

Abstract
BACKGROUND
The endocannabinoid system (ECS) consists of the cannabinoid receptors CB1 and CB2, the main endocannabinoids anandamide (AEA) and 2-arachidonoylglycerol (2-AG) and their metabolic enzymes N-acylphosphatidylethanolamine-specific phospholipase D, fatty acid amide hydrolase, diacylglycerol lipase and monoacylglycerol lipase. This system is involved in the modulation of essential physiological processes. Its role in the reproductive system has become significantly important in recent years, given its major role in events such as gametogenesis, decidualisation, implantation and placentation.
OBJECTIVE AND RATIONALE
In this paper, we review the literature and summarize the role of the ECS elements in reproduction and their potential as early markers for diagnosis of reproductive disorders or as pharmacological targets for treatment.
SEARCH METHODS
Original research and review papers published from 1964 to June 2019 were selected in terms of relevance, reliability and quality by searching PubMed, MEDLINE and Web of Science, using the following search terms: endocannabinoid system and endometriosis; endocannabinoid system and ectopic pregnancy; endocannabinoid system and miscarriage; endocannabinoid system and pre-eclampsia; endocannabinoid system and endometrial cancer; endocannabinoid system and reproduction; endocannabinoid, endometrium; placenta; N-acylethanolamines; anandamide; 2-arachidonoylglycerol; and cannabinoids.
OUTCOMES
This review demonstrates relevant information concerning ECS alterations in endometriosis, ectopic pregnancy, miscarriage, pre-eclampsia and endometrial cancer. We highlight the importance of the endocannabinoids in endometrial and placental physiology and pathophysiology, from studies in vitro and in vivo and in clinical observations. The most studied of the endogenous cannabinoids is AEA. The levels of AEA were increased in plasma of patients with endometriosis and miscarriage, as well as in the fallopian tube of women with ectopic pregnancy and in endometrial biopsies of endometrial cancer. Changes in the pattern of expression of the cannabinoid receptor CB1 were also observed in endometrial biopsies of endometriosis, fallopian tube and decidua of patients with ectopic pregnancy and pre-eclamptic placenta. Moreover, alterations in CB2 expression have been reported in association with endometrial cancer. In general, studies on the cannabinoid signalling through CB2 and on the biological activities of the other major endocannabinoid, namely 2-AG, as well as its metabolic enzymes are scarce and avidly required.
WIDER IMPLICATIONS
The pathophysiological mechanisms involved in the described endometrial and placental pathologies are still unclear and lack the means for an early diagnosis. Based on current evidence, though alterations in ECS are demonstrated at tissue level, it is difficult to associate plasmatic changes in AEA with specific endometrial and placental diseases. Thus, pairing alterations in AEA levels with 2-AG and/or other endocannabinoid-like molecules may provide more accurate and early diagnoses. In addition, patients may benefit from new therapies that target the ECS and endocannabinoid signalling.
Keywords: endocannabinoid system, endocannabinoids, endometrium, placenta, pregnancy, endometriosis, ectopic pregnancy, miscarriage, pre-eclampsia, endometrial cancer
Introduction
Cannabis sativa has been used for centuries for a plethora of purposes. Nowadays, its consumption is growing due to the increasing legalisation for therapeutic applications. Despite its popularity, it was only in 1964 that the main psychoactive compound of C. sativa, the cannabinoid Δ9-tetrahydrocannabinol (THC), was isolated and characterized (Gaoni and Mechoulam, 1964). Many years later, its molecular targets were discovered. The cannabinoid receptor 1 (CB1) was identified in 1990 (Matsuda et al., 1990) followed by the cannabinoid receptor 2 (CB2) (Munro et al., 1993). Their crystallographic structure has, however, only recently been described (Hua et al., 2016; Shao et al., 2016; Li et al., 2019). Both receptors belong to the seven-transmembrane G protein-coupled receptor superfamily and are expressed throughout the central nervous system (Zou and Kumar, 2018) and in peripheral organ systems such as the immune system (Pandey et al., 2009), the gastrointestinal tract (DiPatrizio, 2016), skin (Rio et al., 2018) and reproductive tissues (Rossato et al., 2008; Walker et al., 2019). The identification of the cannabinoid receptors led to the discovery of the endogenous ligands, termed endocannabinoids (eCBs). The first endocannabinoid to be isolated was the ethanolamide of arachidonic acid, arachidonoylethanolamine, also named anandamide (AEA) after ‘ananda’, which means bliss in Sanskrit (Devane et al., 1992). Shortly after, another derivative of arachidonic acid, 2-arachidonoylglycerol (2-AG), was described (Mechoulam et al., 1995).
The Endocannabinoid System
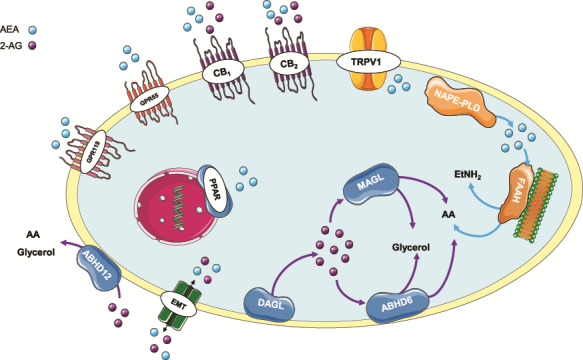
The endocannabinoid system (ECS) is a lipid signalling system composed of the cannabinoid receptors, the eCBs and their main metabolic enzymes (Fig. 1). AEA is produced from membrane phospholipids by the action of N-acetyltransferase followed by N-acylphosphatidylethanolamine-specific phospholipase D (NAPE-PLD) (Di Marzo et al., 1994), while 2-AG is synthesized by the enzyme 1,2-diacylglycerol lipase α/β (DAGL) (Sugiura et al., 1995). However, alternative routes of 2-AG synthesis may exist since it was reported that DAGL−/− mice presented only an 80% reduction of 2-AG levels (Alger and Kim, 2011). Early studies on the eCBs suggested that these were synthesized on demand, meaning that they are not stored but synthesized locally upon need; however, in 2014, it was suggested that the eCBs are stored in intracellular lipid droplets referred to as adiposomes, allowing intracellular accumulation (Maccarrone, 2009; Fezza et al., 2014). The actions of eCBs on their receptors are terminated by degradation through their respective hydrolyzing enzymes. AEA is inactivated by fatty acid amide hydrolase FAAH (Cravatt et al., 1996) mainly located in the intracellular membranes, while the main catabolic route for 2-AG is through monoacylglycerol lipase (MAGL), an enzyme associated with the inner leaflet of plasma membranes (Dinh et al., 2002), and, to a lesser extent, by the recently discovered α/β-hydrolase-6 and -12 (ABHD6 and ABHD12) (Blankman et al., 2007; Chicca et al., 2012). FAAH also hydrolyzes 2-AG although this degradation seems to be minor in vivo. Additionally, cyclooxygenase 2 (COX-2) has the ability to metabolize both AEA and 2-AG, generating arachidonic acid (Fowler, 2007). The transport of the eCBs through the plasma membrane is not fully understood. However, being lipophilic, they have the ability to cross the phospholipid bilayer by simple diffusion as well as through the action of the putative endocannabinoid membrane transporter (EMT) (Kaczocha et al., 2009). Furthermore, eCBs may enter the cell by endocytosis (Chicca et al., 2012). However, there remains considerable debate, as the EMT is yet to be characterized (Nicolussi and Gertsch, 2015). In addition to AEA, other ethanolamides including N-palmitoylethanolamine (PEA) (Di Marzo et al., 1996) and N-oleoylethanolamine (OEA) (Rodriguez de Fonseca et al., 2001) are collectively termed as N-acylethanolamines. More recently, other endocannabinoid-related lipids have been discovered, such as virodhamine (Porter et al., 2002) and 2-arachydonoylglycerol ether (Hanus et al., 2001), but their physiological relevance is not fully understood.
Figure 1. The elements of the endocannabinoid system and related receptors . The endocannabinoid system (ECS) is composed of its two main endocannabinoids AEA and 2-AG, which activate their primary targets CB1 and CB2 as well as their secondary targets GPR55, PPAR and TRPV1, among others. AEA is synthesised from cell membrane lipids by NAPE-PLD and is degraded primarily by FAAH. 2-AG is synthesised also from membrane lipid precursors by the DAGL enzyme and is degraded by MAGL and, also, to a minor extent, by ABHD6 and ABHD12. Besides their proposed ability to move to and from the cell by diffusion, the existence of an endocannabinoid specific transporter, the EMT, has been proposed that eases this bidirectional flow. 2-AG, 2-arachydonoylglycerol; ABHD6, α/β-hydrolase-6; ABHD12, α/β-hydrolase-12, AEA, anandamide; CB1, cannabinoid receptor 1; CB2, cannabinoid receptor 2; DAGL, diacylglycerol lipase; EMT, endocannabinoid membrane transporter; FAAH, fatty acid amide hydrolase; GPR55, G-protein-coupled receptor 55; GPR119, G-protein-coupled receptor 119; MAGL, monoacylglycerol lipase; NAPE-PLD, N-acylphosphatidylethanolamine-specific phospholipase D; PPAR, peroxisome proliferator-activated receptors; TRPV1, transient receptor potential vanilloid 1. This image contains some elements adapted from the Servier Medical Art Image Bank (Servier Laboratories (Aust) Pty Ltd) licensed under a Creative Commons Attribution 3.0 Unported Licence.
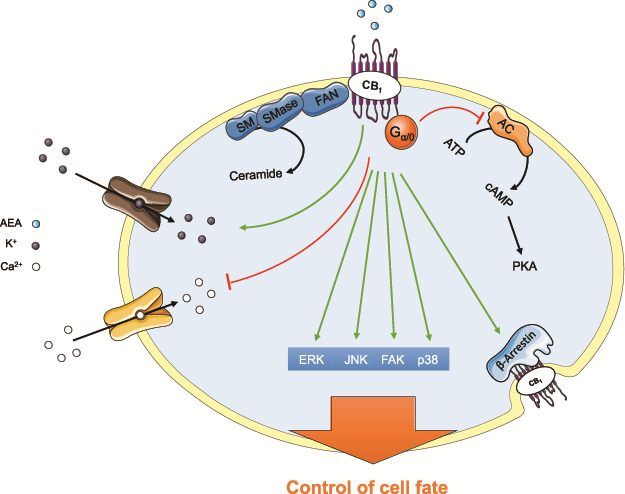
In general, activation of both cannabinoid receptors leads to inhibition of adenyl cyclase, resulting in a decrease in intracellular cAMP (Matsuda et al., 1990; Slipetz et al., 1995). In addition, downstream signalling cascades such as mitogen-activated protein kinase (MAPK) and extracellular signal–regulated kinases 1/2 are activated (Bouaboula et al., 1995; Ibsen et al., 2017). Moreover, β-arrestins, Ca2+ channels, inward rectifying K+ currents and phospholipase C are modulated by cannabinoid receptor activation (Zoratti et al., 2003). Different cannabinoid ligands may activate diverse pathways, leading to diverse responses, a concept termed ‘biased signalling’ (Ibsen et al., 2017). Furthermore, it has been demonstrated that cannabinoids, the natural or synthetic molecules able to bind the cannabinoid receptors, have the ability to modulate the sphingolipid-metabolizing pathways, by increasing ceramidase activity and leading to higher levels of ceramide, a second messenger that is able to control cell fate (Velasco et al., 2005) (Fig. 2). Besides the classic CB1 and CB2, other receptors can be activated by cannabinoids. Among them are the GPR55 (Ryberg et al., 2007), the transient receptor potential vanilloid 1 (TRPV1) (Huang et al., 2002), the peroxisome proliferator-activated receptors (PPAR) (Sun et al., 2006) and the GPR119 (Syed et al., 2012).
Figure 2. Cannabinoid receptor signalling. Cannabinoids exert their effects by binding to specific G-protein-coupled receptors. The major pathways and channels activated by CB1 receptor are depicted. These include inhibition of the adenylyl cyclase (AC)–cyclic AMP–protein kinase A (PKA) pathway; inhibition of voltage-operated Ca2+ channels and activation of K+ inwardly rectifying channels; activation of mitogen-activated protein kinase cascades (ERK; JNK, p38 and FAK); ceramide generation and β-arrestin recruitment. This image contains some elements adapted from the Servier Medical Art Image Bank (Servier Laboratories (Aust) Pty Ltd) licensed under a Creative Commons Attribution 3.0 Unported Licence.
The ECS is able to regulate cell proliferation, differentiation, migration and death. The outcome of these events is dictated by the molecular targets and cell type involved, affecting several physiological systems and their functions. In the central nervous system, the ECS modulates pain perception, motor functions and cognitive functions (Zou and Kumar, 2018). The ECS is also involved in the control of inflammation (Barrie and Manolios, 2017), energy balance through glucose homeostasis (Tibiriça, 2010), blood pressure and heart rate (Pacher et al., 2005), as well as a variety of functions in the reproductive system (Rossato et al., 2008; Walker et al., 2019). However, it was noticed that alterations in the cannabinoid receptors and eCBs metabolic enzyme expression, as well as in eCBs levels, were associated with diverse pathologies (Di Marzo, 2018). Taking these observations into account, therapies centred on the elements of the ECS started to be developed, mainly agonists/antagonists of CB1 and CB2 or inhibitors of the ECS metabolic enzymes (Di Marzo, 2008). However, eCBs interact with multiple receptors and many chemically related mediators also have the ability to activate these receptors. Therefore, the pharmacological manipulation of the ECS remains a challenge.
Several breakthroughs have been made recently on the association between lifestyle factors, such as dietary habits, light exposure and physical activity, and alterations of the ECS and endocannabinoidome (eCBome), which includes endocannabinoid-related mediators, their targets and metabolic enzymes. Increasing evidence is now linking the eCBome and the gut microbiome (Di Marzo and Silvestri, 2019). In the gut, it was found that the microbiota and the ECS have a bidirectional relationship, in which an increase in CB1 signalling, paired with obesity and related disorders, may be associated with impaired metabolism through dysbiosis (Di Marzo, 2018). Moreover, CB1 activation was also associated with intestinal dysbiosis caused by altered intestinal permeability (Karwad et al., 2017). In addition, it has been reported that the probiotic Lactobacillus acidophilus exerts antinociceptive effects against visceral pain through the increase of CB2 expression in the intestinal epithelial cells (Rousseaux et al., 2007).
In addition to the crosstalk between gut microbiota and ECS elements, dietary habits can also affect homeostasis of the ECS. Obesogenic diets, characterized by high fat content, were shown to lead to increased AEA and or 2-AG levels (Piscitelli et al., 2011). Moreover, this type of maternal diet has an impact on the infant eCBome (Miranda et al., 2018) and induces an increase in adiposity as well as a preference for high fat diets (Dias-Rocha et al., 2018). Furthermore, the eCBome was found to have a role as mediator of the positive effects of spicy food in the decrease of overall mortality by ischemic heart diseases, as well as in the reduction of diabetes. These effects appear to be mediated by the activation of TRPV1 and PPARα by the active compound of chili peppers, capsaicin (Kang et al., 2010). Sunlight also has an influence on the eCBome, since ultraviolet-B radiation (UVB) has the ability to upregulate CB1 expression and AEA, PEA and 2-AG levels in keratinocytes (Magina et al., 2011). In addition, physical exercise modulates eCBs levels, as AEA and or 2-AG are increased in active men when compared to sedentary ones (Raichlen et al., 2012; Raichlen et al., 2013). These are examples of the fast-growing roles of the endocannabinoids in several areas of human physiology and how they seem to modulate lipid metabolism by establishing a crosstalk with other lipid mediators.
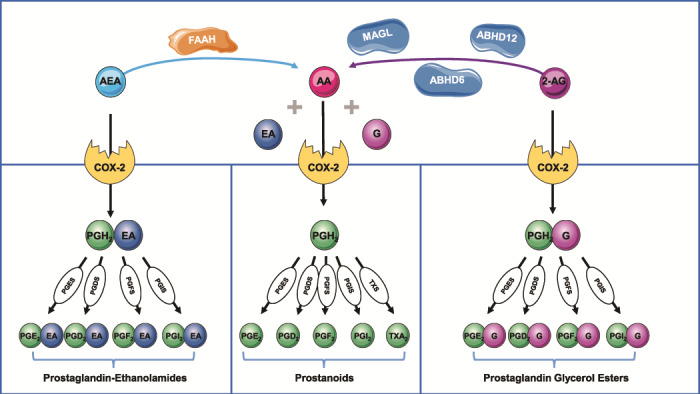
As addressed previously, eCBs are also oxidized by COX-2 and the resulting endoperoxides are converted by prostaglandin synthases to prostaglandin ethanolamides (prostamides) and prostaglandin glycerol esters (Fig. 3), highlighting the crosstalk between endocannabinoids and prostaglandin metabolic pathways (Rouzer and Marnett, 2011). Both of these families of lipid mediators are very important during several reproductive processes, such as development of the placenta, as well as in endometrium remodelling to receive the implanting blastocyst (Mitchell et al., 2016). It is now clear that the ECS is involved in important reproductive events from oocyte production to parturition. Its relevance has been demonstrated by the tight regulation of eCBs (Maccarrone, 2009) in order to fulfil their functions in the uterus (Guo et al., 2005; Wang et al., 2007; Fonseca et al., 2010a; Fonseca et al., 2010b; Scotchie et al., 2015) and placenta (Kenney et al., 1999; Helliwell et al., 2004; Park et al., 2003; Trabucco et al., 2009), where the presence of the ECS has been reported.
Figure 3. Crosstalk between the endocannabinoid and eicosanoid system pathways. FAAH is the main enzyme responsible for the hydrolysis of AEA into arachidonic acid (AA) and ethanolamine (EA). 2-AG is mainly degraded by MAGL and, to a smaller extent, by ABHD6 and ABHD12 into AA and glycerol (G). AA is metabolised by cyclooxygenase 2 (COX-2) leading to the formation of the endoperoxide PGH2. Tissue specific metabolism of PGH2 by PG synthases (PGS) yields the different prostanoids: prostaglandins (PGD2, PGE2 and PGF2α), prostacyclin (PGI2) and thromboxane A2 (TXA2). AEA and 2-AG are also metabolised by COX-2 to produce prostaglandins–ethanolamides (or prostamides) and prostaglandin glycerol esters, respectively. However, as yet the molecular targets of these lipid mediators have not been totally elucidated. This image contains some elements adapted from the Servier Medical Art Image Bank (Servier Laboratories (Aust) Pty Ltd) licensed under a Creative Commons Attribution 3.0 Unported Licence.
Given the involvement of these cannabinoids in fertility, reproduction and endocrine function (Correa et al., 2016; Walker et al., 2019), in this review we explore alterations of the ECS elements in the reproductive system and associated pathologies, their potential or lack thereof for pharmacological manipulation and how the altered levels of eCBs in these pathologies can be used to monitor disease severity or progression.
ECS in the Reproductive Tissues
The majority of ECS elements are expressed in human ovaries (El-Talatini et al., 2009), oviduct (Wang et al., 2006) and uterus (Das et al., 1995; Paria et al., 2001; Scotchie et al., 2015). In fact, the endometrium has the highest levels of N-acylethanolamines found in reproductive tissues (Das et al., 1995).
The cannabinoid receptors, alongside AEA and 2-AG metabolic enzymes, have also been characterized in uterine tissues throughout the menstrual cycle (Taylor et al., 2010; Scotchie et al., 2015). The expression of the metabolic enzymes and of the cannabinoid receptors was assessed by western blot, immunohistochemistry and real-time PCR, showing that these elements undergo changes according to the fluctuations in steroid hormones (Ribeiro et al., 2009; Scotchie et al., 2015; Maia et al., 2017). These variations consequently lead to changes in the eCB levels throughout the menstrual cycle, with an increase in AEA levels in the ovary upon ovulation (El-Talatini et al., 2009) and higher plasma levels in the follicular phase when compared to those in the luteal phase (Habayeb et al., 2004). Curiously, while in the ovarium high intrafollicular levels of AEA must be achieved, uterine and plasma levels must be low for the successful implantation of a fertilized oocyte (Maccarrone, 2009). Furthermore, CB1 expression is higher in the glandular epithelium than in stroma and is under the regulation of progesterone (Resuehr et al., 2012).
ECS in the endometrium
During the menstrual cycle, the endometrial stromal cells proliferate and differentiate giving rise to decidual cells in a process called decidualisation (Okada et al., 2018). This specialized tissue possesses the two main cannabinoid receptors and AEA metabolic enzymes (Habayeb et al., 2004; Almada et al., 2016a). The process of decidualisation can be disrupted by high levels of AEA, which inhibit endometrial proliferation through deregulation of the cell cycle, as well as by impaired differentiation by activation of CB1 receptor (Almada et al., 2016a). In addition, our group recently demonstrated that the negative impact of AEA in decidualisation seems to be caused by a disruption of oestrogen signalling due to the inhibition of aromatase, an enzyme responsible for a key step in the biosynthesis of oestrogens, and to a decrease in aromatase and oestrogen receptor transcripts (Almada et al., 2019). Furthermore, it was shown that WIN 55212-2, a CB1 agonist, decreases the transcription of specific decidualisation markers in human decidual cells, while a CB1 antagonist, AM251, induces the opposite effect (Moghadam et al., 2005). Moreover, AEA is important during decidualisation since AEA levels are lower in decidual cells when compared to non-differentiated cells (Almada et al., 2016b). In addition, miscarriage decidua and uterine natural killer cell-conditioned medium from miscarriage samples exhibit high AEA levels, which, in turn, interfere with the decidualisation process (Fonseca et al., 2019).
Rat models have provided evidence of the importance of CB1 in this process, due to its increased expression during the peak of decidua development (Fonseca et al., 2009). Additionally, AEA induced apoptosis of rat decidual cells through a mechanism that involves ceramide synthesis and p38 MAPK activation, compromising the decidualisation process (Fonseca et al., 2013). Moreover, in primary endometrial stromal cells isolated from pregnant rat uterus and stimulated for decidualisation, AEA treatment markedly diminished the differentiation programme and the administration of AEA in the uterine lumen of pseudopregnant rats prevented the decidualisation process (Fonseca et al., 2015). Several studies in rats have shown that throughout pregnancy there is an AEA gradient in the uterus. This gradient seems to be directly involved in the implantation process and embryo development (Paria et al., 1995; Guo et al., 2005), while promoting trophoblast differentiation and outgrowth (Das et al., 1995). Once the maximum development of decidua is reached, it undergoes regression by apoptosis (Gu et al., 1994; Dai et al., 2000). This phase is associated with the development of the placenta, which requires the genesis of different trophoblast types.
Globally, these studies and observations show the importance of a tight control of eCB levels by the enzymes of synthesis and degradation in order to achieve a successful decidualisation process and blastocyst implantation.
ECS in the placenta
The importance of AEA levels in endometrial turnover and the implications for the success of implantation is well described, but the role of the ECS during gestation, namely in placentation, is still obscure. Throughout pregnancy, plasma AEA levels decrease from the first to the second and third trimester, with no changes between the second and third trimesters. At term, plasma AEA levels dramatically increase during labour (Habayeb et al., 2004). In addition to these evident fluctuations in plasma AEA, local endocannabinoid signalling may also modulate critical processes of cellular turnover during placental development.
Proliferation and differentiation of the trophoblasts is required for normal placentation and physiological functions. Among the most important trophoblast cell types are cytotrophoblast (CT) and syncytiotrophoblast (ST). The CT proliferates and differentiates into extravillous trophoblasts and ST. ST are responsible for the endocrine function and constitute the physical barrier between maternal and foetal blood, where nutrient and gas exchange take place (Kidima, 2015). Therefore, there is a need for tight regulation between proliferation, differentiation and apoptosis among these cells for normal placental development to occur (Hemberger et al., 2019; Knöfler et al., 2019). The cannabinoid receptors, the main eCBs and their metabolic enzymes have been reported in the CT and ST of the first trimester and term placenta and in the trophoblastic type BeWo cell line (Kenney et al., 1999; Park et al., 2003; Helliwell et al., 2004; Habayeb et al., 2008; Trabucco et al., 2009; Marczylo et al., 2010). However, 2-AG levels were only measured in baboon placenta (Brocato et al., 2013) (Table I). In rat placentation, the ECS has been found as early as the 10th day of gestation in the trophoblastic cells of the ectoplacental cone and on the 14th day on the spongiotrophoblast cells (Sun et al., 2010). Not only is the ECS present in placental tissues, but also its importance in placental development has been demonstrated in several studies with knockout mouse models. In CB1−/− mice, trophoblast cells present reduced proliferation and the placenta has a lower weight when compared with wild-type mice (Sun et al., 2010). A lower trophoblast proliferation was also observed in FAAH−/− animals, which suggests the involvement of ECS in trophoblast proliferation and differentiation (Sun and Dey, 2012).
Table I. Endocannabinoid system elements described in the uterus and placenta.
| ECS element | Model | Study |
|---|---|---|
| AEA | Term placenta | (Marczylo et al., 2010) |
| 2-AG | Baboon placenta | (Brocato et al., 2013) |
| CB1 | Uterus | (Taylor et al., 2010) |
| Term placenta | (Kenney et al., 1999) | |
| Bewo cells | (Kenney et al., 1999) | |
| 1st Trimester placenta | (Helliwell et al., 2004) | |
| Amniotic epithelial cells | (Park et al., 2003) | |
| CB2 | Uterus | (Taylor et al., 2010) |
| Term placenta | (Kenney et al., 1999) | |
| BeWo cells | (Kenney et al., 1999) | |
| 1st Trimester placenta | (Helliwell et al., 2004) | |
| NAPE-PLD | Uterus | (Taylor et al., 2010) |
| Term placenta | (Aban et al., 2013) | |
| 1st Trimester placenta | (Trabucco et al., 2009) | |
| FAAH | Uterus | (Taylor et al., 2010) |
| Term placenta | (Park et al., 2003) | |
| 1st Trimester placenta | (Helliwell et al., 2004) | |
| Amniotic cells | (Park et al., 2003) | |
| Decidua | (Park et al., 2003) | |
| DAGLα | BeWo cells | (Costa et al., 2014) |
| Term placenta | (Costa et al., 2014) | |
| MAGL | BeWo cells | (Costa et al., 2014) |
| Term placenta | (Costa et al., 2014) |
2-AG, 2-arachydonoylglycerol; AEA, anandamide; CB1, cannabinoid receptor 1; CB2, cannabinoid receptor 2; DAGL, diacylglycerol lipase; FAAH, fatty acid amide hydrolase; MAGL, monoacylglycerol lipase; NAPE-PLD, N-acylphosphatidylethanolamine-specific phospholipase D.
In cell line models, it has been demonstrated that AEA prevents BeWo cell proliferation (Habayeb et al., 2008) and induces apoptosis (Costa et al., 2015a) and that 2-AG also causes cell death by a CB2-dependent mechanism (Costa et al., 2014). In addition, high levels of both AEA and 2-AG impair the synthesis of ST-related proteins as well as the endocrine function of primary ST (Costa et al., 2015a; Costa et al., 2015b). Once again, this highlights the role of the eCBs, this time in trophoblast turnover, reinforcing the importance of cannabinoid signalling in the fundamental cellular processes for placentation. Over the past few years, growing evidence proposes the ECS as a part of the relevant mechanisms that regulate the complexity of events that occur in the placenta. Nevertheless, further studies are needed to investigate the role of the ECS in the occurrence of placental-mediated complications in pregnancy.
ECS in Placental and Endometrial Disorders
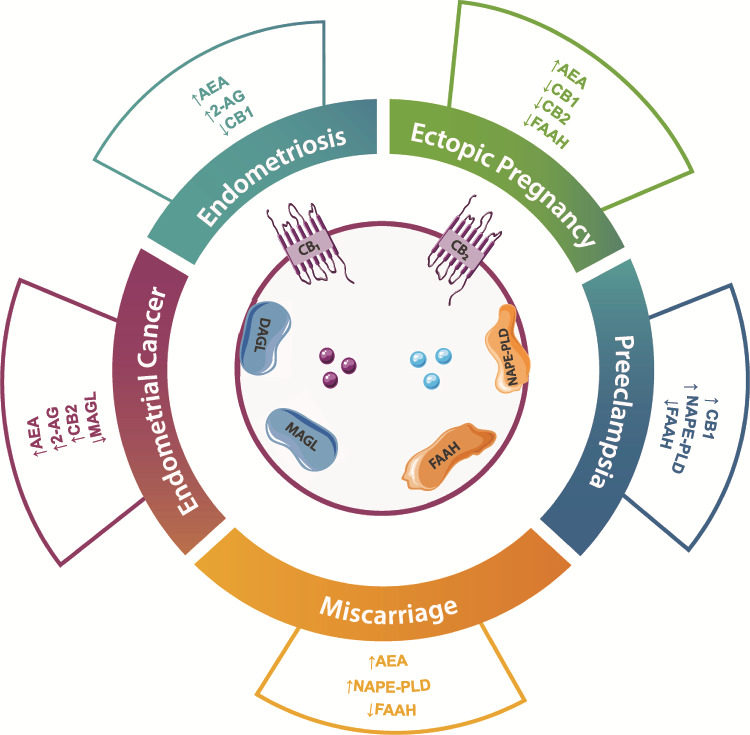
The presence of the ECS in the female reproductive tissues and its strict regulation suggest that changes in the homeostasis of its key components may be associated with several pathologies such as endometriosis, miscarriage, ectopic pregnancy, pre-eclampsia and endometrial cancer (Table II) (Fig. 4).
Table II. Summary of main findings regarding the alterations in the endocannabinoid system elements in endometrial and placental diseases.
| Pathology | Altered ECS | Tissue | Study |
|---|---|---|---|
| Endometriosis | AEA | ↑Plasma | (Sanchez et al., 2016) |
| 2-AG | ↑Plasma | (Sanchez et al., 2016) | |
| CB1 | ↓Endometrium | (Sanchez et al., 2016) | |
| ↑Myometrium | (Shen et al., 2019) | ||
| CB2 | ↑Myometrium | (Shen et al., 2019) | |
| Ectopic pregnancy | AEA | ↑Fallopian tubes | (Horne et al., 2008) |
| ↑Plasma | (Gebeh et al., 2013) | ||
| OEA | ↑Plasma | (Gebeh et al., 2013) | |
| PEA | ↑Plasma | (Gebeh et al., 2013) | |
| CB1 | ↓Decidua and Fallopian tubes | (Horne et al., 2008) | |
| FAAH | ↓Fallopian tubes | (Gebeh et al., 2012) | |
| ↓Plasma | (Gebeh et al., 2013) | ||
| Miscarriage | AEA | ↑Plasma | (Maccarrone et al., 2002) |
| ↑Uterine NK cells | (Fonseca et al., 2019) | ||
| CB1 | ↑1st Trimester placenta | (Trabucco et al., 2009) | |
| NAPE-PLD | ↑1st Trimester placenta | (Trabucco et al., 2009) | |
| FAAH | ↓Peripheral lymphocytes | (Maccarrone et al., 2000) | |
| ↓1st Trimester placenta | (Maccarrone et al., 2002) | ||
| ↓1st Trimester placenta | (Trabucco et al., 2009) | ||
| Pre-eclampsia | AEA | ↓Plasma | (Molvarec et al., 2015) |
| CB1 | ↑Placenta | (Fugedi et al., 2014) | |
| NAPE-PLD | ↑Placenta | (Fugedi et al., 2014) | |
| ↓Placenta | (Aban et al., 2013) | ||
| FAAH | ↓Placenta | (Fugedi et al., 2014) | |
| ↑Placenta | (Aban et al., 2013) | ||
| Endometrial cancer | AEA | ↑Endometrium | (Ayakannu et al., 2019) |
| ↑Endometrium | (Schmid et al., 2002) | ||
| ↑Plasma | (Ayakannu et al., 2014) | ||
| 2-AG | ↑Endometrium | (Guida et al., 2010) | |
| CB1 | ↓Endometrium | (Ayakannu et al., 2014) | |
| CB2 | ↑Endometrium | (Guida et al., 2010) | |
| ↓Endometrium | (Ayakannu et al., 2014) | ||
| MAGL | ↓Endometrium | (Guida et al., 2010) |
NK, natural killer; OEA, N-oleoylethanolamine; PEA, N-palmitoylethanolamine.
Figure 4. Alterations in the ECS elements in endometrial and placental diseases. Main findings regarding alterations in members of the ECS in endometriosis, miscarriage, ectopic pregnancy, pre-eclampsia and endometrial cancer. This image contains some elements adapted from the Servier Medical Art Image Bank (Servier Laboratories (Aust) Pty Ltd) licensed under a Creative Commons Attribution 3.0 Unported Licence.
Endometriosis and adenomyosis
Endometriosis is defined by the presence of endometrial glands or stroma at ectopic sites, such as the peritoneum and ovary, causing a chronic inflammatory reaction (Farquhar, 2007). Among the subtypes of endometriosis, there is one characterized by the migration of endometrial cells into the myometrium, named adenomyosis.
Endometriosis is one of the most common gynaecological disorders, with a prevalence of 2–10% among women worldwide (Horton et al., 2019). However, the prevalence may be higher in women of reproductive age, causing infertility (Meuleman et al., 2009). The great majority of patients present with pain, which is one of the major clinical features and is often associated with psychological distress and fatigue (Gibbons, 2004). In addition, in more than 95% of the cases, patients with deep infiltrating endometriosis are prone to severe pain (Bouaziz et al., 2017). There is currently no cure, and treatments are mainly intended to reduce endometriosis-associated pain and improve fertility (Dunselman et al., 2014).
Alterations in the ECS have been reported in endometriosis; however, little is known about its potential role in disease establishment and progression. Nevertheless, this is a topic that deserves further investigation since endocannabinoids modulate inflammation, cell proliferation and cell survival, as well as cell migration, all of which are processes that are critical to endometriosis (Sanchez et al., 2012). When it comes to cell migration, this condition is characterized by a higher endometrial expression of matrix metalloproteinase-2 (MMP-2) and lower expression of tissue inhibitor of MMP (TIMP)-2 and TIMP-3, compared with the endometrium of women without endometriosis (Chung et al., 2002). It is known that in cancer cells the cannabinoids have the ability to reduce their migratory effects, as shown by the inhibition of metastatic nodule formation in a Lewis lung carcinoma model (Portella et al., 2003), and block the migration of human colon carcinoma cells SW480 through a CB1 receptor-dependent mechanism (Joseph et al., 2004; Laezza et al., 2008). Moreover, the migration of endometrial cells is dependent on CB1 receptor activation (Gentilini et al., 2010).
Inflammation is one of the hallmarks of endometriosis. In experiments performed in mouse models of endometriosis, a specific subset of macrophages in the peritoneal fluid and macrophages in the ectopic lesions are involved in the angiogenic process (Capobianco et al., 2011). Moreover, several cytokines and growth factors are increased in the peripheral blood and peritoneal fluid of patients with endometriosis (Pizzo et al., 2002). It is well described that the CB1 and CB2 receptors are expressed in human and mouse immune cells, CB2 being present at higher levels (Parolaro, 1999; Lee et al., 2001), though the cannabinoids have the ability to modulate the immune response by mechanisms that are dependent or independent of CB signalling. In fact, the phytocannabinoid THC has the ability to reduce the chemotactic response of peritoneal macrophages to early inflammatory response (Raborn et al., 2008), while Rimonabant, a CB1 receptor inverse agonist, also inhibits the inflammatory response in human umbilical vein endothelial cells (Huang et al., 2010).
Given the importance of the ECS in the cellular processes that characterize endometriosis, we reviewed the recent findings regarding an association between this condition and alterations in the ECS elements. Using a rat model of endometriosis, Dmitrieva et al. (2010) demonstrated that CB1 receptor is expressed in the somata and fibres of the sensory and sympathetic neurons that innervate endometriotic lesions. This same study showed that CB1 agonists decrease endometriosis associated hyperalgesia, whereas antagonists increase it (Dmitrieva et al., 2010). Moreover, in women with endometriosis, there occurs a significant increase in plasmatic AEA and 2-AG levels together with a decrease in CB1 expression in endometrial biopsies, when compared with a control group in the secretory phase. This suggests a tissue downregulation of the cannabinoid signalling (Table II) (Sanchez et al., 2016). A similar conclusion regarding the decrease in CB1 receptor was also obtained in a study with human endometrial biopsies but this time in all phases of the cycle, not only in the secretory phase (Resuehr et al., 2012). On the other hand, CB1 and CB2 mRNA and protein levels are significantly higher in junctional zone and outer myometrium tissue samples from patients with adenomyosis than from women without adenomyosis. Also, CB1 levels are directly correlated with dysmenorrhea severity (Shen et al., 2019). Nonetheless, the involvement of the ECS in this condition may also be a way to relieve pain, since deep-infiltrating endometriotic nodules implanted in nude mice treated with the synthetic cannabinoid WIN 55212-2, a CB1 agonist, led to a reduction in nodule size. In addition, WIN 55212-2 also exerted in vitro antiproliferative effects on human endometriotic cells, coupled with an inhibition of the AKT (or protein kinase B) pathway (Leconte et al., 2010). In a mouse model of endometriosis, methanandamide increased the lesion volume, as well as the mRNA levels of survivin, N-cadherin, integrin β1 and interleukin-6, molecules that are involved in cell adhesion and inflammation (Sanchez et al., 2017). Moreover, in this study, by using CB1−/− mice, the authors demonstrated that the lack of CB1 expression in the recipient peritoneal environment leads to an impairment in lesion volume and lower survivin and N-cadherin expression levels (Sanchez et al., 2017). Although the observed changes in ECS indicate the involvement of cannabinoid receptors and of the major eCB, AEA, in endometriosis, little is known about the expression of the metabolic enzymes and their potential contribution to the eCB fluctuation.
Despite the fact that some of these reports were exploratory, their findings on the alteration of ECS elements and cannabinoid signalling in endometriosis provides momentum for further investigation of the pharmacotherapy for this disease (Pertwee, 2009). Nevertheless, several factors need to be taken into account, including the time and tissue-specific availability of mediators involved and their modulation by endogenous factors (such as hormones), inflammatory factors and environmental stimuli (Bisogno & Maccarrone, 2014). However, two clinical trials in patients with deep endometriosis-induced hyperalgesia that include cannabinoids have been ongoing for some time. The oral formulation of THC/CBD (1:1) (clinical trial NCT03875261) and the combination of PEA and transpolydatin (clinical trial NCT02372903) are being tested, although the results have not yet been published.
Given the relevance of pain in this pathology, the use of phytocannabinoids as in the clinical trial to ease the pain seems to be a good strategy. However, given the existence of cannabinoid receptors throughout the body, this approach is not very specific. Since the ECS is closely related to pain modulation, future strategies could involve the downregulation and antagonism of the cannabinoid receptors and/or TRPV1, or to modulate their actions locally.
Ectopic pregnancy
Ectopic pregnancy occurs as a result of an embryo implanting outside the uterus, most commonly in the Fallopian tube, with a prevalence of 1–2% of all pregnancies (Shaw et al., 2010). The characteristics of this pathology are abnormal bleeding, pain and rupture of the fallopian tube, which accounts for the leading cause of pregnancy-related first trimester maternal death in Europe and the USA (Farquhar, 2005; Varma & Gupta, 2009).
Given the risk of mortality and morbidity associated with the lack of a reliable method of diagnosis, knowledge about the molecular mechanisms that lead to tubular implantation is needed. Alterations in the ECS elements have been linked with ectopic pregnancy. Studies in mice have shown that pharmacological and genetic silencing of the CB1 receptor leads to retention of embryos in the oviduct and, consequently, implantation failure (Wang et al., 2004). A similar effect was reported when methanandamide was administered. In humans, the fallopian tube and decidua of women with ectopic pregnancy express less CB1 when compared to healthy women (Horne et al., 2008). In addition to the downregulation of the receptors, it has been reported that the fallopian AEA levels in these women are higher than in healthy non-pregnant women, which seems to be related to a reduced FAAH expression in the fallopian tube epithelium (Table II) (Gebeh et al., 2012). A similar increase in AEA levels, as well as in OEA and PEA, were also reported in plasma, this effect being mainly due to a decrease of FAAH levels in the peripheral blood cells (Gebeh et al., 2013). Therefore, the local and plasma levels seem to be directly correlated with the downregulation or reduced activity of local or peripheral FAAH.
Nonetheless, these studies suggest that the high eCB levels observed in women with an ectopic pregnancy may have a functional impact in modulating fallopian tube function. However, even though local alterations in AEA levels are reflected in the plasma, given the variety of physiological functions in which the AEA is involved, it is not easy to use this eCB as an early predictor of ectopic pregnancy. Therefore, AEA would always have to be associated with other molecules related to the condition. Further studies are required to evaluate the complementary mechanisms and pathways involved in how eCBs modulate cilia movement and/or tubal smooth muscle contractility.
Miscarriage and preterm birth
The correlation between cannabinoids and adverse effects on pregnancy has been demonstrated through several epidemiological studies. Among the described negative effects, there are early foetus loss, preterm birth and low birthweight (Fried, 1995; El Marroun et al., 2009). The importance of a proper AEA tonus for embryo implantation has been described, as deficiencies in FAAH levels lead to impaired implantation and, consequently, infertility (Wang et al., 2006). These observations suggest that possible alterations in AEA levels may cause miscarriage and implantation failure. In fact, a study that compared FAAH with CB1 protein expression and NAPE-PLD transcription in first trimester placenta of spontaneous miscarriage and voluntary pregnancy termination reported a decrease in FAAH protein expression and an increase in both NAPE-PLD transcripts and CB1 protein in women of the spontaneous miscarriage group when compared to the voluntary pregnancy termination group (Trabucco et al., 2009). Moreover, a decrease in FAAH was found in peripheral lymphocytes of women who miscarried in comparison with pregnant women at 6–11 weeks of gestation (Maccarrone et al., 2000) and in the endometrium of women with unexplained infertility (Cui et al., 2017). A similar study also observed that the same decrease in FAAH activity and protein level, resulting in high AEA levels, is related to failure in IVF embryo transfer and spontaneous miscarriage (Table II) (Maccarrone et al., 2002). Furthermore, the rise in plasma AEA levels observed in non-viable first-trimester pregnancies seems to be correlated with pregnancy-related hormones such as pregnancy-associated plasma protein A (PAPP-A) (Tayloret al., 2011). Contrary to the decrease in FAAH observed in most of these reports, in the trophoblasts that invaded the decidua from patients who experienced multiple miscarriages, this enzyme is in fact overexpressed and was found in the nucleus of these cells in 67% of the women when compared to those who had early pregnancy termination (Chamley et al., 2008).
These reports undoubtedly indicate that in order to allow pregnancy progression a fine regulation of AEA levels in the placental environment is necessary. In addition, low FAAH and CB2 levels and high CB1 and NAPE-PLD expression characterize peripheral blood cells and/or placental tissues from spontaneous miscarriage. Since progesterone and estradiol have the ability to activate the FAAH promoter (Maccarrone et al., 2003; Grimaldi et al., 2012), more research on the molecular events leading to placental FAAH regulation should be carried out. Moreover, given the inhibition of choriocarcinoma growth in vitro by AEA via CB2 (Habayeb et al., 2008), additional mechanisms related to CB2 activity have to be taken into account.
Preterm birth is the leading cause of perinatal morbidity and mortality worldwide. As previously referred to, plasma AEA levels increase dramatically during labour (Habayeb et al., 2008). Also as mentioned above, COX-2 has the ability to oxidize the eCBs. In addition, the degradation product of AEA and 2-AG, arachidonic acid, is also a substrate for the synthesis of prostaglandins, showing an important crosstalk between eCBs and prostaglandins (Alhouayek and Muccioli, 2014). Given the implications of AEA and other lipid mediators, such as prostaglandins, in uterine contractions, by using a mouse model of preterm labour, the downregulation of CB2 as well as an upregulation of CB1 and NAPE-PLD accompanied by an increase in prostaglandin F2α has been identified (Bariani et al., 2015).
The first study on a single blood test of AEA levels as a predictor of preterm labour has recently been published (Bachkangi et al., 2019). Both AEA and PEA levels predicted the gestational age of delivery and remaining period of pregnancy with better accuracy than conventional tests. Although further studies need to be performed for both PEA and OEA (since the number of women tested was significantly lower when compared to the group tested for AEA), this approach is remarkable, since it uses eCB for the first time as biomarker for reproductive pathology in a single blood test.
Pre-eclampsia
Pre-eclampsia is a pregnancy-specific disorder characterized by the development of hypertension and proteinuria after Week 20 of gestation in previously normotensive women. This condition has a prevalence of 3–8% worldwide (Abalos et al., 2013) and is among the leading causes of maternal and perinatal mortality and morbidity. The main causes of this condition seem to be an excess of maternal systemic inflammatory response to pregnancy and a systemic oxidative stress, as well as an impairment of circulating angiogenic and antiangiogenic factors (Redman and Sargent, 2004; Aouache et al., 2018; Helmo et al.,2018). The diagnosis of this condition is still very ineffective. Some women with pre-eclampsia report an absence of hypertension and proteinuria, indicating that adverse events occur in the mother even when the standard clinical definition of pre-eclampsia is yet to be met (Kent, 2008).
Unrecognized foetal compromise contributes to the rate of foetal demise, and 1 in 20 stillbirths without congenital abnormality is complicated by, or attributable to pre-eclampsia (Heslehurst et al., 2010). Recent works have demonstrated that changes in the ECS expression pattern are associated with this condition. Firstly, women with pre-eclampsia exhibit reduced levels of AEA (Molvarec et al., 2015). However, this reduction was not correlated with the currently used standard pre-eclamptic markers, soluble Fms-like tyrosine kinase-1 and phosphatidylinositol-glycan biosynthesis class F protein. When looking into other ECS members, CB1 receptor levels are higher in pre-eclamptic placental tissues, though no changes have been reported at the CB2 and FAAH levels (Fugedi et al., 2014). These impairments are not consensual, since a previous study verified no changes in CB1 protein levels, but instead an increase in NAPE-PLD and a decrease in FAAH in pre-eclamptic placentas when compared to placentas of normotensive subjects (Aban et al., 2013) (Table II). Moreover, a study regarding the association between polymorphisms in the CB1 gene (CNR1) and pre-eclampsia in the population of central Europe found that the single nucleotide polymorphism rs806368 in this gene is correlated with this condition. This suggests a potential role of this polymorphism as a susceptibility marker for pre-eclampsia (Bienertova-Vasku et al., 2011).
The described reports have shown that serum AEA concentrations are lower in women with pre-eclampsia. However, it is also possible that changes in the ECS elements are a consequence rather than a cause of pre-eclampsia; nevertheless, there is an association between placental ECS expression levels and pre-eclampsia risk factors or pathophysiological signals. The study of this pathology in the early stages is still a challenge. Moreover, given the lack of correlation of lower concentrations of AEA with other known markers of pre-eclampsia, other lipids such as 2-AG, OEA or PEA should be evaluated in order to potentially provide a better association with this pathology.
Endometrial cancer
Endometrial cancer is the most prevalent gynaecological cancer in developed countries (Torre et al., 2015). The number of cases is increasing mainly due to a variety of environmental and lifestyle factors, life span and obesity being among them (Amant et al., 2005), as well as genetic mutations (Colombo et al., 2013). This cancer is classified as either type I oestrogen-dependent or type II oestrogen-independent. Type I tumours are mainly low grade, oestrogen-dependent, hormone-receptor-positive adenocarcinomas with endometrioid morphology and are often referred to as endometrioid endometrial cancers (Buhtoiarova et al., 2016; Morice et al., 2016). Type II ECs are generally high grade, hormone-receptor negative and have poor survival rates (Morice et al., 2016). Therefore, the latter requires a more aggressive treatment (Colombo et al., 2013).
Cannabinoids have the ability to regulate or inhibit certain cellular processes essential for cancer progression and development. It has been shown that AEA (Laezza et al., 2006) and THC (Caffarel et al., 2006) arrest the cell cycle progression in breast cancer cells, being THC effects dependent on CB2 activation. In addition, cannabinoids can either inhibit cancer cell proliferation or induce apoptosis, as reviewed by Ayakannu et al. (2015). Moreover, the capacity of cannabinoids to inhibit cancer angiogenesis was shown by a decrease in vascular endothelial growth factor (VEGF) and VEGF receptor-1 (R-I) in thyroid cancer (Portella et al., 2003), as well as VEGF and VEGFR-2 expression in patients with glioblastoma (Blazquez et al., 2004).
In this endometrial disease, it has been reported that AEA has the ability to induce the migration of endometrial cancer cell line HEC-1B by a mechanism independent of CB1 (McHugh et al., 2012). This observation remains controversial, as it has been described as dependent on this same receptor for migration following Met-AEA treatment (Gentilini et al., 2010). Moreover, in human endometrial carcinoma biopsies, CB2 expression and 2-AG levels were higher and MAGL expression was lower, when compared to normal endometrium (Guida et al., 2010). This same increase in AEA was verified in endometrial carcinoma biopsies (Schmid et al., 2002). Again, these results are not consensual, since another study using biopsies of patients with both types of endometrial cancer reported a decrease in CB2 and CB1, accompanied by an increase in plasma AEA (Ayakannu et al., 2014; Ayakannu et al., 2019). These data show that although changes in AEA levels as well as in CB1 and CB2 expression seem to occur, there is no definitive correlation between these alterations and endometrial cancer (Table II).
Overall, the literature addressing this issue suggests that downregulation of CB1 and CB2 receptor transcript levels and higher tissue and plasma AEA concentrations are correlated with endometrial cancer. This allows speculation about the role of ECS in the aetiology of this cancer, although further research is warranted. In addition, the correlation between plasma and tissue AEA concentrations suggests that AEA could be used as a predictor for some forms of endometrial malignant disease, especially type 1 endometrial adenocarcinoma. Another strategy may be the search in endometrial biopsies for alterations in CB2 expression since these studies have found a downregulation of this receptor in endometrial cancer. Thus, the ECS presents an attractive target for pharmacological intervention in some endometrial malignancies.
Discussion
The study of the ECS in human reproduction started nearly 20 years ago. Nowadays, the importance of a tight regulation of eCB levels throughout the menstrual cycle, decidualisation and placenta development is well known. Alterations in ECS homeostasis can lead to abnormal modulation of fundamental cellular processes involved in reproductive pathologies, such as pre-eclampsia, ectopic pregnancy and endometriosis. However, despite an apparent link between alterations in AEA levels and most of these pathologies, it is difficult to use AEA as a biomarker since it is highly unspecific, an exception being made for its successful use as a predictor of preterm birth. Yet, even in this case, the combination of AEA with other lipid mediators, such as OEA or PEA, could improve even further the accuracy of this test. As in this case, in other pathologies, the use of AEA as a biomarker for diagnosis would also benefit from the association with other molecules. Moreover, alterations in the expression of CB1 and CB2 in biopsies could be used to predict alterations that seem to be characteristic of some pathologies such as endometrial cancer.
The great challenge of these observations remains whether or not the alterations in the elements of the ECS are the cause or the consequence of the genesis of some of these pathologies. Still, much has yet to be unveiled and concluded about the role of the ECS since 2-AG and its metabolic enzymes, as well as other promising endocannabinoid-related lipids such as PEA or OEA, were not included in these studies.
Patients could potentially benefit from therapies developed to correct alterations in the ECS and the ongoing clinical studies will contribute to the understanding of how this complex system could be pharmacologically manipulated in these pathologies.
Acknowledgements
Some parts of the images used in this article were adapted from the Servier Medical Art Image Bank (Servier Laboratories (Aust) Pty Ltd). The authors also thank Joana Macedo for helping with image design.
Authors’ roles
J.M. collected the information, designed the figures and wrote the manuscript. B.M.F, N.T. and G.C.-S. critically revised the manuscript. All the authors have seen and approved the final version.
Funding
FEDER Funds through the Operational Competitiveness Factors Program (COMPETE); National Funds through FCT (Foundation for Science and Technology) within the scope of the project ‘PTDC/DTP-FTO/5651/2014—POCI-01-0145-FEDER-016562’; FCT (PhD grant BD/136105/2018 to J.M.).
Conflict of interest
The authors declare no conflict of interest.
References
- Abalos E, Cuesta C, Grosso AL, Chou D, Say L. Global and regional estimates of preeclampsia and eclampsia: a systematic review. Eur J Obstet Gynecol Reprod Biol 2013;170:1–7. [DOI] [PubMed] [Google Scholar]
- Aban C, Leguizamon GF, Cella M, Damiano A, Franchi AM, Farina MG. Differential expression of endocannabinoid system in normal and preeclamptic placentas: effects on nitric oxide synthesis. Placenta 2013;34:67–74. [DOI] [PubMed] [Google Scholar]
- Alger BE, Kim J. Supply and demand for endocannabinoids. Trends Neurosci 2011;34:304–315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alhouayek M, Muccioli GG. COX-2-derived endocannabinoid metabolites as novel inflammatory mediators. Trends Pharmacol Sci 2014;35:284–292. [DOI] [PubMed] [Google Scholar]
- Almada M, Amaral C, Diniz-da-Costa M, Correia-da-Silva G, Teixeira NA, Fonseca BM. The endocannabinoid anandamide impairs in vitro decidualization of human cells. Reproduction 2016a;152:351–361. [DOI] [PubMed] [Google Scholar]
- Almada, M, Cunha, S, Fonseca, BM, Amaral, C, Piscitelli, F, Di Marzo, V, Correia-da-Silva, G, Teixeira, N. Anandamide interferes with human endometrial stromal-derived cell differentiation: an effect dependent on inhibition of cyclooxygenase-2 expression and prostaglandin E2 release. Biofactors 2016b;42:277–286. [DOI] [PubMed] [Google Scholar]
- Almada M, Oliveira A, Amaral C, Fernandes PA, Ramos MJ, Fonseca B, Correia-da-Silva G, Teixeira N. Anandamide targets aromatase: a breakthrough on human decidualization. Biochim Biophys Acta Mol Cell Biol Lipids 2019;158512. [DOI] [PubMed] [Google Scholar]
- Amant F, Moerman P, Neven P, Timmerman D, Van Limbergen E, Vergote I. Endometrial cancer. Lancet 2005;366:491–505. [DOI] [PubMed] [Google Scholar]
- Aouache R, Biquard L, Vaiman D, Miralles F. Oxidative stress in preeclampsia and placental diseases. Int J Mol Sci 2018;19:1496. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ayakannu T, Taylor AH, Marczylo TH, Maccarrone M, Konje JC. Identification of novel predictive biomarkers for endometrial malignancies: N-Acylethanolamines. Front Oncol 2019;9:430. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ayakannu T, Taylor AH, Marczylo TH, Willets JM, Brown L, Davies Q, Moss E, Konje JC. Association of cannabinoid receptor expression with anandamide concentrations in endometrial cancer. The Lancet 2014;383:S23. [Google Scholar]
- Ayakannu T, Taylor AH, Willets JM, Konje JC. The evolving role of the endocannabinoid system in gynaecological cancer. Hum Reprod Update 2015;21:517–535. [DOI] [PubMed] [Google Scholar]
- Bachkangi P, Taylor AH, Bari M, Maccarrone M, Konje JC. Prediction of preterm labour from a single blood test: the role of the endocannabinoid system in predicting preterm birth in high-risk women. Eur J Obstet Gynecol Reprod Biol 2019;243:1–6. [DOI] [PubMed] [Google Scholar]
- Bariani MV, Dominguez Rubio AP, Cella M, Burdet J, Franchi AM, Aisemberg J. Role of the endocannabinoid system in the mechanisms involved in the LPS-induced preterm labor. Reproduction 2015;150:463–472. [DOI] [PubMed] [Google Scholar]
- Barrie N, Manolios N. The endocannabinoid system in pain and inflammation: its relevance to rheumatic disease. Eur J Rheumatol 2017;4:210–218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bienertova-Vasku J, Bienert P, Dostalova Z, Chovanec J, Vasku A, Vasku V. A common variation in the cannabinoid 1 receptor (CNR1) gene is associated with pre-eclampsia in the central European population. Eur J Obstet Gynecol Reprod Biol 2011;155:19–22. [DOI] [PubMed] [Google Scholar]
- Bisogno T, Maccarrone M. Endocannabinoid signaling and its regulation by nutrients. Biofactors 2014;40:373–380. [DOI] [PubMed] [Google Scholar]
- Blankman JL, Simon GM, Cravatt BF. A comprehensive profile of brain enzymes that hydrolyze the endocannabinoid 2-arachidonoylglycerol. Chem Biol 2007;14:1347–1356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blazquez C, Gonzalez-Feria L, Alvarez L, Haro A, Casanova ML, Guzman M. Cannabinoids inhibit the vascular endothelial growth factor pathway in gliomas. Cancer Res 2004;64:5617–5623. [DOI] [PubMed] [Google Scholar]
- Bouaboula M, Poinot-Chazel C, Bourrié B, Canat X, Calandra B, Rinaldi-Carmona M, Le Fur G, Casellas P. Activation of mitogen-activated protein kinases by stimulation of the central cannabinoid receptor CB1. Biochem J 1995;312(Pt 2):637–641. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bouaziz J, Bar On A, Seidman DS, Soriano D. The clinical significance of endocannabinoids in endometriosis pain management. Cannabis Cannabinoid Res 2017;2:72–80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brocato B, Zoerner AA, Janjetovic Z, Skobowiat C, Gupta S, Moore BM, Slominski A, Zhang J, Schenone M, Phinehas R et al. Endocannabinoid crosstalk between placenta and maternal fat in a baboon model (Papio spp.) of obesity. Placenta 2013;34:983–989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buhtoiarova TN, Brenner CA, Singh M. Endometrial carcinoma: role of current and emerging biomarkers in resolving persistent clinical dilemmas. Am J Clin Pathol 2016;145:8–21. [DOI] [PubMed] [Google Scholar]
- Caffarel MM, Sarrio D, Palacios J, Guzman M, Sanchez C. Delta9-tetrahydrocannabinol inhibits cell cycle progression in human breast cancer cells through Cdc2 regulation. Cancer Res 2006;66:6615–6621. [DOI] [PubMed] [Google Scholar]
- Capobianco, A, Monno, A, Cottone, L, Venneri, MA, Biziato, D, Di Puppo, F, Ferrari, S, De Palma, M, Manfredi, AA, and Rovere-Querini, P. Proangiogenic Tie2(+) macrophages infiltrate human and murine endometriotic lesions and dictate their growth in a mouse model of the disease. Am J Pathol 2011;179:2651–2659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chamley LW, Bhalla A, Stone PR, Liddell H, O'Carroll S, Kearn C, Glass M. Nuclear localisation of the endocannabinoid metabolizing enzyme fatty acid amide hydrolase (FAAH) in invasive trophoblasts and an association with recurrent miscarriage. Placenta 2008;29:970–975. [DOI] [PubMed] [Google Scholar]
- Chicca A, Marazzi J, Nicolussi S, Gertsch J. Evidence for bidirectional endocannabinoid transport across cell membranes. J Biol Chem 2012;287:34660–34682. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chung HW, Lee JY, Moon HS, Hur SE, Park MH, Wen Y, Polan ML. Matrix metalloproteinase-2, membranous type 1 matrix metalloproteinase, and tissue inhibitor of metalloproteinase-2 expression in ectopic and eutopic endometrium. Fertil Steril 2002;78:787–795. [DOI] [PubMed] [Google Scholar]
- Colombo N, Preti E, Landoni F, Carinelli S, Colombo A, Marini C, Sessa C. Endometrial cancer: ESMO clinical practice guidelines for diagnosis, treatment and follow-up. Ann Oncol 2013;24:vi33–vi38. [DOI] [PubMed] [Google Scholar]
- Correa F, Wolfson ML, Valchi P, Aisemberg J, Franchi AM. Endocannabinoid system and pregnancy. Reproduction 2016;152:R191–r200. [DOI] [PubMed] [Google Scholar]
- Costa MA, Fonseca BM, Keating E, Teixeira NA, Correia-da-Silva G. 2-Arachidonoylglycerol effects in cytotrophoblasts: metabolic enzymes expression and apoptosis in BeWo cells. Reproduction 2014;147:301–311. [DOI] [PubMed] [Google Scholar]
- Costa MA, Fonseca BM, Teixeira NA, Correia-da-Silva G. The endocannabinoid anandamide induces apoptosis in cytotrophoblast cells: involvement of both mitochondrial and death receptor pathways. Placenta 2015a;36:69–76. [DOI] [PubMed] [Google Scholar]
- Costa MA, Keating E, Fonseca BM, Teixeira NA, Correia-da-Silva G. 2-Arachidonoylglycerol impairs human cytotrophoblast cells syncytialization: influence of endocannabinoid signalling in placental development. Mol Cell Endocrinol 2015b;399:386–394. [DOI] [PubMed] [Google Scholar]
- Cravatt BF, Giang DK, Mayfield SP, Boger DL, Lerner RA, Gilula NB. Molecular characterization of an enzyme that degrades neuromodulatory fatty-acid amides. Nature 1996;384:83–87. [DOI] [PubMed] [Google Scholar]
- Cui N, Wang C, Zhao Z, Zhang J, Xu Y, Yang Y, Hao G. The roles of anandamide, fatty acid amide hydrolase, and leukemia inhibitory factor on the endometrium during the implantation window. Front Endocrinol 2017;8:268–268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dai D, Moulton BC, Ogle TF. Regression of the decidualized mesometrium and decidual cell apoptosis are associated with a shift in expression of Bcl2 family members. Biol Reprod 2000;63:188–195. [DOI] [PubMed] [Google Scholar]
- Das SK, Paria BC, Chakraborty I, Dey SK. Cannabinoid ligand-receptor signaling in the mouse uterus. Proc Natl Acad Sci USA 1995;92:4332–4336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Devane WA, Hanus L, Breuer A, Pertwee RG, Stevenson LA, Griffin G, Gibson D, Mandelbaum A, Etinger A, Mechoulam R. Isolation and structure of a brain constituent that binds to the cannabinoid receptor. Science 1992;258:1946–1949. [DOI] [PubMed] [Google Scholar]
- Di Marzo V Targeting the endocannabinoid system: to enhance or reduce? Nat Rev Drug Discov 2008;7:438–455. [DOI] [PubMed] [Google Scholar]
- Di Marzo V New approaches and challenges to targeting the endocannabinoid system. Nat Rev Drug Discov 2018;17:623–639. [DOI] [PubMed] [Google Scholar]
- Di Marzo V, De Petrocellis L, Sepe N, Buono A. Biosynthesis of anandamide and related acylethanolamides in mouse J774 macrophages and N18 neuroblastoma cells. Biochem J 1996;316(Pt 3): 977–984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Di Marzo V, Fontana A, Cadas H, Schinelli S, Cimino G, Schwartz JC, Piomelli D. Formation and inactivation of endogenous cannabinoid anandamide in central neurons. Nature 1994;372:686–691. [DOI] [PubMed] [Google Scholar]
- Di Marzo V, Silvestri C. Lifestyle and metabolic syndrome: contribution of the endocannabinoidome. Nutrients 2019;11:1956. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dias-Rocha CP, Almeida MM, Santana EM, Costa JCB, Franco JG, Pazos-Moura CC, Trevenzoli IH. Maternal high-fat diet induces sex-specific endocannabinoid system changes in newborn rats and programs adiposity, energy expenditure and food preference in adulthood. J Nutr Biochem 2018;51:56–68. [DOI] [PubMed] [Google Scholar]
- Dinh TP, Carpenter D, Leslie FM, Freund TF, Katona I, Sensi SL, Kathuria S, Piomelli D. Brain monoglyceride lipase participating in endocannabinoid inactivation. Proc Natl Acad Sci USA 2002;99:10819–10824. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DiPatrizio NV Endocannabinoids in the gut. Cannabis and Cannabinoid Res 2016;1:67–77. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dmitrieva N, Nagabukuro H, Resuehr D, Zhang G, McAllister SL, McGinty KA, Mackie K, Berkley KJ. Endocannabinoid involvement in endometriosis. Pain 2010;151:703–710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dunselman GA, Vermeulen N, Becker C, Calhaz-Jorge C, D'Hooghe T, De Bie B, Heikinheimo O, Horne AW, Kiesel L, Nap A et al. ESHRE guideline: management of women with endometriosis. Hum Reprod 2014;29:400–412. [DOI] [PubMed] [Google Scholar]
- El Marroun H, Tiemeier H, Steegers EA, Jaddoe VW, Hofman A, Verhulst FC, van den Brink W, Huizink AC. Intrauterine cannabis exposure affects fetal growth trajectories: the generation R study. J Am Acad Child Adolesc Psychiatry 2009;48:1173–1181. [DOI] [PubMed] [Google Scholar]
- El-Talatini MR, Taylor AH, Elson JC, Brown L, Davidson AC, Konje JC. Localisation and function of the endocannabinoid system in the human ovary. PloS One 2009;4:e4579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Farquhar C Endometriosis. BMJ 2007;334:249–253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Farquhar CM Ectopic pregnancy. Lancet 2005;366:583–591. [DOI] [PubMed] [Google Scholar]
- Fezza F, Bari M, Florio R, Talamonti E, Feole M, Maccarrone M. Endocannabinoids, related compounds and their metabolic routes. Molecules 2014;19:17078–17106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fonseca BM, Correia-da-Silva G, Taylor AH, Konje JC, Bell SC, Teixeira NA. Spatio-temporal expression patterns of anandamide-binding receptors in rat implantation sites: evidence for a role of the endocannabinoid system during the period of placental development. Reprod Biol Endocrinol 2009;7:121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fonseca BM, Correia-da-Silva G, Taylor AH, Lam PM, Marczylo TH, Bell SC, Konje JC, Teixeira NA. The endocannabinoid 2-arachidonoylglycerol (2-AG) and metabolizing enzymes during rat fetoplacental development: a role in uterine remodelling. Int J Biochem Cell Biol 2010a;42:1884–1892. [DOI] [PubMed] [Google Scholar]
- Fonseca BM, Correia-da-Silva G, Taylor AH, Lam PM, Marczylo TH, Konje JC, Bell SC, Teixeira NA. N-Acylethanolamine levels and expression of their metabolizing enzymes during pregnancy. Endocrinology 2010b;151:3965–3974. [DOI] [PubMed] [Google Scholar]
- Fonseca BM, Correia-da-Silva G, Teixeira NA. The endocannabinoid anandamide induces apoptosis of rat decidual cells through a mechanism involving ceramide synthesis and p38 MAPK activation. Apoptosis 2013;18:1526–1535. [DOI] [PubMed] [Google Scholar]
- Fonseca BM, Correia-da-Silva G, Teixeira NA. Anandamide restricts uterine stromal differentiation and is critical for complete decidualization. Mol Cell Endocrinol 2015;411:167–176. [DOI] [PubMed] [Google Scholar]
- Fonseca BM, Gonçalves D, Braga J, Mendes A, Correia-da-Silva G, Teixeira NA. Decidual NK cell-derived conditioned medium from spontaneous abortion affects endometrial stromal cell decidualisation: the endocannabinoid anandamide and tumor necrosis factor-α cross-talk. Hum Reprod 2019; Article in Press. [DOI] [PubMed] [Google Scholar]
- Fowler CJ The contribution of cyclooxygenase-2 to endocannabinoid metabolism and action. Br J Pharmacol 2007;152:594–601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fried PA The Ottawa Prenatal Prospective Study (OPPS): methodological issues and findings--it’s easy to throw the baby out with the bath water. Life Sci 1995;56:2159–2168. [DOI] [PubMed] [Google Scholar]
- Fugedi G, Molnar M, Rigo J Jr, Schonleber J, Kovalszky I, Molvarec A. Increased placental expression of cannabinoid receptor 1 in preeclampsia: an observational study. BMC Pregnancy Childbirth 2014;14:395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gaoni Y, Mechoulam R. Isolation, structure, and partial synthesis of an active constituent of hashish. J Am Chem Soc 1964;86:1646–1647. [Google Scholar]
- Gebeh AK, Willets JM, Bari M, Hirst RA, Marczylo TH, Taylor AH, Maccarrone M, Konje JC. Elevated anandamide and related N-acylethanolamine levels occur in the peripheral blood of women with ectopic pregnancy and are mirrored by changes in peripheral fatty acid amide hydrolase activity. J Clin Endocrinol Metab 2013;98:1226–1234. [DOI] [PubMed] [Google Scholar]
- Gebeh AK, Willets JM, Marczylo EL, Taylor AH, Konje JC. Ectopic pregnancy is associated with high anandamide levels and aberrant expression of FAAH and CB1 in fallopian tubes. J Clin Endocrinol Metab 2012;97:2827–2835. [DOI] [PubMed] [Google Scholar]
- Gentilini D, Besana A, Vigano P, Dalino P, Vignali M, Melandri M, Busacca M, Di Blasio AM. Endocannabinoid system regulates migration of endometrial stromal cells via cannabinoid receptor 1 through the activation of PI3K and ERK1/2 pathways. Fertil Steril 2010;93:2588–2593. [DOI] [PubMed] [Google Scholar]
- Gibbons WE Management of endometriosis in fertility patients. Fertil Steril 2004;81:1204–1205. [DOI] [PubMed] [Google Scholar]
- Grimaldi P, Pucci M, Di Siena S, Di Giacomo D, Pirazzi V, Geremia R, Maccarrone M. The faah gene is the first direct target of estrogen in the testis: role of histone demethylase LSD1. Cell Mol Life Sci 2012;69:4177–4190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gu Y, Jow GM, Moulton BC, Lee C, Sensibar JA, Park-Sarge OK, Chen TJ, Gibori G. Apoptosis in decidual tissue regression and reorganization. Endocrinology 1994;135:1272–1279. [DOI] [PubMed] [Google Scholar]
- Guida M, Ligresti A, De Filippis D, D'Amico A, Petrosino S, Cipriano M, Bifulco G, Simonetti S, Orlando P, Insabato L et al. The levels of the endocannabinoid receptor CB2 and its ligand 2-arachidonoylglycerol are elevated in endometrial carcinoma. Endocrinology 2010;151:921–928. [DOI] [PubMed] [Google Scholar]
- Guo Y, Wang H, Okamoto Y, Ueda N, Kingsley PJ, Marnett LJ, Schmid HH, Das SK, Dey SK. N-Acylphosphatidylethanolamine-hydrolyzing phospholipase D is an important determinant of uterine anandamide levels during implantation. J Biol Chem 2005;280:23429–23432. [DOI] [PubMed] [Google Scholar]
- Habayeb OM, Taylor AH, Bell SC, Taylor DJ, Konje JC. Expression of the endocannabinoid system in human first trimester placenta and its role in trophoblast proliferation. Endocrinology 2008;149:5052–5060. [DOI] [PubMed] [Google Scholar]
- Habayeb OM, Taylor AH, Evans MD, Cooke MS, Taylor DJ, Bell SC, Konje JC. Plasma levels of the endocannabinoid anandamide in women--a potential role in pregnancy maintenance and labor? J Clin Endocrinol Metab 2004;89:5482–5487. [DOI] [PubMed] [Google Scholar]
- Hanus L, Abu-Lafi S, Fride E, Breuer A, Vogel Z, Shalev DE, Kustanovich I, Mechoulam R. 2-Arachidonyl glyceryl ether, an endogenous agonist of the cannabinoid CB1 receptor. Proc Natl Acad Sci USA 2001;98:3662–3665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Helliwell RJ, Chamley LW, Blake-Palmer K, Mitchell MD, Wu J, Kearn CS, Glass M. Characterization of the endocannabinoid system in early human pregnancy. J Clin Endocrinol Metab 2004;89:5168–5174. [DOI] [PubMed] [Google Scholar]
- Helmo FR, Lopes AMM, Carneiro A, Campos CG, Silva PB, Dos Reis Monteiro MLG, Rocha LP, Dos Reis MA, Etchebehere RM, Machado JR et al. Angiogenic and antiangiogenic factors in preeclampsia. Pathol Res Pract 2018;214:7–14. [DOI] [PubMed] [Google Scholar]
- Hemberger M, Hanna CW, Dean W. Mechanisms of early placental development in mouse and humans. Nat Rev Genet 2019;21:27–43. [DOI] [PubMed] [Google Scholar]
- Heslehurst N, Rankin J, Wilkinson JR, Summerbell CD. A nationally representative study of maternal obesity in England, UK: trends in incidence and demographic inequalities in 619 323 births, 1989-2007. Int J Obes (Lond) 2010;34:420–428. [DOI] [PubMed] [Google Scholar]
- Horne AW, Phillips JA 3rd, Kane N, Lourenco PC, McDonald SE, Williams AR, Simon C, Dey SK, Critchley HO. CB1 expression is attenuated in fallopian tube and decidua of women with ectopic pregnancy. PloS One 2008;3:e3969. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Horton J, Sterrenburg M, Lane S, Maheshwari A, Li TC, Cheong Y. Reproductive, obstetric, and perinatal outcomes of women with adenomyosis and endometriosis: a systematic review and meta-analysis. Hum Reprod Update 2019;25:592–632. [DOI] [PubMed] [Google Scholar]
- Hua T, Vemuri K, Pu M, Qu L, Han GW, Wu Y, Zhao S, Shui W, Li S, Korde A et al. Crystal structure of the human cannabinoid receptor CB(1). Cell 2016;167:750, e714–762. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang NL, Juang JM, Wang YH, Hsueh CH, Liang YJ, Lin JL, Tsai CT, Lai LP. Rimonabant inhibits TNF-alpha-induced endothelial IL-6 secretion via CB1 receptor and cAMP-dependent protein kinase pathway. Acta Pharmacol Sin 2010;31:1447–1453. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang SM, Bisogno T, Trevisani M, Al-Hayani A, De Petrocellis L, Fezza F, Tognetto M, Petros TJ, Krey JF, Chu CJ et al. An endogenous capsaicin-like substance with high potency at recombinant and native vanilloid VR1 receptors. Proc Natl Acad Sci USA 2002;99:8400–8405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ibsen MS, Connor M, Glass M. Cannabinoid CB1 and CB2 receptor signaling and bias. Cannabis Cannabinoid Res 2017;2:48–60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Joseph J, Niggemann B, Zaenker KS, Entschladen F. Anandamide is an endogenous inhibitor for the migration of tumor cells and T lymphocytes. Cancer Immunol Immunother 2004;53:723–728. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaczocha M, Glaser ST, Deutsch DG. Identification of intracellular carriers for the endocannabinoid anandamide. Proc Natl Acad Sci USA 2009;106:6375–6380. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kang, JH, Goto, T, Han, IS, Kawada, T, Kim, YM, Yu, R. Dietary capsaicin reduces obesity-induced insulin resistance and hepatic steatosis in obese mice fed a high-fat diet. Obesity (Silver Spring) 2010;18:780–787. [DOI] [PubMed] [Google Scholar]
- Karwad MA, Couch DG, Theophilidou E, Sarmad S, Barrett DA, Larvin M, Wright KL, Lund JN, O'Sullivan SE. The role of CB1 in intestinal permeability and inflammation. FASEB J 2017;31:3267–3277. [DOI] [PubMed] [Google Scholar]
- Kenney SP, Kekuda R, Prasad PD, Leibach FH, Devoe LD, Ganapathy V. Cannabinoid receptors and their role in the regulation of the serotonin transporter in human placenta. Am J Obstet Gynecol 1999;181:491–497. [DOI] [PubMed] [Google Scholar]
- Kent A Diagnosing preeclampsia. Rev Obstet Gynecol 2008;1:146–147. [PMC free article] [PubMed] [Google Scholar]
- Kidima WB Syncytiotrophoblast functions and fetal growth restriction during placental malaria: updates and implication for future interventions. Biomed Res Int 2015;2015:451735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Knöfler M, Haider S, Saleh L, Pollheimer J, Gamage TKJB, James J. Human placenta and trophoblast development: key molecular mechanisms and model systems. Cell Mol Life Sci 2019;76:3479–3496. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laezza C, Pisanti S, Crescenzi E, Bifulco M. Anandamide inhibits Cdk2 and activates Chk1 leading to cell cycle arrest in human breast cancer cells. FEBS Lett 2006;580:6076–6082. [DOI] [PubMed] [Google Scholar]
- Laezza C, Pisanti S, Malfitano AM, Bifulco M. The anandamide analog, Met-F-AEA, controls human breast cancer cell migration via the RHOA/RHO kinase signaling pathway. Endocr Relat Cancer 2008;15:965–974. [DOI] [PubMed] [Google Scholar]
- Leconte M, Nicco C, Ngo C, Arkwright S, Chereau C, Guibourdenche J, Weill B, Chapron C, Dousset B, Batteux F. Antiproliferative effects of cannabinoid agonists on deep infiltrating endometriosis. Am J Pathol 2010;177:2963–2970. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee SF, Newton C, Widen R, Friedman H, Klein TW. Differential expression of cannabinoid CB(2) receptor mRNA in mouse immune cell subpopulations and following B cell stimulation. Eur J Pharmacol 2001;423:235–241. [DOI] [PubMed] [Google Scholar]
- Li X, Hua T, Vemuri K, Ho JH, Wu Y, Wu L, Popov P, Benchama O, Zvonok N, Locke K et al. Crystal structure of the human cannabinoid receptor CB2. Cell 2019;176:459–467 e413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maccarrone M Endocannabinoids: friends and foes of reproduction. Prog Lipid Res 2009;48:344–354. [DOI] [PubMed] [Google Scholar]
- Maccarrone M, Bari M, Di Rienzo M, Finazzi-Agro A, Rossi A. Progesterone activates fatty acid amide hydrolase (FAAH) promoter in human T lymphocytes through the transcription factor Ikaros. Evidence for a synergistic effect of leptin. J Biol Chem 2003;278:32726–32732. [DOI] [PubMed] [Google Scholar]
- Maccarrone, M, Bisogno, T, Valensise, H, Lazzarin, N, Fezza, F, Manna, C, Di Marzo, V, Finazzi-Agro, A. Low fatty acid amide hydrolase and high anandamide levels are associated with failure to achieve an ongoing pregnancy after IVF and embryo transfer. Mol Hum Reprod 2002;8:188–195. [DOI] [PubMed] [Google Scholar]
- Maccarrone M, Valensise H, Bari M, Lazzarin N, Romanini C, Finazzi-Agro A. Relation between decreased anandamide hydrolase concentrations in human lymphocytes and miscarriage. Lancet 2000;355:1326–1329. [DOI] [PubMed] [Google Scholar]
- Magina S, Esteves-Pinto C, Moura E, Serrao MP, Moura D, Petrosino S, Di Marzo V, Vieira-Coelho MA. Inhibition of basal and ultraviolet B-induced melanogenesis by cannabinoid CB(1) receptors: a keratinocyte-dependent effect. Arch Dermatol Res 2011;303:201–210. [DOI] [PubMed] [Google Scholar]
- Maia J, Almada M, Silva A, Correia-da-Silva G, Teixeira N, Sa SI, Fonseca BM. The endocannabinoid system expression in the female reproductive tract is modulated by estrogen. J Steroid Biochem Mol Biol 2017;174:40–47. [DOI] [PubMed] [Google Scholar]
- Marczylo TH, Lam PM, Amoako AA, Konje JC. Anandamide levels in human female reproductive tissues: solid-phase extraction and measurement by ultraperformance liquid chromatography tandem mass spectrometry. Anal Biochem 2010;400:155–162. [DOI] [PubMed] [Google Scholar]
- Matsuda LA, Lolait SJ, Brownstein MJ, Young AC, Bonner TI. Structure of a cannabinoid receptor and functional expression of the cloned cDNA. Nature 1990;346:561–564. [DOI] [PubMed] [Google Scholar]
- McHugh D, Page J, Dunn E, Bradshaw HB. Delta(9) -Tetrahydrocannabinol and N-arachidonyl glycine are full agonists at GPR18 receptors and induce migration in human endometrial HEC-1B cells. Br J Pharmacol 2012;165:2414–2424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mechoulam R, Ben-Shabat S, Hanus L, Ligumsky M, Kaminski NE, Schatz AR, Gopher A, Almog S, Martin BR, Compton DR et al. Identification of an endogenous 2-monoglyceride, present in canine gut, that binds to cannabinoid receptors. Biochem Pharmacol 1995;50:83–90. [DOI] [PubMed] [Google Scholar]
- Meuleman C, Vandenabeele B, Fieuws S, Spiessens C, Timmerman D, D'Hooghe T. High prevalence of endometriosis in infertile women with normal ovulation and normospermic partners. Fertil Steril 2009;92:68–74. [DOI] [PubMed] [Google Scholar]
- Miranda RA, De Almeida MM, Rocha CPDD, de Brito Fassarella L, De Souza LL, Souza AFPD, Andrade CBVD, Fortunato RS, Pazos-Moura CC, Trevenzoli IH. Maternal high-fat diet consumption induces sex-dependent alterations of the endocannabinoid system and redox homeostasis in liver of adult rat offspring. Sci Rep 2018;8:14751–14751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mitchell MD, Rice GE, Vaswani K, Kvaskoff D, Peiris HN. Differential regulation of eicosanoid and endocannabinoid production by inflammatory mediators in human choriodecidua. PloS one 2016;11:e0148306–e0148306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moghadam KK, Kessler CA, Schroeder JK, Buckley AR, Brar AK, Handwerger S. Cannabinoid receptor I activation markedly inhibits human decidualization. Mol Cell Endocrinol 2005;229:65–74. [DOI] [PubMed] [Google Scholar]
- Molvarec A, Fugedi G, Szabo E, Stenczer B, Walentin S, Rigo J Jr. Decreased circulating anandamide levels in preeclampsia. Hypertens Res 2015;38:413–418. [DOI] [PubMed] [Google Scholar]
- Morice P, Leary A, Creutzberg C, Abu-Rustum N, Darai E. Endometrial cancer. Lancet 2016;387:1094–1108. [DOI] [PubMed] [Google Scholar]
- Munro S, Thomas KL, Abu-Shaar M. Molecular characterization of a peripheral receptor for cannabinoids. Nature 1993;365:61–65. [DOI] [PubMed] [Google Scholar]
- Nicolussi S, Gertsch J. Endocannabinoid transport revisited. Vitam Horm 2015;98:441–485. [DOI] [PubMed] [Google Scholar]
- Okada H, Tsuzuki T, Murata H. Decidualization of the human endometrium. Reprod Med Biol 2018;17:220–227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pacher P, Bátkai S, Kunos G. Blood pressure regulation by endocannabinoids and their receptors. Neuropharmacology 2005;48:1130–1138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pandey R, Mousawy K, Nagarkatti M, Nagarkatti P. Endocannabinoids and immune regulation. Pharmacol Res 2009;60:85–92. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paria BC, Das SK, Dey SK. The preimplantation mouse embryo is a target for cannabinoid ligand-receptor signaling. Proc Natl Acad Sci USA 1995;92:9460–9464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paria BC, Song H, Wang X, Schmid PC, Krebsbach RJ, Schmid HH, Bonner TI, Zimmer A, Dey SK. Dysregulated cannabinoid signaling disrupts uterine receptivity for embryo implantation. J Biol Chem 2001;276:20523–20528. [DOI] [PubMed] [Google Scholar]
- Park B, Gibbons HM, Mitchell MD, Glass M. Identification of the CB1 cannabinoid receptor and fatty acid amide hydrolase (FAAH) in the human placenta. Placenta 2003;24:990–995. [DOI] [PubMed] [Google Scholar]
- Parolaro D Presence and functional regulation of cannabinoid receptors in immune cells. Life Sci 1999;65:637–644. [DOI] [PubMed] [Google Scholar]
- Pertwee RG Emerging strategies for exploiting cannabinoid receptor agonists as medicines. Br J Pharmacol 2009;156:397–411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Piscitelli F, Carta G, Bisogno T, Murru E, Cordeddu L, Berge K, Tandy S, Cohn JS, Griinari M, Banni S et al. Effect of dietary krill oil supplementation on the endocannabinoidome of metabolically relevant tissues from high-fat-fed mice. Nutr Metab (Lond) 2011;8:51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pizzo A, Salmeri FM, Ardita FV, Sofo V, Tripepi M, Marsico S. Behaviour of cytokine levels in serum and peritoneal fluid of women with endometriosis. Gynecol Obstet Invest 2002;54:82–87. [DOI] [PubMed] [Google Scholar]
- Portella G, Laezza C, Laccetti P, De Petrocellis L, Di Marzo V, Bifulco M. Inhibitory effects of cannabinoid CB1 receptor stimulation on tumor growth and metastatic spreading: actions on signals involved in angiogenesis and metastasis. FASEB J 2003;17:1771–1773. [DOI] [PubMed] [Google Scholar]
- Porter AC, Sauer JM, Knierman MD, Becker GW, Berna MJ, Bao J, Nomikos GG, Carter P, Bymaster FP, Leese AB et al. Characterization of a novel endocannabinoid, virodhamine, with antagonist activity at the CB1 receptor. J Pharmacol Exp Ther 2002;301:1020–1024. [DOI] [PubMed] [Google Scholar]
- Raborn ES, Marciano-Cabral F, Buckley NE, Martin BR, Cabral GA. The cannabinoid delta-9-tetrahydrocannabinol mediates inhibition of macrophage chemotaxis to RANTES/CCL5: linkage to the CB2 receptor. J Neuroimmune Pharmacol 2008;3:117–129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raichlen, DA, Foster, AD, Gerdeman, GL, Seillier, A, Giuffrida, A. Wired to run: exercise-induced endocannabinoid signaling in humans and cursorial mammals with implications for the ‘runner’s high’. J Exp Biol 2012;215:1331–1336. [DOI] [PubMed] [Google Scholar]
- Raichlen DA, Foster AD, Seillier A, Giuffrida A, Gerdeman GL. Exercise-induced endocannabinoid signaling is modulated by intensity. Eur J Appl Physiol 2013;113:869–875. [DOI] [PubMed] [Google Scholar]
- Redman CW, Sargent IL. Preeclampsia and the systemic inflammatory response. Semin Nephrol 2004;24:565–570. [DOI] [PubMed] [Google Scholar]
- Resuehr D, Glore DR, Taylor HS, Bruner-Tran KL, Osteen KG. Progesterone-dependent regulation of endometrial cannabinoid receptor type 1 (CB1-R) expression is disrupted in women with endometriosis and in isolated stromal cells exposed to 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD). Fertil Steril 2012;98:948, e941–956. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ribeiro ML, Vercelli CA, Sordelli MS, Farina MG, Cervini M, Billi S, Franchi AM. 17beta-oestradiol and progesterone regulate anandamide synthesis in the rat uterus. Reprod Biomed Online 2009;18:209–218. [DOI] [PubMed] [Google Scholar]
- Rio CD, Millan E, Garcia V, Appendino G, DeMesa J, Munoz E. The endocannabinoid system of the skin. A potential approach for the treatment of skin disorders. Biochem Pharmacol 2018;157:122–133. [DOI] [PubMed] [Google Scholar]
- Rodriguez de Fonseca F, Navarro M, Gomez R, Escuredo L, Nava F, Fu J, Murillo-Rodriguez E, Giuffrida A, LoVerme J, Gaetani S et al. An anorexic lipid mediator regulated by feeding. Nature 2001;414:209–212. [DOI] [PubMed] [Google Scholar]
- Rossato M, Pagano C, Vettor R. The cannabinoid system and male reproductive functions. J Neuroendocrinol 2008;20:90–93. [DOI] [PubMed] [Google Scholar]
- Rousseaux C, Thuru X, Gelot A, Barnich N, Neut C, Dubuquoy L, Dubuquoy C, Merour E, Geboes K, Chamaillard M et al. Lactobacillus acidophilus modulates intestinal pain and induces opioid and cannabinoid receptors. Nat Med 2007;13:35–37. [DOI] [PubMed] [Google Scholar]
- Rouzer CA, Marnett LJ. Endocannabinoid oxygenation by cyclooxygenases, lipoxygenases, and cytochromes P450: cross-talk between the eicosanoid and endocannabinoid signaling pathways. Chem Rev 2011;111:5899–5921. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ryberg E, Larsson N, Sjogren S, Hjorth S, Hermansson NO, Leonova J, Elebring T, Nilsson K, Drmota T, Greasley PJ. The orphan receptor GPR55 is a novel cannabinoid receptor. Br J Pharmacol 2007;152:1092–1101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanchez AM, Cioffi R, Vigano P, Candiani M, Verde R, Piscitelli F, Di Marzo V, Garavaglia E, Panina-Bordignon P. Elevated systemic levels of endocannabinoids and related mediators across the menstrual cycle in women with endometriosis. Reprod Sci 2016;23:1071–1079. [DOI] [PubMed] [Google Scholar]
- Sanchez AM, Quattrone F, Pannese M, Ulisse A, Candiani M, Diaz-Alonso J, Velasco G, Panina-Bordignon P. The cannabinoid receptor CB1 contributes to the development of ectopic lesions in a mouse model of endometriosis. Hum Reprod 2017;32:175–184. [DOI] [PubMed] [Google Scholar]
- Sanchez AM, Vigano P, Mugione A, Panina-Bordignon P, Candiani M. The molecular connections between the cannabinoid system and endometriosis. Mol Hum Reprod 2012;18:563–571. [DOI] [PubMed] [Google Scholar]
- Schmid PC, Wold LE, Krebsbach RJ, Berdyshev EV, Schmid HH. Anandamide and other N-acylethanolamines in human tumors. Lipids 2002;37:907–912. [DOI] [PubMed] [Google Scholar]
- Scotchie JG, Savaris RF, Martin CE, Young SL. Endocannabinoid regulation in human endometrium across the menstrual cycle. Reprod Sci 2015;22:113–123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shao Z, Yin J, Chapman K, Grzemska M, Clark L, Wang J, Rosenbaum DM. High-resolution crystal structure of the human CB1 cannabinoid receptor. Nature 2016;540:602–606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shaw JLV, Dey SK, Critchley HOD, Horne AW. Current knowledge of the aetiology of human tubal ectopic pregnancy. Hum Reprod Update 2010;16:432–444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shen X, Duan H, Wang S, Hong W, Wang YY, Lin SL. Expression of cannabinoid receptors in myometrium and its correlation with dysmenorrhea in adenomyosis. Reprod Sci 2019;1933719119833483. [DOI] [PubMed] [Google Scholar]
- Slipetz DM, O'Neill GP, Favreau L, Dufresne C, Gallant M, Gareau Y, Guay D, Labelle M, Metters KM. Activation of the human peripheral cannabinoid receptor results in inhibition of adenylyl cyclase. Mol Pharmacol 1995;48:352–361. [PubMed] [Google Scholar]
- Sugiura T, Kondo S, Sukagawa A, Nakane S, Shinoda A, Itoh K, Yamashita A, Waku K. 2-Arachidonoylglycerol: a possible endogenous cannabinoid receptor ligand in brain. Biochem Biophys Res Commun 1995;215:89–97. [DOI] [PubMed] [Google Scholar]
- Sun X, Dey SK. Endocannabinoid signaling in female reproduction. ACS Chem Nerosci 2012;3:349–355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun X, Xie H, Yang J, Wang H, Bradshaw HB, Dey SK. Endocannabinoid signaling directs differentiation of trophoblast cell lineages and placentation. Proc Natl Acad Sci USA 2010;107:16887–16892. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun, Y, Alexander, SP, Kendall, DA, Bennett, AJ Cannabinoids and PPARalpha signalling. Biochem Soc Trans 2006;34:1095–1097. [DOI] [PubMed] [Google Scholar]
- Syed SK, Bui HH, Beavers LS, Farb TB, Ficorilli J, Chesterfield AK, Kuo MS, Bokvist K, Barrett DG, Efanov AM. Regulation of GPR119 receptor activity with endocannabinoid-like lipids. Am J Physiol Endocrinol Metab 2012;303:E1469–E1478. [DOI] [PubMed] [Google Scholar]
- Taylor AH, Abbas MS, Habiba MA, Konje JC. Histomorphometric evaluation of cannabinoid receptor and anandamide modulating enzyme expression in the human endometrium through the menstrual cycle. Histochem Cell Biol 2010;133:557–565. [DOI] [PubMed] [Google Scholar]
- Taylor AH, Finney M, Lam PM, Konje JC. Modulation of the endocannabinoid system in viable and non-viable first trimester pregnancies by pregnancy-related hormones. Reprod Biol Endocrinol 2011;9:152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tibiriça E The multiple functions of the endocannabinoid system: a focus on the regulation of food intake. Diabetol Metab Syndr 2010;2:5–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Torre LA, Bray F, Siegel RL, Ferlay J, Lortet-Tieulent J, Jemal A. Global cancer statistics, 2012. CA Cancer J Clin 2015;65:87–108. [DOI] [PubMed] [Google Scholar]
- Trabucco E, Acone G, Marenna A, Pierantoni R, Cacciola G, Chioccarelli T, Mackie K, Fasano S, Colacurci N, Meccariello R et al. Endocannabinoid system in first trimester placenta: low FAAH and high CB1 expression characterize spontaneous miscarriage. Placenta 2009;30:516–522. [DOI] [PubMed] [Google Scholar]
- Varma R, Gupta J. Tubal ectopic pregnancy. BMJ Clin Evid 2009;2009. [PMC free article] [PubMed] [Google Scholar]
- Velasco G, Galve-Roperh I, Sanchez C, Blazquez C, Haro A, Guzman M. Cannabinoids and ceramide: two lipids acting hand-by-hand. Life Sci 2005;77:1723–1731. [DOI] [PubMed] [Google Scholar]
- Walker OS, Holloway AC, Raha S. The role of the endocannabinoid system in female reproductive tissues. J Ovarian Res 2019;12:3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang H, Guo Y, Wang D, Kingsley PJ, Marnett LJ, Das SK, DuBois RN, Dey SK. Aberrant cannabinoid signaling impairs oviductal transport of embryos. Nat Med 2004;10:1074–1080. [DOI] [PubMed] [Google Scholar]
- Wang H, Xie H, Guo Y, Zhang H, Takahashi T, Kingsley PJ, Marnett LJ, Das SK, Cravatt BF, Dey SK. Fatty acid amide hydrolase deficiency limits early pregnancy events. J Clin Invest 2006;116:2122–2131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang H, Xie H, Sun X, Kingsley PJ, Marnett LJ, Cravatt BF, Dey SK. Differential regulation of endocannabinoid synthesis and degradation in the uterus during embryo implantation. Prostaglandins Other Lipid Mediat 2007;83:62–74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zoratti C, Kipmen-Korgun D, Osibow K, Malli R, Graier WF. Anandamide initiates Ca(2+) signaling via CB2 receptor linked to phospholipase C in calf pulmonary endothelial cells. Br J Pharmacol 2003;140:1351–1362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zou S, Kumar U. Cannabinoid receptors and the endocannabinoid system: Signaling and function in the central nervous system. Int J Mol Sci 2018;19:833. [DOI] [PMC free article] [PubMed] [Google Scholar]