
Figure 6. NR enhances circadian transcription and evening locomotor activity in old mice.
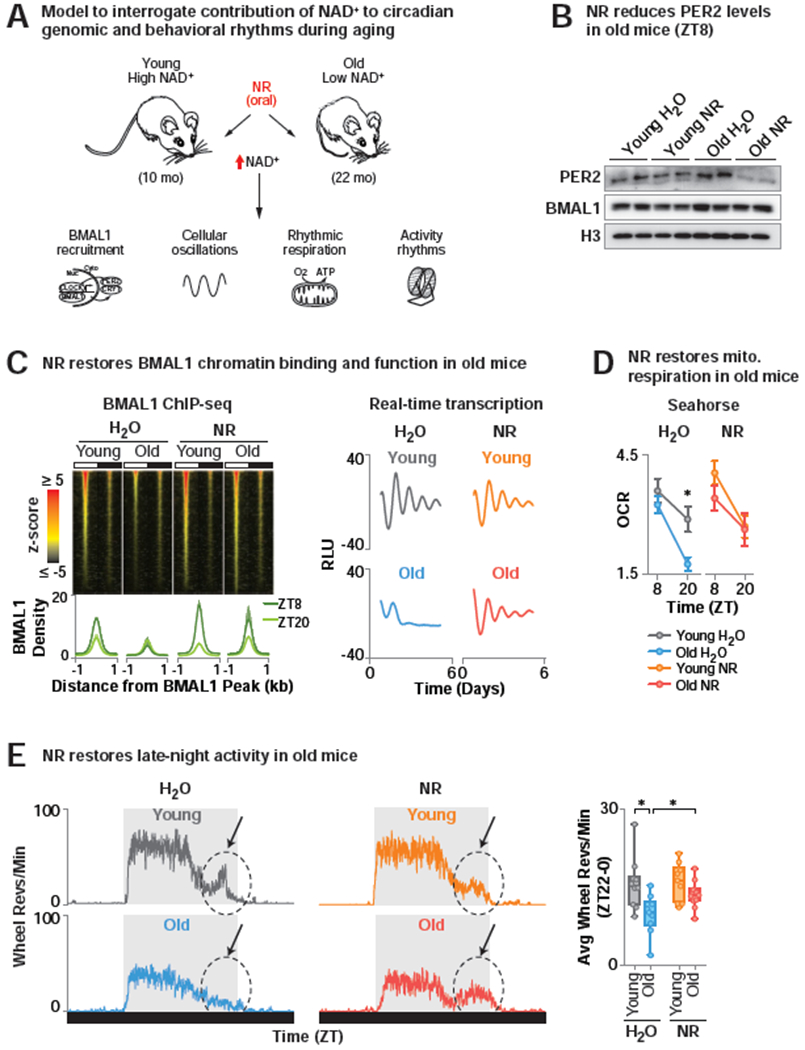
(A) Model to interrogate the contribution of NAD+ to circadian genomic and behavioral rhythmic phenotypes during aging. (B) PER2 and BMAL1 levels in liver (ZT8) of 10 month (“young”) and 22 month (“old”) WT mice treated either with NR-supplemented or regular drinking water for 6 months. (C) (Top) BMAL1 ChIP-seq in liver (ZT8 and ZT20). Z-score normalized average tag-densities for genomic regions corresponding to the center of each BMAL1 peak identified in old mice treated with NR ± 1 kb and sorted by size (n=2) (Bottom) Average BMAL1 tag-density and standard error for both replicates within each group. (right) Average background-subtracted PER2::LUC oscillations from soleus of young and old water- and NR-treated mice transgenic for the Per2::Luciferase reporter monitored ex vivo in a lumicycle (n=3). (D) Oxygen consumption rates (OCR) of mitochondria isolated from young and old, water- and NR-treated liver (ZT8 and ZT20) by Seahorse Bioanalyzer. FCCP-treated respiration with palmitate relative to basal (*p<0.05) (n=4-5). (E) (Left) Representative average 24 hr profile of ad lib wheel running activity of young and old water- and NR-treated WT mice. Arrow denotes ‘late-night activity’. (Right) Quantification of average wheel revolutions/min in the late night (ZT22-0) (n=8-10) (*p<0.05). See also Figure S4 and Table S5.