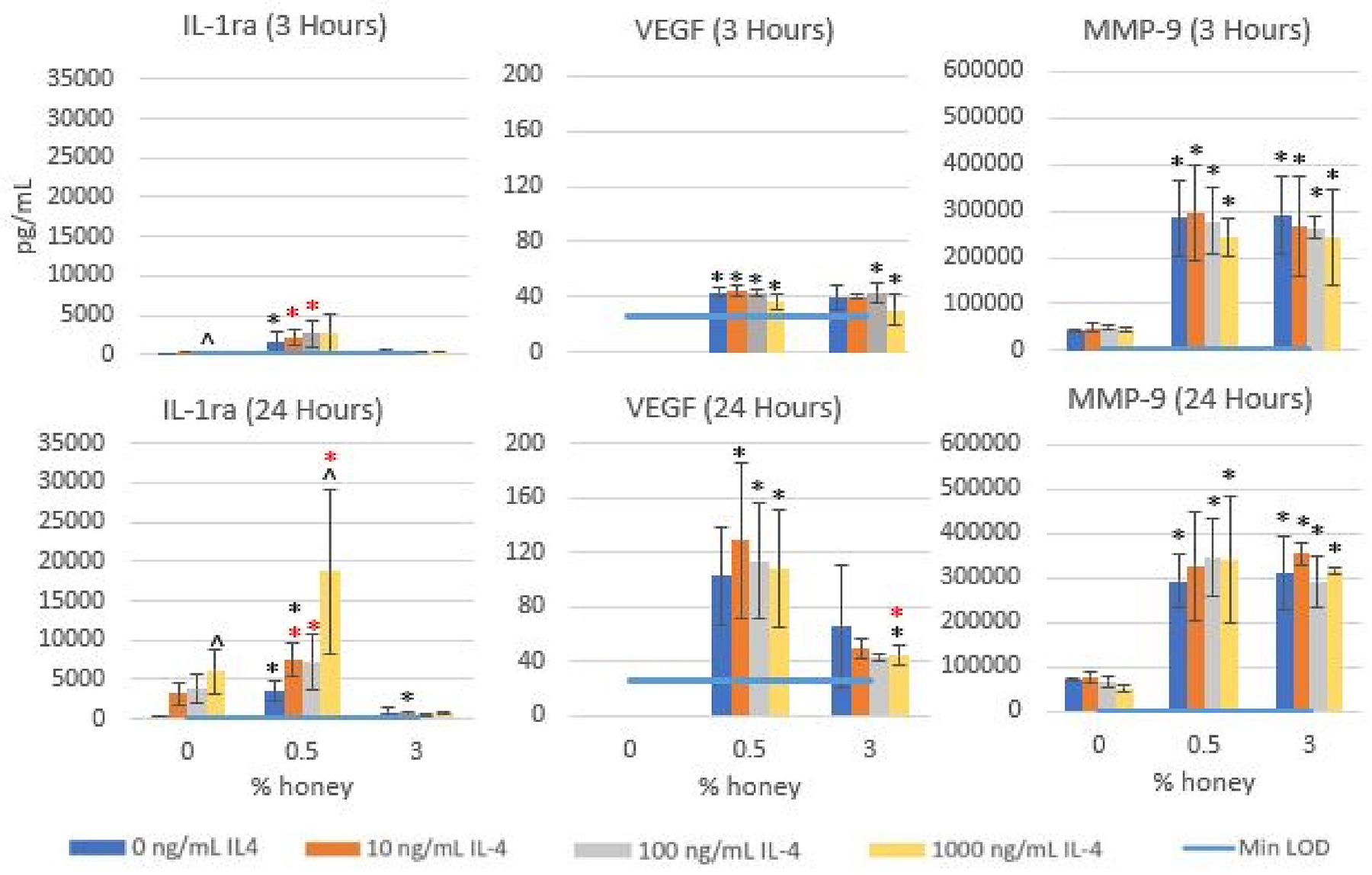
Release levels of MMP-9, VEGF, and IL-1ra in the absence of honey or presence of 0.5% or 3% honey at 3 and 24 hours under stimulation with 0, 10, 100, or 1000 ng/mL IL-4. Bars indicate mean ± standard deviation. * indicates a statistically significant difference from the non-honey control at the respective IL-4 level, * indicates a statistically significant difference from the 3% honey group at that IL-4 level, and “^” indicates a statistically significant difference from the 0 ng/mL IL-4 group at that honey level. Values below Min LOD are shown as 0. The Kruskal-Wallis test was used with a Bonferroni multiple comparisons adjustment to establish significant differences between stimulation types at each honey level and between honey levels at each stimulation type (α = 0.05). Sample size = 4 independent
4. Discussion
We have previously examined the effect of 0.5% and 3% Manuka honey on the cytokine profile of the dHL-60 model in the presence of various pro-inflammatory stimulators and found that the effect of the honey on this cytokine expression is dependent on the concentration of Manuka honey used. The presence of 0.5% honey decreased the release of TNF-α, IL-1β, MIP-1α, MIP-1β, IL-12 p70, MMP-9, MMP-1, FGF-13, IL-1ra, and IL-4, but increased MIP-3α, Proteinase 3, VEGF, and IL-8 release by the pro-inflammatory models. However, 3% honey increased the release of TNF-α, but decreased the release of all other analytes measured [48]. Majtan has previously theorized that the immunomodulatory effects of honey are dependent on the inflammatory state of the wound, with inflammation being increased in acute wounds but decreased in chronic wounds due to the presence of the honey [59]. This hypothesis would explain some of the effects that we previously observed, such as Manuka honey decreasing Proteinase 3 release in dHL-60s stimulated by LPS and fMLP, but would not explain the increase in TNF-α release in this inflammatory-stimulated model at 3% honey.
A similar difference between the effects of 0.5% honey and 3% honey was observed in results reported in this study in the anti-inflammatory-stimulated dHL-60 models. The 24-hour release results suggest that 3% honey has a pro-inflammatory effect by significantly increasing TNF-α and CXCL8/IL-8 output, although this honey level does significantly decrease the output of the pro-inflammatory signals CCL2/MCP-1, CCL4/MIP-1β, CCL20/MIP-3α, CCL3/MIP-1α, and CCL5/RANTES, as well as the matrix-degrading enzymes MMP-1, MMP-9, and Proteinase 3. In contrast, 0.5% honey increased the release of most analytes measured, including the inflammatory mediators CXCL8/IL-8, CCL2/MCP-1, CCL4/MIP-1β, CCL20/MIP-3α, CCL3/MIP-1α, IL-12 p70, and CCL5/RANTES, the matrix-degrading enzymes MMP-1 and MMP-9, and the anti-inflammatory mediators IL-4 and IL-1ra in the TGF-β-stimulated cells, the growth factor FGF-13 in all groups, and the growth factor VEGF in the non-stimulated control group. Although 0.5% honey decreased Proteinase 3 release in the stimulated groups, it increased Proteinase 3 release in the non-stimulated control cells. As such, the overall effect of the 0.5% honey concentration is to ramp up both pro-inflammatory and anti-inflammatory cytokine output.
There are several notable changes in expression profiles between the 3-hour and 24-hour timepoints. In the TNF-α graphs, the 3% honey control, IL-4, and IL-4, IL-13 groups show a substantial increase over time while the 0.5% honey groups remain static, indicating that 3% honey is much more effective at driving this release. IL-8 release exceeded the Max LOD in all 3% honey groups and the IL-4 non-honey group by 24 hours, and the IL-4, IL-13 group increased close to this level. The release of VEGF increased in the 0.5% honey non-stimulated control group, but remained static in the 0.5% honey stimulation groups, indicating that these signals are suppressing VEGF output. The IL-1ra graphs show a substantial increase in the non-honey IL-4 and IL4, IL-13 groups, indicating the effectiveness of these stimuli at driving IL-1ra release. MMP-9 had no measurable release at the 3-hour timepoint, indicating a latency in the release of this analyte. The Proteinase 3 graphs indicate that all non-honey groups and the 0.5% honey-stimulated groups substantially increased in Proteinase 3 release between 3 and 24 hours. In contrast, 3% honey repressed Proteinase 3 at both timepoints. There was also a substantial increase in CCL5/RANTES release in the 0.5% honey TGF-β group and a smaller increase in the 0.5% honey IL-4 and IL-4, IL-13 groups between the 3-hour and 24-hour timepoints. However, there was no change in CCL5/RANTES release in the non-honey or 3% honey groups over time, indicating that 0.5% honey is unique among these conditions in its stimulation of CCL5/RANTES release.
Some conclusions can be drawn about the relative impacts of TGF-β, IL-4, and IL-4 + IL-13 on the cytokine profile of the dHL-60 model in the absence of honey. In particular, the IL-4 and IL-4 + IL-13 groups had an increase in the release of CCL2/MCP-1, IL-4, Proteinase 3, IL-1ra, and MMP-1 in the absence of honey relative to the control cells. In contrast, the TGF-β-stimulated group had a significant decrease in CCL2/MCP-1, IL-4, Proteinase 3, MMP-1, and MMP-9 release in the absence of honey relative to the control cells, and also had lower IL-1ra release than the IL-4 and IL-4 + IL-13 groups. TGF-β also reduced the impact of 3% honey on TNF-α release, with much lower release at this honey level than the control cells and the IL-4 and IL-4 + IL-13 groups. However, TGF-β increased the release of CCL20/MIP-3α, CCL3/MIP-1α, CCL5/RANTES, and MMP-9 at 0.5% honey relative to the control cells and the IL-4 and IL-4 + IL-13 groups. The different effects between TGF-β and the other two stimulation groups can be partly explained by the fact that these stimulators act through different signaling pathways, as mentioned in the Introduction section. TGF-β activates the SMAD/RSMAD pathway, with IL-4 activating the Akt/PKB pathway and, in combination with IL-13, activating the Jak/STAT pathway. These pathways are regulated by different decoy receptors, ubiquitin-protein ligases, inhibitory proteins, kinases, and feedback-inhibition cycles, and activate different transcription factors which promote transcription of the relevant genes at different levels [60–62]. It is feasible that the effect of Manuka honey may be different on these different regulation mechanisms and transcription proteins, leading to different downstream effects on the cytokine release profile of these different stimulation groups.
The primary human neutrophil experiments provide a useful dataset to compare with the dHL-60 results and extrapolate more clinically-relevant findings. In both the dHL-60s and the primary human neutrophils, IL-4 stimulated the release of IL-1ra, and this release was increased by 0.5% honey but decreased by 3% honey. Similarly, at 24 hours both dHL-60s and primary human neutrophils exhibit an increase in VEGF release in response to 0.5% honey. One notable difference between these cell types was that in the dHL-60s, IL-4 stimulation blocked the ability of 0.5% honey to elicit an increase in VEGF release, while in the primary human neutrophils, IL-4 stimulation did not block this effect. The MMP-9 release results demonstrate that in both dHL-60s and primary human neutrophils, 0.5% honey caused a significant increase in MMP-9 release. However, the MMP-9 release of the dHL-60s was suppressed in the presence of 3% honey, while the MMP-9 release of the primary human neutrophils was stimulated by 3% honey to a level similar to the 0.5% honey samples. These differences in the response to 3% honey suggest that the dHL-60 cells behave differently from primary human neutrophils with regards to MMP-9 production. However, the similarities in the findings regarding the response to 0.5% honey illustrate the clinical benefit that a more precise fine-tuning the local honey concentration may have with regard to reducing inflammation via IL-1ra and stimulating angiogenesis via VEGF.
The ability of 0.5% to increase the release of some pro-inflammatory cytokines as well as some anti-inflammatory cytokines is similar to the effect of the IL-4 and IL-4, IL-13 groups in the absence of honey. As shown in Figure 2, these stimuli increase the release of the pro-inflammatory signals IL-8 and CCL2/MCP-1, as well as the matrix-reorganization enzyme MMP-1 and the anti-inflammatory signals IL-4 and IL-1ra. This partial pro-inflammatory effect of IL-4 has been previously observed in literature [63]. Despite this effect, IL-4 has an overall effect of decreasing wound inflammation, and, like Manuka honey, topical application of IL-4 has been shown to increase the rate of wound closure [64]. As such, it can be inferred that the honey-driven increase in the release of some pro-inflammatory cytokines is not problematic within the wound site. It should also be noted that the osmotic gradient caused by the high sugar content of the honey creates a net fluid flow from the wound bed towards the surface which would reduce the overall level of these cytokines in the wound environment. In addition to its direct application to a wound, Manuka honey is also being investigated as a tissue template additive to reduce inflammation in the area around the template and promote cellular ingrowth and tissue-template integration [2, 3, 65, 66]. In this form of use, these honey-doped templates will be implanted within the body, with cells coming into direct contact with the honey within the microenvironment of the template. With the implantation of these templates into the body, the osmotic gradient will cause fluid to flow into the interior of the template, rather than to the surface of a wound, which could cause higher levels of these honey-induced cytokines within the template microenvironment. As such, in vivo testing of these templates should involve assaying cytokine levels within the template interior to confirm that this microenvironment is not overly inflammatory.
The current predominant method of use of Manuka honey in the clinic is to spread non-diluted Manuka honey onto a wound site, sometimes covering the honey with a bandage after application [15–17, 67–70]. This usage of non-diluted Manuka honey has been shown to effectively promote healing in vivo, but in vitro testing has indicated that concentrations above 5% v/v Manuka honey are cytotoxic [27, 30]. Corroborating experiments have indicated that non-diluted Manuka honey can kill cells and cause tissue damage when applied to ovine frontal sinuses and chinchilla ears [13, 29]. We have previously theorized that when non-diluted Manuka honey is applied to a wound site, it creates a gradient with a “zone of death” at the top of the wound killing human cells as well as infiltrating bacteria, and lower honey concentrations deeper into the wound environment [48]. In the context of honey-containing tissue templates, cells will come into direct contact with the honey within the template microenvironment. To avoid cytotoxic effects on these cells, groups fabricating these honey-doped templates have focused on tailoring the Manuka honey content and release rate to elicit the desired cell responses while staying below the cytotoxic limit [2, 71–73]. As such, the concentrations of 0.5% and 3% Manuka honey which were tested in this study are in the general range of the honey concentrations which will be encountered by a neutrophil within the wound bed or in contact with a honey-doped tissue engineering template. The findings of this study are thus relevant to the design of such templates and can provide a general understanding of how these honey concentrations will affect neutrophils during the resolution of inflammation and transition to the pro-healing, pro-angiogenesis phase of the wound. In particular, the TNF-α results suggest that template-released honey should be maintained below 3% v/v in the surrounding tissue to avoid stimulating inflammation, and a target of 0.5% v/v would be optimal to elicit IL-1ra and VEGF release from neutrophils.
5. Conclusion
Manuka honey’s effect on dHL-60 and primary human neutrophil cytokine release under anti-inflammatory stimulation is dependent on the concentration of Manuka honey used. The presence of 3% honey & IL-4 caused a significant increase in the release of TNF-α and CXCL8/IL-8 while reducing the release of all other analytes measured regardless of the anti-inflammatory stimulator that was used. In contrast, 0.5% honey significantly increased the release of CXCL8/IL-8, CCL2/MCP-1, CCL4/MIP-1β, CCL20/MIP-3α, IL-4, IL-1ra, and FGF-13 in at least one stimulation group while reducing the release of Proteinase 3 in the anti-inflammatory-stimulated models at 24 hours. The results also indicate that at 0.5% honey at 24 hours, TGF-β-stimulation significantly increased the release of CCL20/MIP-3α, CCL3/MIP-1α, CCL5/RANTES, and MMP-9 relative to the other groups. Additionally, in the absence of honey, TGF-β stimulation significantly reduced IL-4 and MMP-9 release and caused a non-significantly decreasing trend in the release of CCL2/MCP-124 hours relative to the control cells and the other anti-inflammatory stimulation groups. These results indicate the importance of tailoring the release rate of Manuka honey from tissue templates to elicit the desired effects on the neutrophils in the wound during the resolution of inflammation. Future work will investigate the use of Manuka honey-releasing templates in modulating neutrophil inflammatory behavior and NETosis to improve tissue-template integration [57]. Future experimentation should also focus on identifying the honey components responsible for these results, perhaps through the use of flavonoid-doped artificial honey solutions. Additionally, levels of honey between 0.5% and 3% v/v should be tested to further define the response continuum of the neutrophil cytokine expression profile to Manuka honey.
Highlights.
Manuka honey alters the secretome of an anti-inflammatory dHL-60 neutrophil model
0.5% honey increases both pro-inflammatory and anti-inflammatory signal release
3% honey reduces all release except TNF-α and IL-8
In primary human neutrophils, 0.5% honey increases IL-1ra, VEGF, and MMP-9 release
6. Acknowledgements
The authors thank the University of Memphis and the University of Tennessee Health Science Campus, as well as the Van Vleet Memorial Doctoral Award for their support of this project. Research was supported by the National Institute of Biomedical Imaging and Bioengineering of the National Institutes of Health under Award Number R15EB022345.
Abbreviations
- Akt
a serine-threonine kinase
- ATCC
American Type Culture Collection
- dHL-60
differentiated HL-60
- CCL
C-C motif ligand
- CXCL
C-X-C motif ligand
- DAPI
4’6-diamidino-2-phenylindole
- DMSO
dimethylsulfoxide
- ECM
extracellular matrix
- FBS
fetal bovine serum
- FGF
fibroblast growth factor
- HPLC-MS
high performance liquid chromatography – mass spectroscopy
- IL
interleukin
- Jak
janus kinase
- LDL
low-density lipoprotein
- MIP
macrophage inflammatory protein
- MMP
matrix metalloproteinase
- NF-κB
nuclear transcription factor κB
- Nrf2
nuclear factor erythroid 2
- PKB
protein kinase B
- SMAD, STAT
signal transducer and activator of transcription; small mothers against decapentaplegic
- RANTES
regulated on activation, normal T cell expressed and secreted
- TGF-β
transforming growth factor β
- TNF
tumor necrosis factor
- VEGF
vascular endothelial growth factor
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Conflict of Interest
The authors declare no conflict of interest.
8 References
- 1.Molan PC 2002. Re-introducing honey in the management of wounds and ulcers-theory and practice, Ostomy Wound Manage. 48, 28–40. [PubMed] [Google Scholar]
- 2.Minden-Birkenmaier BA, Neuhalfen RM, Janowiak BE, Sell SA 2015. Preliminary Investigation and Characterization of Electrospun Polycaprolactone and Manuka Honey Scaffolds for Dermal Repair, Eng Fiber Fabr. 10. [Google Scholar]
- 3.Minden-Birkenmaier B and Bowlin G 2018. Honey-based templates in wound healing and tissue engineering, Bioengineering. 5, 46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Alvarez-Suarez JM, Giampieri F, Cordero M, Gasparrini M, Forbes-Hernández TY, Mazzoni L, Afrin S, Beltrán-Ayala P, González-Paramás AM, Santos-Buelga C 2016. Activation of AMPK/Nrf2 signalling by Manuka honey protects human dermal fibroblasts against oxidative damage by improving antioxidant response and mitochondrial function promoting wound healing, J. Funct. Foods 25, 38–49. [Google Scholar]
- 5.Ranzato E, Martinotti S, Burlando B 2013. Honey exposure stimulates wound repair of human dermal fibroblasts, Burns trauma. 1, 32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Suguna L, Chandrakasan G, Joseph KT 1992. Influence of honey on collagen metabolism during wound healing in rats, J. Clin. Biochem. Nutr 13, 7–12. [Google Scholar]
- 7.Oryan A, Alemzadeh E, Moshiri A 2016. Biological properties and therapeutic activities of honey in wound healing: a narrative review and meta-analysis, Journal of tissue viability. 25, 98–118. [DOI] [PubMed] [Google Scholar]
- 8.Jenkins R, Burton N, Cooper R 2011. Manuka honey inhibits cell division in methicillin-resistant Staphylococcus aureus, Journal of Antimicrobial Chemotherapy. 66, 2536–2542. [DOI] [PubMed] [Google Scholar]
- 9.Cooper R, Molan P, Harding K 2002. The sensitivity to honey of Gram-positive cocci of clinical significance isolated from wounds, Journal of Applied microbiology. 93, 857–863. [DOI] [PubMed] [Google Scholar]
- 10.Molan P, Cooper R, Molan P, White R 2009. Why honey works, Honey in modern wound management. 9, 36–37. [Google Scholar]
- 11.Molan PC 2006. The evidence supporting the use of honey as a wound dressing, Int J Low Extrem Wounds. 5, 40–54. [DOI] [PubMed] [Google Scholar]
- 12.Alvarez-Suarez J, Gasparrini M, Forbes-Hernández T, Mazzoni L, Giampieri F 2014. The composition and biological activity of honey: a focus on Manuka honey, Foods. 3, 420–432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Oryan A and Zaker S 1998. Effects of topical application of honey on cutaneous wound healing in rabbits, Vet Med A. 45, 181–188. [DOI] [PubMed] [Google Scholar]
- 14.Moghazy A, Shams M, Adly O, Abbas A, El-Badawy M, Elsakka D, Hassan S, Abdelmohsen W, Ali O, Mohamed B 2010. The clinical and cost effectiveness of bee honey dressing in the treatment of diabetic foot ulcers, Diabetes Res. Clin. Pract 89, 276–281. [DOI] [PubMed] [Google Scholar]
- 15.Gethin GT, Cowman S, Conroy RM 2008. The impact of Manuka honey dressings on the surface pH of chronic wounds, Int. Wound J 5, 185–194. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 16.Visavadia BG, Honeysett J, Danford MH 2008. Manuka honey dressing: An effective treatment for chronic wound infections, Br. J. Oral Maxillofac. Surg 46, 55–56. [DOI] [PubMed] [Google Scholar]
- 17.Al-Waili N, Salom K, Al-Ghamdi AA 2011. Honey for wound healing, ulcers, and burns; data supporting its use in clinical practice, Sci. World J 11, 766–787. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.El-Guendouz S, Al-Waili N, Aazza S, Elamine Y, Zizi S, Al-Waili T, Al-Waili A, Lyoussi B 2017. Antioxidant and diuretic activity of co-administration of Capparis spinosa honey and propolis in comparison to furosemide, Asian Pacific journal of tropical medicine. 10, 974–980. [DOI] [PubMed] [Google Scholar]
- 19.Deng J, Liu R, Lu Q, Hao P, Xu A, Zhang J, Tan J 2018. Biochemical properties, antibacterial and cellular antioxidant activities of buckwheat honey in comparison to manuka honey, Food Chem. 252, 243–249. [DOI] [PubMed] [Google Scholar]
- 20.Ahmed S and Othman NH 2013. Review of the medicinal effects of tualang honey and a comparison with manuka honey, The Malaysian journal of medical sciences: MJMS. 20, 6. [PMC free article] [PubMed] [Google Scholar]
- 21.Sherlock O, Dolan A, Athman R, Power A, Gethin G, Cowman S, Humphreys H 2010. Comparison of the antimicrobial activity of Ulmo honey from Chile and Manuka honey against methicillin-resistant Staphylococcus aureus, Escherichia coli and Pseudomonas aeruginosa, BMC complementary and alternative medicine. 10, 1–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Hillitt K, Jenkins R, Spiller OB, Beeton ML 2017. Antimicrobial activity of Manuka honey against antibiotic-resistant strains of the cell wall-free bacteria Ureaplasma parvum and Ureaplasma urealyticum, Lett. Appl. Microbiol 64, 198–202. [DOI] [PubMed] [Google Scholar]
- 23.Atrott J and Henle T 2009. Methylglyoxal in manuka honey-correlation with antibacterial properties, Czech J Food Sci. 27, S163–S165. [Google Scholar]
- 24.Blair S, Cokcetin N, Harry E, Carter D 2009. The unusual antibacterial activity of medical-grade Leptospermum honey: antibacterial spectrum, resistance and transcriptome analysis, Eur. J. Clin. Microbiol. Infect. Dis 28, 1199–1208. [DOI] [PubMed] [Google Scholar]
- 25.Caskey PR (2010). Use of honey in dressings. Google Patents. [Google Scholar]
- 26.Minden-Birkenmaier BA and Bowlin GL 2018. Honey-based templates in wound healing and tissue engineering, Bioengineering. 5, 46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Sell SA, Wolfe PS, Spence AJ, Rodriguez IA, McCool JM, Petrella RL, Garg K, Ericksen JJ, Bowlin GL 2012. A preliminary study on the potential of manuka honey and platelet-rich plasma in wound healing, Int. J. Biomater 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Aron M, Akinpelu OV, Dorion D, Daniel S 2012. Otologic safety of manuka honey, Otolaryngol. Head Neck Surg 41. [PubMed] [Google Scholar]
- 29.Paramasivan S, Drilling AJ, Jardeleza C, Jervis-Bardy J, Vreugde S, Wormald PJ 2014. Methylglyoxal-augmented manuka honey as a topical anti-Staphylococcus aureus biofilm agent: safety and efficacy in an in vivo model, Int Forum Allergy Rhinol. 4, 187–195. [DOI] [PubMed] [Google Scholar]
- 30.Minden-Birkenmaier BA, Cherukuri K, Smith RA, Radic MZ, Bowlin GL 2019. Manuka Honey Modulates the Inflammatory Behavior of a dHL-60 Neutrophil Model Under the Cytotoxic Limit, Int. J. Biomater [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Ng LG, Qin JS, Roediger B, Wang Y, Jain R, Cavanagh LL, Smith AL, Jones CA, De Veer M, Grimbaldeston MA 2011. Visualizing the neutrophil response to sterile tissue injury in mouse dermis reveals a three-phase cascade of events, Journal of Investigative Dermatology. 131, 2058–2068. [DOI] [PubMed] [Google Scholar]
- 32.Selders GS, Fetz AE, Radic MZ, Bowlin GL 2017. An overview of the role of neutrophils in innate immunity, inflammation and host-biomaterial integration, Regenerative biomaterials. 4, 55–68. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Borregaard N and Cowland JB 1997. Granules of the human neutrophilic polymorphonuclear leukocyte, Blood. 89, 3503–3521. [PubMed] [Google Scholar]
- 34.Neely CJ, Kartchner LB, Mendoza AE, Linz BM, Frelinger JA, Wolfgang MC, Maile R, Cairns BA 2014. Flagellin treatment prevents increased susceptibility to systemic bacterial infection after injury by inhibiting anti-inflammatory IL-10+ IL-12-neutrophil polarization, PloS one. 9, e85623. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Wilgus TA, Roy S, McDaniel JC 2013. Neutrophils and wound repair: positive actions and negative reactions, Advances in wound care. 2, 379–388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Diegelmann RF and Evans MC 2004. Wound healing: an overview of acute, fibrotic and delayed healing, Front Biosci. 9, 283–289. [DOI] [PubMed] [Google Scholar]
- 37.Yager DR and Nwomeh BC 1999. The proteolytic environment of chronic wounds, Wound Repair Regen. 7, 433–441. [DOI] [PubMed] [Google Scholar]
- 38.Martin P and Leibovich SJ 2005. Inflammatory cells during wound repair: the good, the bad and the ugly, Trends in cell biology. 15, 599–607. [DOI] [PubMed] [Google Scholar]
- 39.Dalli J, Norling LV, Renshaw D, Cooper D, Leung K-Y, Perretti M 2008. Annexin 1 mediates the rapid anti-inflammatory effects of neutrophil-derived microparticles, Blood. 112, 2512–2519. [DOI] [PubMed] [Google Scholar]
- 40.Gasser O and Schifferli JA 2004. Activated polymorphonuclear neutrophils disseminate anti-inflammatory microparticles by ectocytosis, Blood. 104, 2543–2548. [DOI] [PubMed] [Google Scholar]
- 41.Kolaczkowska E and Kubes P 2013. Neutrophil recruitment and function in health and inflammation, Nat. Rev. Immunol 13, 159. [DOI] [PubMed] [Google Scholar]
- 42.Collins SJ 1987. The HL-60 promyelocytic leukemia cell line: proliferation, differentiation, and cellular oncogene expression, Blood. 70, 1233–1244. [PubMed] [Google Scholar]
- 43.Collins S, Ruscetti F, Gallagher R, Gallo R 1979. Normal functional characteristics of cultured human promyelocytic leukemia cells (HL-60) after induction of differentiation by dimethylsulfoxide, Journal of Experimental Medicine. 149, 969–974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Santos-Beneit AM and Mollinedo F 2000. Expression of genes involved in initiation, regulation, and execution of apoptosis in human neutrophils and during neutrophil differentiation of HL-60 cells, Journal of leukocyte biology. 67, 712–724. [DOI] [PubMed] [Google Scholar]
- 45.Mollinedo F, Santos-Beneit AM, Gajate C (1998) The human leukemia cell line HL-60 as a cell culture model to study neutrophil functions and inflammatory cell responses. In Animal cell culture techniquesSpringer; 264–297. [Google Scholar]
- 46.Trellakis S, Farjah H, Bruderek K, Dumitru C, Hoffmann T, Lang S, Brandau S 2011. Peripheral blood neutrophil granulocytes from patients with head and neck squamous cell carcinoma functionally differ from their counterparts in healthy donors, Int. J. Immunopathol. Pharmacol 24, 683–693. [DOI] [PubMed] [Google Scholar]
- 47.Naegelen I, Plancon S, Nicot N, Kaoma T, Muller A, Vallar L, Tschirhart EJ 2015. An essential role of syntaxin 3 protein for granule exocytosis and secretion of IL-1α, IL-1β, IL-12b, and CCL4 from differentiated HL-60 cells, Journal of leukocyte biology. 97, 557–571. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Minden-Birkenmaier B, Meadows M, Cherukuri K, Smeltzer MP, Radic M, Bowlin G, Smith R 2019. The Effect of Manuka Honey on dHl-60 Cytokine, Chemokine, and Matrix-Degrading Enzyme Release under Inflammatory Conditions, Med One. 4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Heldin C-H, Miyazono K, Ten Dijke P 1997. TGF-β signalling from cell membrane to nucleus through SMAD proteins, Nature 390, 465. [DOI] [PubMed] [Google Scholar]
- 50.Scheid MP and Duronio V 1998. Dissociation of cytokine-induced phosphorylation of Bad and activation of PKB/akt: involvement of MEK upstream of Bad phosphorylation, Proceedings of the National Academy of Sciences. 95, 7439–7444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Albanesi C, Fairchild HR, Madonna S, Scarponi C, De Pità O, Leung DY, Howell MD 2007. IL-4 and IL-13 negatively regulate TNF-α-and IFN-γ-induced β-defensin expression through STAT-6, suppressor of cytokine signaling (SOCS)-1, and SOCS-3, J. Immunol 179, 984–992. [DOI] [PubMed] [Google Scholar]
- 52.Marie C, Pitton C, Fitting C, Cavaillon J-M 1996. IL-10 and IL-4 synergize with TNF-α to induce IL-1ra production by human neutrophils, Cytokine. 8, 147–151. [DOI] [PubMed] [Google Scholar]
- 53.Shen L, Smith J, Shen Z, Eriksson M, Sentman C, Wira C 2007. Inhibition of human neutrophil degranulation by transforming growth factor-β1, Clin. Exp. Immunol 149, 155–161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Gallagher R, Collins S, Trujillo J, McCredie K, Ahearn M, Tsai S, Metzgar R, Aulakh G, Ting R, Ruscetti F 1979. Characterization of the continuous, differentiating myeloid cell line (HL-60) from a patient with acute promyelocytic leukemia, Blood. 54, 713–733. [PubMed] [Google Scholar]
- 55.Martin S, Bradley J, Cotter T 1990. HL-60 cells induced to differentiate towards neutrophils subsequently die via apoptosis, Clin. Exp. Immunol 79, 448–453. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Collins SJ, Ruscetti FW, Gallagher RE, Gallo RC 1978. Terminal differentiation of human promyelocytic leukemia cells induced by dimethyl sulfoxide and other polar compounds, Proceedings of the National Academy of Sciences. 75, 2458–2462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Fetz AE, Neeli I, Rodriguez IA, Radic MZ, Bowlin GL 2017. Electrospun template architecture and composition regulate neutrophil NETosis in vitro and in vivo, Tissue Engineering Part A. 23, 1054–1063. [DOI] [PubMed] [Google Scholar]
- 58.Berton G, Zeni L, Cassatella MA, Rossi F 1986. Gamma interferon is able to enhance the oxidative metabolism of human neutrophils, Biochem. Biophys. Res. Commun 138, 1276–1282. [DOI] [PubMed] [Google Scholar]
- 59.Majtan J 2014. Honey: an immunomodulator in wound healing, Wound Repair Regen. 22, 187–192. [DOI] [PubMed] [Google Scholar]
- 60.Moustakas A, Souchelnytskyi S, Heldin C-H 2001. Smad regulation in TGF-β signal transduction, J. Cell Sci 114, 4359–4369. [DOI] [PubMed] [Google Scholar]
- 61.Hemmings BA and Restuccia DF 2012. Pi3k-pkb/akt pathway, Cold Spring Harbor perspectives in biology. 4, a011189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Shuai K and Liu B 2003. Regulation of JAK-STAT signalling in the immune system, Nat. Rev. Immunol 3, 900. [DOI] [PubMed] [Google Scholar]
- 63.Lavoie-Lamoureux A, Moran K, Beauchamp G, Mauel S, Steinbach F, Lefebvre-Lavoie J, Martin JG, Lavoie J-P 2010. IL-4 activates equine neutrophils and induces a mixed inflammatory cytokine expression profile with enhanced neutrophil chemotactic mediator release ex vivo, American Journal of Physiology-Lung Cellular Molecular Physiology. 299, L472–L482. [DOI] [PubMed] [Google Scholar]
- 64.Salmon-Ehr V, Ramont L, Godeau G, Birembaut P, Guenounou M, Bernard P, Maquart F-X 2000. Implication of interleukin-4 in wound healing, Lab. Invest 80, 1337. [DOI] [PubMed] [Google Scholar]
- 65.Hilliard G, DeClue CE, Minden-Birkenmaier BA, Dunn AJ, Sell SA, Shornick LP 2019. Preliminary investigation of honey-doped electrospun scaffolds to delay wound closure, J. Biomed. Mater. Res. B [DOI] [PubMed] [Google Scholar]
- 66.Hixon KR, Lu T, McBride-Gagyi SH, Janowiak BE, Sell SA 2017. A Comparison of Tissue Engineering Scaffolds Incorporated with Manuka Honey of Varying UMF, BioMed Res. Int. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Efem SE 1993. Recent advances in the management of Fournier’s gangrene: preliminary observations, Surgery. 113, 200–204. [PubMed] [Google Scholar]
- 68.Gethin G and Cowman S 2005. Case series of use of Manuka honey in leg ulceration, Int. Wound J 2, 10–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Gethin G and Cowman S 2008. Bacteriological changes in sloughy venous leg ulcers treated with manuka honey or hydrogel: an RCT, Wound Care. 17, 241–247. [DOI] [PubMed] [Google Scholar]
- 70.Efem S 1988. Clinical observations on the wound healing properties of honey, Br. J. Surg 75, 679–681. [DOI] [PubMed] [Google Scholar]
- 71.Kadakia PU, Growney Kalaf EA, Dunn AJ, Shornick LP, Sell SA 2018. Comparison of silk fibroin electrospun scaffolds with poloxamer and honey additives for burn wound applications, J. Bioact. Compat. Polym 33, 79–94. [Google Scholar]
- 72.Yang X, Fan L, Ma L, Wang Y, Lin S, Yu F, Pan X, Luo G, Zhang D, Wang H 2017. Green electrospun Manuka honey/silk fibroin fibrous matrices as potential wound dressing, Mater. Des 119, 76–84. [Google Scholar]
- 73.Wang P and He J-H 2013. Electrospun polyvinyl alcohol-honey nanofibers, Thermal Science. 17, 1549–1550. [Google Scholar]