
Figure 3.
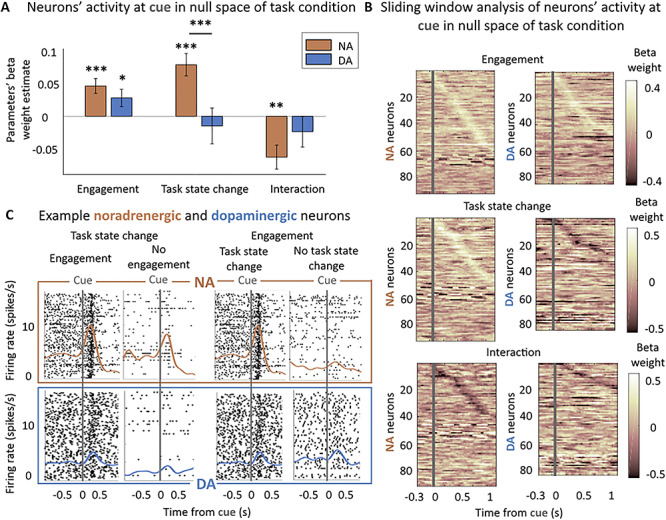
Noradrenergic but not dopaminergic neurons are sensitive to the task state change. (A) Sensitivity to engagement and task state change in null space of task condition at cue (0–500 ms from cue onset). Multilevel linear regression of neurons’ activity by: engagement, trial repetition (task state change), and their interaction. Bars and error bars represent means of the beta weights ± SEM. Noradrenergic neurons were sensitive to the task sate change, the engagement and their interaction (all P < 0.01). Dopaminergic neurons were only significantly sensitive to the engagement (P < 0.05). * P < 0.05; ** P ≤ 0.01; *** P ≤ 0.001. (B) Sliding window analysis of the sensitivity of the firing rate to engagement, task state change, and their interaction in null space of task condition at cue. Multilevel linear regression of neurons’ activity by: engagement, trial repetition (task state change), and their interaction in 200 ms windows with an increment of 25 ms. X-axis represents the center of the window. Y-axis corresponds to each neuron. Color code represents the value of the beta weight in the regression. Neurons are ordered (top to bottom) by latency of their peak if it was significant (P < 0.05). (C) Example noradrenergic and dopaminergic neurons. Neuronal activity of two representative neurons around the cue onset (gray vertical line). Each panel corresponds to a different number of trials (each trial is a line in the raster plot). Top: spike activity (raster and spike density function) of a noradrenergic neuron showing a strong activation at cue. The activation is slightly stronger in engaged versus nonengaged trials (β = 0.04 in regression shown in Fig. 3A) (all task conditions pooled together in nonrepeated trials (task state change) only: 156 and 25 trials) and much stronger (β = 0.36) for nonrepeated (task state change) versus repeated trials (no task state change a) (engage trials only: 156 and 63 trials). Bottom: same but for a dopaminergic neuron showing an intermediate activation at cue onset. Its activity was greater for engaged than nonengaged trials (β = 0.23, 153 and 27 trials) but was not modulated by the task state change of the cue (153 and 57 trials). Note that even though the baseline firing of this example dopaminergic neuron appears modulated by the engagement, it is not the case (P = 0.14) and actually reflects the sensitivity of this particular neuron to trial number (β = −0.16, P = 0.01), which is not reliable at the population level (P = 0.08).