
Figure 2.
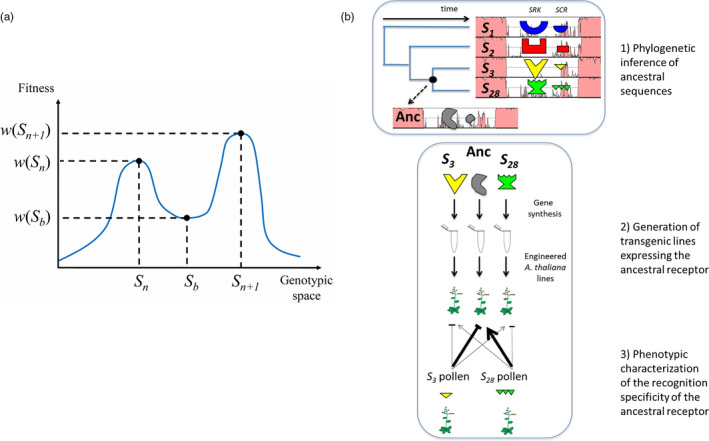
Theoretical (a) and empirical (b) approaches to study allelic diversification at the S‐locus. (a) Emergence of new S‐alleles in two‐component genetic systems involves the crossing of a fitness valley (see section 2.2). A simplified fitness landscape showing diversification in the two‐component genetic model of Uyenoyama et al. (2001) and Gervais et al. (2011). On the X‐axis is represented a simplified 1D genotypic space where the different haplotypes are placed. Sn: ancestral SI haplotype, Sb: intermediate SC mutant haplotype on the male component, Sn+1: newly formed haplotype. Each genotype has an associated relative fitness (w) on the Y‐axis that depends on the combination of four evolutionary forces: transmission advantage of SC haplotypes, the universal compatibility of pollen Sb, negative frequency‐dependent selection and inbreeding depression and proportion of selfed offspring. (b) An experimental approach based on ancestral protein resurrection to test evolutionary scenarios by which new SI alleles arise (see section 2.3 for details of the experiment). The asymmetrical recognition phenotype of the putative ancestor (Anc) shows that the S3 pollen but not S28 pollen has retained the capacity to trigger a SI reaction by the ancestral receptor. Hence, the ancestral recognition specificity has been conserved over a time at least equal to the divergence between S3 and S28, rejecting models predicting rapid turnover of SI specificities. Note that these results also reject models of dual‐specificity intermediates, where the ancestral receptor would have recognized and rejected both S3 and S28 pollen