

Abstract
Dermatophytosis, a zoonotic disease, is caused by fungi of three main genera, namely, Micropsorum, Trichophyton, and Epidermophyton. Specific lesions of dermatophyte infections are localized in the face, legs, and/or tail. Skin lesions in infected animals demonstrate localized alopecia, erythema, and crust, which are more commonly known as ringworm. Factors that affect dermatophytosis include the dermatophyte species; virulence factors of the agent; and the immune status, age, and sex of the host. High levels of cortisol and pro-inflammatory cytokines have also been reported to play an important role in dermatophyte infection. This review aims to explore and understand factors that affect dermatophyte infection with an emphasis on the prevalence, clinical signs, pathogenesis, immune response, and the roles of cortisol and cytokines in companion animals infected by a dermatophyte.
Keywords: clinical signs, cortisol, cytokine, dermatophytosis, immune response, pathogenesis
Introduction
Dermatophytosis is a zoonotic disease caused by fungal infection of a species of dermatophyte [1,2]. Dermatophytosis is more commonly known as ringworm, which is macroscopically characterized by multifocal alopecia and crust on the skin with a specific formation [3,4]. This disease is distributed globally and has gained special attention in public health [2,5]. Dermatophyte infections in humans occur after contact with contaminated products or specimens, such as soil, hair, or crust on the epidermal layer of infected animals [5].
Annual cases of dermatophytosis have increased not only in humans but also in animals, particularly dogs and cats [6]. In Indonesia, cases of dermatophytosis are reported to be more prevalent in adult female cats and kittens than male cats [1]. The results of the research on dogs showed that 34% of dogs in Yogyakarta, Indonesia, were positive for dermatophytosis [7]. In Europe, the incidence of dermatophytosis in dogs and cats ranges from 20 to 30% [8]. The results of various studies concluded that the main species causing dermatophytosis in pets is Microsporum canis, in 81.8%-97% of the cases [1,2,6]. The incidence of zoonotic dermatophytes M. canis was the highest in dogs, cats, and human (60.0%) compared to other species [9]. High levels of cortisol over a long period of time modulate the immune response and induce immune suppression as the result of lymphatic tissue atrophy [8]. Stress also induces cortisol production and results in inhibition of the Th1 cytokine pro-inflammatory response and upregulates the Th2 anti-inflammatory cytokine response [8,10]. It has been reported that high levels of cortisol in cats, as a consequence of stress, alter host immune responses and might promote the dermatophyte infections [10].
This review discusses the prevalence, clinical signs, pathogenesis, immune responses, and roles of cortisol and cytokines in companion animals infected with a dermatophyte.
Etiology and Prevalence
Dermatophytes consist of 40 species of fungi derived from three genera, namely, Micropsorum, Trichophyton, and Epidermophyton [11,12]. The two genera of fungi which are the main causes of dermatophytosis in animals (especially in dogs and cats) are Microsporum spp. and Trichophyton spp. M. canis, Microsporum gypseum, Trichophyton mentagrophytes, Trichophyton equinum, Trichophyton verrucosum, and Microsporum nanum species of fungi have important roles in veterinary medicine [3,13,14]. Common dermatophytes that infect small animals include M. canis, T. verrucosum, and T. mentagrophytes [15]. This study also showed that T. mentagrophytes is often found in guinea pigs and rabbits [15]. A colony of M. canis and T. mentagrophytes is shown in Figures-1 and 2, respectively.
Figure-1.
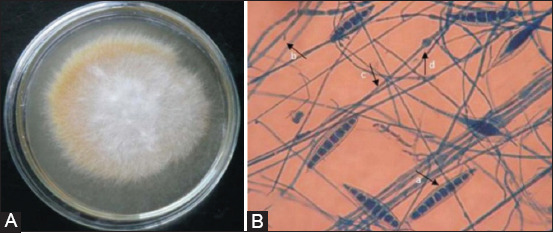
Colony of Microsporum canis on SDA media (14 days). Colony is cottonny, white to buff in color; with increasing age becomes brownish yellow (A); artrospora of M. canis viewed under microscope (B): a. macroconidia, b. microconidia, c. hyphae, d. chlamydoconidia, viewed under microscope with lacto phenol cotton blue staining [7].
Figure-2.
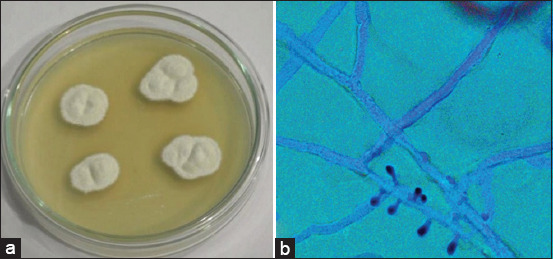
Colony of Trichophyton rubrum on SDA media (14 days). Colony is white, fluffy, with a central umbo and flat periphery (a); microaleurispores of T. rubrum are scanty and slim viewed under microscope with lacto phenol cotton blue staining (b) (Private Documentation, 2020).
More than 90% of dermatophyte infections in cats are caused by M. canis. M. canis also infects humans and other animal species such as dogs, cattle, horses, pigs, goats, rabbits, guinea pigs, apes, monkey, tigers, and mice [6]. A study in Lisbon, Portugal, showed that 82% of 89 cats positive for dermatophytosis were infected with M. canis [16]. Subclinical infection of M. canis also occurs in long-haired juvenile cats [6]. An Indonesian study reported that 17 (56.7%) of 30 cats with dermatitis infections are positive for dermatophytosis caused by M. canis [1]. Several studies also showed that dermatophytosis occurs more often in female animals than in males [3,17]. Thus, sex and age play important roles in the prevalence of dermatophytosis in animals.
In a retrospective study in dogs (1970-2002), T. mentagrophytes were found in 66 cases (66/3854, 1.7%), while M. canis was diagnosed in 840 cases (840/3854, 21.8%) [18]. A study to determine dermatophyte distribution in dogs and cats in West Turkey found that 14.4% of samples tested were positive for dermatophytosis in a sample size of 326 [19]. In Italy, the reported prevalence of dermatophytosis was 7.5%-20.5% in dogs and 24.7-33.3% in cats [8]. In Eastern India, out of 1209 samples from dogs and 292 samples from cats, 253 (20.93%) and 109 (37.33%), respectively, were positive for dermatophyte spores [20]. It was reported that 34% dogs in Yogyakarta, Indonesia, are positive for dermatophytosis [7]. A study in Baku in the Middle East showed that 108 of 193 dogs and cats were infected with a dermatophyte [21].
Dermatophytosis occurs not only in small animals but also in other species of animal. A study of dermatophytosis in farm animals done in Beheira, Egypt, demonstrated that 74% of 150 samples collected from different species of animals were dermatophyte positive. The highest dermatophytosis case rates reported occurred in sheep (78%), followed by buffaloes (76%), cattle (72%), and horses (68%) [3]. The samples from female camels farmed in Central Saudi Arabia showed a dermatophytosis prevalence of 11.5%. Interestingly, the higher incidence occurred at a young age [17]. A study showed that prevalence of Arabian horse dermatophytosis in Egypt was 16.8% of the total prevalence of 81% [22]. It was reported that the prevalence of dermatophytosis in cattle farms in Irbid, Jordan, ranges from 10 to 100% [23]. That study also showed that 115 (30.6%) of 375 cattle were identified as having a typical macroscopic lesion of dermatophytosis.
Clinical Signs
Lesions in cases of dermatophytosis are variable for each species of animal [1,24-26], the most common clinical symptoms being hair loss, skin crust, erythema, and pruritus (Figures-3 and 4). Other studies showed that dermatophyte infection in dogs cause lesions localized to the face, legs, and/or tail [27]. The previous studies [1,19,26] have demonstrated that infected dogs clinically showed lesions in the skin such as multifocal alopecia, erythema, papule, pustule, scale, and crust with a distinctive formation known as ringworm. Infection of M. canis results in alopecia (1-3 mm) in the infected area and then permanent alopecia occurs, especially if the inflammatory reaction lasts a long period of time [28]. Lesions caused by dermatophytes can be mild to severe depending on several factors, including the infecting dermatophyte species, virulence factors, area of infection, secondary infections, and environmental conditions [29].
Figure-3.

Lesions from Microsporum canis in body part of cat. Formation of round shape lesions in form alopecia, and erythema in the skin [7].
Figure-4.
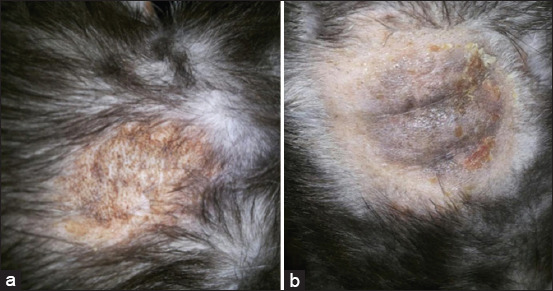
Lesions from dermatophyte infection in the body part of cat with crust and alopecia in the skin (a), hyperpigmentation (b) (Private Documentation, 2020).
In rabbits, macroscopic lesions caused by fungal infection include yellowish-white dry crust formation in the face, eyelids, ears, nails, legs, and back area, alopecia, and erythema [30]. Round shaped, white, gray, or blackish brown lesions (hyperpigmentation) along with hyperkeratosis, scales, and crust are found on the skin of Balinese cattle (Bos javanicus) infected by dermatophytes [31].
Based on the previous studies, it can be concluded that the clinical symptoms of dermatophyte infection vary depending on infected the dermatophyte species, infected area of the body, and the species of the host. In general, dermatophyte infections cause the formation of lesions; namely, alopecia, crust, papula, pustule, and erythema in the skin and are usually in a round shape known as ringworm.
Pathogenesis
At the start of infection, dermatophytes attach to the keratin tissue in the nails or hair of the host [32,33]. Virulence factors of the dermatophyte, the immunological status, and the age of the host play an important role in the progression of this disease. The young and the old among animals are reported to be more susceptible to direct or indirect dermatophyte infection [2,14].
Predisposing factors affecting dermatophytosis include the number of infective spores, frequency of transmission, health conditions, and the physiological stress experienced by animals [26,34]. The route of infection occurs through skin wounds, scars, or burns [34]. This infection occurs either directly through contact with sick cats or indirectly through blankets, bed covers, toys, cages, clothes, and other objects contaminated with spores [26,34].
The incubation period of M. canis ranges from 1 to 4 weeks [26]. The attachment of arthroconidia to keratin occurs maximally at 6 h after infection [34]. The rapidity of attachment of arthroconidia to keratin is affected by the number of infecting arthroconidia [35]. Germination and invasion of the stratum corneum take approximately 4-6 h [34,35,36]. The colonization processes result in various immune system reactions in the host including inflammation [37]. Subsequently, an inflammation reaction can be seen in infected skin area as redness, swelling, and alopecia [36,38].
Dermatophytes have the ability to produce proteolytic and keratolytic enzymes that enable keratin to be used as the sole source of nutrition after colonization, facilitating fungal growth in the stratum corneum, and resulting in keratinization in the epidermis [6,33,34]. Proteolytic activity of dermatophyte by releasing serine proteinase (urokinase and activator plasminogen tissue), which causes damage of external protein of the host and facilitated by the process of injury in skin [34,38]. The ability of dermatophytes to secrete enzymes as mentioned above is one of the virulence factors of dermatophyte infection [33,36]. Hence, the ability of dermatophytes to infect the host depends on the dermatophyte species, the number of infection spores, virulence factors, and the immune status of the host. The route of dermatophyte infection is shown in Figure-5.
Figure-5.
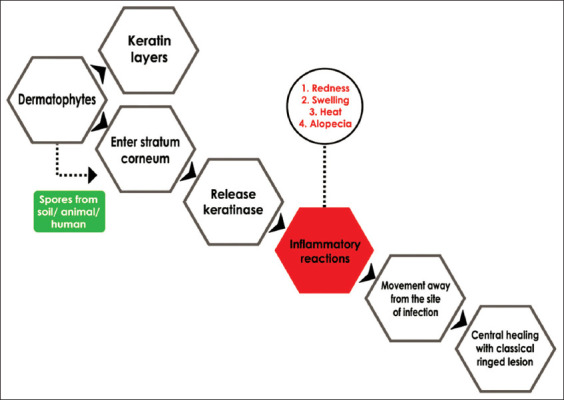
The schematic route of entry of dermatophytes into the host [38].
Host Immune Responses to Dermatophyte Infection
Immune responses from fungal infection vary dependent on the type of infectious agent [32]. During the infection process, dermatophyte interferes the host’s defense mechanism which affects the manifestation and severity of dermatophyte infection, depends on patient’s immune response [36,39,40]. Dermatophytes have non-specific antigens, namely, glycopeptide and keratinase [33]. The glycopeptide stimulates cellular immune response, while the polysaccharide part of the glycopeptide stimulates humoral immune response [33,34].
The previous studies reported that several cell wall components of dermatophyte might act as pathogen-associated molecular patterns that are recognized by pattern recognition receptors (PRRs), including toll-like receptors (TLRs), which expressed by phagocytes and dendritic cells (DCs) [36,41-43]. Following dermatophyte infection, TLR2 signaling induces the production of inflammatory cytokines and leukotrienes, such as tumor-necrosis factor (TNF), Interleukin (IL)-1b, and IL-10 [43-48]. During the inflammatory process, PRRs activate neutrophils and macrophages to produce powerful antifungal defectins. This antifungal acts by disturbing osmotic imbalance in the fungal cells. Activated macrophages and neutrophils also release massive production of cytokines, namely, IL-1, IL-12, and TNF, which are toxic to fungal cells [40,42].
Skin is a physical barrier that prevents fungal infection through epidermal cell proliferation and keratinization. Physical barriers of the skin play an important role in immune responses at the first stage of infection by preventing and eliminating fungal infiltration on the stratum corneum [35,48,49]. During inflammatory processes in the skin, keratinocytes express the major histocompatibility complex II, cytokines, and colony-stimulating factor [36,38,47]. Other components of the immune response in the skin are Langerhans cells (LCs), DCs, macrophages, natural killer (NK) cells, and memory T cells [47]. Thus, recognition dermatophytes antigens by PRRs on phagocytes and keratinocytes result in release inflammatory cytokines, which play an important role on host immune response.
Role of Cortisol and Dermatophyte Infection
Stress hormones (glucocorticoid, catecholamine, and neuroendocrine) can modulate various aspects of the immune system. Stress hormones also have direct effects on all cells of the immune system and influence the expression of various cytokines [50,51]. Glucocorticoid qualitatively and quantitatively induces the suppression of immune system of the host against invasive fungal infection [52]. Following stress, the concentration of glucocorticoid increases through the regulation of cellular and humoral immunity and results in host susceptibility to the fungal infection [53]. Chronic stress increases the concentration of glucocorticoid over a long period of time and downregulates the synthesis of pro-inflammatory cytokines, namely, TNF-α, Interferon (IFN)-γ, and IL-2 in animals and humans [53]. Glucocorticoids suppress the production of IL-12 by antigen presenting cells (APC) and downregulates the expression of IL-12 receptors on NK and T cells [50,51]. During fungal infection, the high concentrations of cortisol affect the normal function of the immune system [52,54]. Chronic high levels of cortisol in cats, as a consequence of unidentified stressors or for endogenous reasons, could alter immune responses and might promote dermatophyte infections in the presence of a contaminated environment [8]. A stress study in fish demonstrated the same phenomena. High concentrations of cortisol distract the fish immune responses against pathogenic agents [55]. Hence, cortisol plays an important role during fungal infection through its ability to suppress the host immune system by downregulating TNF-α, IFN-γ, and IL-2 cytokines.
Role of Cytokines and Dermatophyte Infection
Cytokines have important effects on the activity of many cells. Cytokines are reported to be important because of their significance in regulating the immune system [56-58]. Fungal infection in skin induces Th1 cells as a humoral immune response. During fungal infection, Th1 cells secrete pro-inflammatory cytokines such as IL-2, IL-12, IL-18, and IFN-γ which stimulates phagocytic activity, cytotoxic CD4+ T cell generation, and prevents the hypersensitivity reaction [3,59,60].
CD4+ is an important component of cellular immune responses [61]. The presence of pathogenic agents will activate naïve CD4+ cells to differentiate into various Th cell groups (Th1, Th2, and Th17) [61,62]. Cytokines such as IL-1, IL-6, IL-8, IL-10, IL-15, TNF-α , and transforming growth factor-β also have immunological roles in responses to fungal infections [63]. During fungal infection, IL-12 and IL-10 act as regulators of Th cell growth and humoral immune responses [6,59,63].
Fungal infection is capable of inducing IL-17 production as a humoral immune response during the initial phase of infection [64]. IL-17 is known as a pro-inflammatory cytokine which provides complex connections between humoral and cellular immunity; this connection is the center of inflammatory responses to fungal infection [64]. IL-17 is also reported to be responsible for preventing uncontrolled fungal growth and inflammatory reactions in M. canis infection [65]. Studies have demonstrated that patients deficient in IL-17 are susceptible to dermatophyte infection [65-67].
IL-1 is a pro-inflammatory cytokine that is produced in response of host humoral immunity to fungal infection [68]. IL-1 consists of 11 components, including IL-1α, IL-1β, and IL-1Ra which are important for chronic inflammatory infection [69]. It was reported that the process of attachment to and fungal germination in host cell walls strongly induced the release of IL-1 [70]. IL-1 is reported to have an important role in the pathogenesis of chronic cavitary pulmonary aspergillosis (CCPA), due to its ability to induce activation of IL-1α and IL-1β pro-inflammatory cytokines [71].
IL-2 is produced from Th1 cells, CD4+, CD8+, cytotoxic T cells (CTL), DC, NK cells, and NKT cells [6,72]. IL-2 has dual functions as both an immune suppressor and/or activator system [73]. Therefore, the pro-inflammatory function from IL-2 can be inhibited by antigen-specific T reg. Low amounts of IL-2 and IFN-γ secretion in response to antigen produced from pathogenic fungal indicated a deficiency of cell-mediated immunity in infected patients. Low concentrations of IL-2, IL-17, IFN-γ, and IL-10 also indicated that fungal antigens were unable to induce cytokine secretion from Th1 cells, and leads to upregulated cytokine secretion from Th2 cells [72,74]. Therefore, in fungal infection low concentrations of IL-2 correlate with upregulated levels of Th2 cytokine secretion.
IL-10 has the ability to modulate natural immune responses and prevents pro-inflammatory cytokine production from Th1 and Th2 cell secretion, such as IL-2 and IFN-γ [74,75]. IL-10 is produced by monocytes and macrophages, while other cells also produce cytokines such as DC cells, B cells, CTL, γδ-T cells, NK cells, mast cells, and neutrophil granulocytes [75,76]. Several studies showed that IL-10 prevents APC (monocytes, macrophages, and DC) actions [77,78]. IL-10 is known to have a strong inhibitory effect on fungal infection. It is known that mice deficient in IL-10 more susceptible to fungal infection [79]. Patients with a fungal infection and sarcoidosis have low levels of IL-10 within the serum resulting from granuloma formation which blocks IL-10 secretion [80]. Thus, a high level of IL-10 in a host’s serum may inhibit the development of fungal infections.
IL-12 is cytokine with a role in regulating T cell response to infection. IL-12 is secreted by monocytes/macrophages, neutrophils, and also other cells [81]. IL-12 is needed for the growth of Th1 cells. The main function of IL-12 is to induce IFN- γ production from NK cells and T cells. Another function is to increase cytotoxicity of NK cells, CTL, and to differentiate naïve T cells into Th1-cells [81,82]. IL-12 has an important role in regulating T cell responses, initiating immune cells to differentiate into Th1, Th17, and Treg cells [83], and maintaining acquired immunity to microorganism infections [84]. It has been reported that IL-12 plays an important role in humoral and adaptive immune response to fungal infection. Mice with an IL-12p40 deficiency experienced systemic infection after oral infection with fungus; this study also showed that IL-12 is important in preventing the spread of the fungal infection within the host body [85]. Consequently, the high level of IL-12 in the host serum might be crucial for inhibiting the dissemination of the fungal/dermatophyte infection.
Conclusion
This review highlights the important factors that affect dermatophyte infection in companion animals. Susceptibility to dermatophyte infection is dependent on animal age, sex, and species. The host immune system and virulence factor from the agent are also crucial for disease progression. Virulence factors from dermatophytes, namely, glycopeptide and keratinase, are responsible for initiating the host immune response. However, released stress hormone during dermatophyte infection has adverse effects on the host immune response. Stress hormone impairs the host immune response by downregulating or upregulating pro-inflammatory cytokines
Authors’ Contributions
SW and SI conceptualized the review, drafted the review, prepared, and edited the manuscript according to the title. SW and ADP collected the literature, edited the manuscript, and finalized the manuscript. All authors read and approved the final manuscript.
Acknowledgments
We thank the Directorate of Research and Community Service, General Directorate of Higher Education and Cultural Ministry, Indonesia, for funding Program Magister menuju Doktor untuk Sarjana Unggul (PMDSU) scholarship from KEMENRISTEK DIKTI NO. 6304/UNI/DITLIT/DIT-LIT/LT/2019 BATCH IV.
Competing Interests
The authors declare that they have no competing interests.
Publisher’s Note
Veterinary World remains neutral with regard to jurisdictional claims in published institutional affiliation.
References
- 1.Indarjulianto S, Yanuartono, Widyarini S, Raharjo S, Purnamaningsih H, Nururrozi A, Hariwibowo N, Jainudin H.A. Microsporum canis infection in dermatitis cats. J. Vet. 2017;18(2):207–210. [Google Scholar]
- 2.Ivaskiene M, Matusevicius A.P, Grigonis A, Zamokas G, Babickaite L. Efficacy of topical therapy with newly developed terbinafine and econazole formulations in the treatment of dermatophytosis in cats. Pol. J. Vet. Sci. 2016;19(3):535–543. doi: 10.1515/pjvs-2016-0067. [DOI] [PubMed] [Google Scholar]
- 3.Haggag Y.N, Samaha H.A, Nossair M.A, Mohammad R.M.H. Prevalence of dermatophytosis in some animals and human in Bahera Province, Egypt. Alex. J. Vet. Sci. 2017;53(2):64–71. [Google Scholar]
- 4.Tater K.C. An approach to pruritus. In: Jackson H, Marsella R, editors. Manual of Canine and Feline Dermatology. 3rd ed. England: British Small Animal Veterinary Association; 2015. pp. 41–42. [Google Scholar]
- 5.Rashidian S, Falahati M, Kordbacheh P, Mahmoudi M, Safara M, Sadeghi T.H, Mahmoudi S, Zaini F. A study on etiologic agents and clinical manifestations of dermatophytosis in Yazd, Iran. Curr. Med. Mycol. 2015;1(4):20–25. doi: 10.18869/acadpub.cmm.1.4.20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Frymus T, Gruffydd-Jones T, Pennisi M.G, Addie D, Belak S, Boucraut-Baralon C, Egberink H, Hartmann K, Hosie M.J, Lloret A, Lutz H, Marsilio F, Mostl K, Radford A.D, Thiry E, Truyen U, Hornizek M.C.D. Dermatophytosis in cats:ABDC guidelines on prevention and management. J. Feline Med. Surg. 2013;15(7):598–604. doi: 10.1177/1098612X13489222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Indarjulianto S, Yanuartono Purnamaningsih H, Wikansari P, Sakan G.Y.I. Isolation and identification of Microsporum canis from dermatophytosis dogs in Yogyakarta. J. Vet. 2014;15(2):212–216. [Google Scholar]
- 8.Galluppi R, Leveque J.F.C, Beghelli V, Bonoli C, Mattioli M, Ostanello F, Tampieri M.P, Accorsi P.A. Cortisol levels in cats, hair in presence or absence of Microsporum canis infection. Res. Vet. Sci. 2013;95(3):1076–1080. doi: 10.1016/j.rvsc.2013.07.023. [DOI] [PubMed] [Google Scholar]
- 9.Murmu S, Debnath C, Pramanik A.K, Mitra T, Jana S, Dey S, Banerjee S, Batabyal K. Detection and characterization of zoonotic dermatophytes from dogs and cats in and around Kolkata. Vet. World. 2015;8(9):1078–1082. doi: 10.14202/vetworld.2015.1078-1082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Chaitanya V.S, Lavania M, Nigam A, Turankar R.P, Singh I, Horo I, Sengupta U, Jadhava R.S. Cortisol and pro-inflammatory cytokine profiles in Type 1 (reversal) reactions of leprosy. Immunol. Lett. 2013;156(1-2):159–167. doi: 10.1016/j.imlet.2013.10.008. [DOI] [PubMed] [Google Scholar]
- 11.Pin D. Non-dermatophyte dermatoses mimicking dermatophytoses in animals. Mycopathologia. 2017;182(1-2):113–126. doi: 10.1007/s11046-016-0090-8. [DOI] [PubMed] [Google Scholar]
- 12.Azrad M, Freidus V, Kassem R, Peretz A. Identification of dermatophytes by MALDI-TOF MS technology in the clinical laboratory. Int. J. Mass. Spectrom. 2019;440(2019):32–36. [Google Scholar]
- 13.Dalis J.S, Kazeem H.M, Kwaga J.K.P, Kwanashie C.N. Prevalence and distribution of dermatophytosis lesions on cattle in Plateau State, Nigeria. Vet. World. 2019;12(9):1484–1490. doi: 10.14202/vetworld.2019.1484-1490. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Quinn P.J, Markey B.K, Carter M.E, Donelly W.J, Leonard F.C. Blackwell Science. United States: 2004. Veterinary Microbiology and Microbial Disease; pp. 224–226. [Google Scholar]
- 15.Fehr M. Zoonotic potential of dermatophytosis in small mammals. J. Exot. Pet Med. 2015;24(3):308–316. [Google Scholar]
- 16.Bernardo F, Guerra A, Martins H. Dermatophytes isolated from pet, dogs and cats, in Lisbon, Portugal (2000-2004) Rev. Port. Cienc. Vet. 2005;100(553-554):85–88. [Google Scholar]
- 17.Almuzaini A.M, Osman S.A, Saeed E.M.A. An outbreak of dermatophytosis in camels (Camelus dromedarius) at Qassim region, central of Saudi Arabia. J. Appl. Anim. Res. 2016;44(1):126–129. [Google Scholar]
- 18.Pinter L, Stritof Z. A retrospective study of Trichophyton mentagrophytes infection in dogs (1970-2002) Vet. Arhiv. 2004;74(4):251–260. [Google Scholar]
- 19.Seker E, Dogan N. Isolation of dermatophytes from dogs and cats with suspected dermatophytosis in Western Turkey. Prev. Vet. Med. 2011;98(1):46–51. doi: 10.1016/j.prevetmed.2010.11.003. [DOI] [PubMed] [Google Scholar]
- 20.Debnath C, Mitra T, Kumar A, Samanta I. Detection of dermatophytes in healthy companion dogs and cats in eastern India. Iran. J. Vet. Res. 2015;17(1):20–24. [PMC free article] [PubMed] [Google Scholar]
- 21.Roshanzamir H, Naserli S, Ziaie B, Fakour M. Incidence of dermatophytes isolated from dogs and cats in the city of Baku, Azerbaijan. Comp. Clin. Pathol. 2016;25(2):327–329. [Google Scholar]
- 22.Ahdy A.M, Sayed-Ahmed M.Z, Younis E.E, Baraka H.N, El-Khodery S.A. Prevalence and potential risk factors of dermatophytosis in Arabian horses in Egypt. J. Equine Vet. Sci. 2016;37(1):71–76. [Google Scholar]
- 23.Al-Ani F.K, Younes F.A, Al-Rawashdeh O.F. Ringworm infection in cattle and horses in Jordan. Act. Vet. Brno. 2002;71(1):55–60. [Google Scholar]
- 24.Gangil R, Dutta P, Tripathi R, Singathia R, Lakhotia R.L. Incidence of dermatophytosis in canine cases presented at Apollo veterinary college, Rajashtan, India. Vet. World. 2012;5(11):682–684. [Google Scholar]
- 25.Ciesielska A, Staczek P. Selection and validation of reference genes for qRT-PCR analysis of gene expression in Microsporum canis growing under different adhesion-inducing conditions. Sci. Rep. 2018;8(1197):1–10. doi: 10.1038/s41598-018-19680-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Moriello K.A. Zoonotic skin disease of dogs and cats. Anim. Health Res. Rev. 2003;4(2):157–168. doi: 10.1079/ahr200355. [DOI] [PubMed] [Google Scholar]
- 27.Cunha M.M.D, Capote-Bonato F, Capoci I.R.G, Bonato D.V, Ghizzi L.G, Paiva-Lima P, Baeza L.C, Svidzinski T.I.E. Epidemiological investigation and molecular typing of dermatophytosis caused by Microsporum canis in dogs and cats. Prev. Vet. Med. 2019;167(2019):39–45. doi: 10.1016/j.prevetmed.2019.03.019. [DOI] [PubMed] [Google Scholar]
- 28.Sakuragi Y, Sawada Y, Hara Y, Ohmori S, Omoto D, Haruyama S, Yoshioka M, Nishio D, Nakamura M. Increased circulating Th17 cell in a patient with tinea capitis caused by Microsporum canis. Allergol. Int. 2016;65(2):215–216. doi: 10.1016/j.alit.2015.10.003. [DOI] [PubMed] [Google Scholar]
- 29.Aktas E, Yigit N. Hemolytic activity of dermatophytes species isolated from clinical specimens. J. Mycol. Med. 2015;25(1):25–30. doi: 10.1016/j.mycmed.2014.10.014. [DOI] [PubMed] [Google Scholar]
- 30.Moretti A, Agnetti F, Mancianti F, Nardoni S, Righi C, Moretta I, Morganti G, Papini M. Dermatophytosis in animals:Epidemiological, clinical and zoonotic aspects. J. Ital. Dermatol. Venereol. 2013;148(6):563–572. [PubMed] [Google Scholar]
- 31.Putriningsih P.A.S, Widyastuti S.K, Arjentinia I.P.G, Batan I.W. Identification and prevalence of ringworm in Balinese cow. J. Vet. 2016;17(1):1–6. [Google Scholar]
- 32.Ilhan Z, Karaca M, Ekin I.H, Solmaz H, Akkan H.A, Tutuncu M. Detection of seasonal asymptomatic dermatophytes in Van cats. Braz. J. Microbiol. 2016;47(1):225–230. doi: 10.1016/j.bjm.2015.11.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Monod M. Secreted proteases from dermatophytes. Mycopathologia. 2008;166(5-6):285–294. doi: 10.1007/s11046-008-9105-4. [DOI] [PubMed] [Google Scholar]
- 34.Kurniati C.R. Etiopathogenesis of dermatophytoses. BIKKK. 2008;20(3):243–250. [Google Scholar]
- 35.Vermout S, Tabart J, Baldo A, Mathy A, Losson B, Mignon B. Pathogenesis of dermatophytosis. Mycopathologia. 2008;166(5-6):267–275. doi: 10.1007/s11046-008-9104-5. [DOI] [PubMed] [Google Scholar]
- 36.Peres N.T.D, Rossi A, Maranho F.C.A, Rossi N.M.M. Dermatophytes:Host-pathogen interaction and antifungal resistance. An. Bras. Dermatol. 2010;10(5):657–667. doi: 10.1590/s0365-05962010000500009. [DOI] [PubMed] [Google Scholar]
- 37.Reis A.P.C, Correia F.F, Jesus T.M, Pagliari C, Sakai-Valente N.Y, Junior W.B, Criado P.R, Benard G, Sousa M.G.T. In situ immune response in human dermatophytosis:Possible role of Langerhans cells (CD1a+) as risk factor for dermatophyte infection. Rev. Inst. Med. Trop. Sao Paulo. 2019;61(e56):1–6. doi: 10.1590/S1678-9946201961056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Laksmipathy D.T, Kannabiran K. Review on dermatomycosis:Pathogenesis and treatment. Nat. Sci. 2010;2(7):726–731. [Google Scholar]
- 39.Almeida S.R. Immunology of dermatophytosis. Mycopathologia. 2008;277(5-6):277–283. doi: 10.1007/s11046-008-9103-6. [DOI] [PubMed] [Google Scholar]
- 40.Romani L. Reviews:Immunity to fungal infections. Nat. Rev. Immunol. 2004;4(1):1–13. doi: 10.1038/nri1255. [DOI] [PubMed] [Google Scholar]
- 41.Hernandez-Chavez M.J, Perez-Garcia L.A, Nino-Vega G.A, Mora-Montes H.M. Review:Fungal strategies to evade the host immune recognition. J. Fungi. 2017;3(51):1–28. doi: 10.3390/jof3040051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Aristotle Immunity to infection. Primer to the Immune Response. 2nd ed. Netherlands: Ch. 13. Elsevier; 2014. pp. 295–332. [Google Scholar]
- 43.Thomas D.S, Ingham E, Bojar R.A, Holland K.T. In vitro modulation of human keratinocyte pro-and anti-inflammatory cytokine production by the capsule of Malassezia species. FEMS Immunol. Med. Microbiol. 2008;54(2):203–214. doi: 10.1111/j.1574-695X.2008.00468.x. [DOI] [PubMed] [Google Scholar]
- 44.Mambula S.S, Sau K, Henneke P, Golenbock D.T, Levitz S.M. Toll-like receptor (TLR) signaling in response to Aspergillus fumigatus. J. Biol. Chem. 2002;277(42):39320–39326. doi: 10.1074/jbc.M201683200. [DOI] [PubMed] [Google Scholar]
- 45.Netea M.G, Van Der Graff C.A, Vonk A.G, Verschueren I, Van Der Meer J.W, Kullberg B.J. The role of toll-like receptor (TLR) 2 and TLR4 in the host defense against disseminated candidiasis. J. Infect. Dis. 2002;185(10):1483–9. doi: 10.1086/340511. [DOI] [PubMed] [Google Scholar]
- 46.Gantner B.N, Simmons R.M, Canavera S.J, Akira S, Underhill D.M. Collaborative induction of inflammatory responses by dectin-1 and toll-like receptor 2. J. Exp. Med. 2003;197(9):1107–1117. doi: 10.1084/jem.20021787. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Richmond J.M, Harris J.E. Immunology and skin in health and disease. Cold Spring Harb. Perspect. Med. 2014;4(12):1–20. doi: 10.1101/cshperspect.a015339. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Blanco J.L, Garcia M.E. Immune response to fungal infections. Vet. Immunol. Immunopathol. 2008;125(1-2):47–70. doi: 10.1016/j.vetimm.2008.04.020. [DOI] [PubMed] [Google Scholar]
- 49.García-Romero M.T, Arenas R. New insights into genes, immunity, and the occurrence of dermatophytosis. J. Invest. Dermatol. 2015;135(3):655–657. doi: 10.1038/jid.2014.498. [DOI] [PubMed] [Google Scholar]
- 50.Tian R, Hou G, Li D, Yuan T.F. A possible change process of inflammatory cytokines in the prolonged chronic stress and its ultimate implications for health. Sci. World J. 2014;2014 doi: 10.1155/2014/780616. Article ID 780616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Padgett D.A, Glaser R. How stress influences the immune response. Trends Immunol. 2003;24(8):444–448. doi: 10.1016/s1471-4906(03)00173-x. [DOI] [PubMed] [Google Scholar]
- 52.Lionakis M.S, Kontoyiannis D.P. Glucocorticoids and invasive fungal infections. Lancet. 2003;362(9398):1828–1838. doi: 10.1016/S0140-6736(03)14904-5. [DOI] [PubMed] [Google Scholar]
- 53.Janicki-Deverts D, Cohen S, Turner R.B, Doyle W.J. Basal salivary cortisol secretion and susceptibility to upper respiratory infection. Brain Behav. Immun. 2016;53(2016):255–261. doi: 10.1016/j.bbi.2016.01.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Elenkov I.J, Chrousos G.P. Pro/anti-inflammatory cytokines and susceptibility to disease. Trend. Endocrinol. Metab. 1999;10(9):359–368. doi: 10.1016/s1043-2760(99)00188-5. [DOI] [PubMed] [Google Scholar]
- 55.Baker M.R, Swanson P, Young G. Injuries from non-retention in gillnet fisheries suppress reproductive maturation in escaped fish. PLoS One. 2013;8(7):1–15. doi: 10.1371/journal.pone.0069615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Widyarini S, Spinks N, Husbands A.J, Reeve V.E. Isoflavonoid compounds from red clover (Trifolium pratense) protect from inflammation and immune suppression induced by UV radiation. Photochem. Photobiol. 2001;74(3):465–470. doi: 10.1562/0031-8655(2001)074<0465:icfrct>2.0.co;2. [DOI] [PubMed] [Google Scholar]
- 57.Bandara M.A, Allanson S.J, Widyarini S, Chai Z, Reeve V.E. Topical isoflavonoids reduce experimental cutaneous inflammation in mice. Immunol. Cell Biol. 2010;88(7):727–733. doi: 10.1038/icb.2010.26. [DOI] [PubMed] [Google Scholar]
- 58.Abdulkhaleq L.A, Assi M.A, Abdullah R, Zamri-saad M, Taufiq-Yap Y.H, Hezmee M.N.M. The crucial roles of inflammatory mediators in inflammation:A review. Vet. World. 2018;11(5):627–635. doi: 10.14202/vetworld.2018.627-635. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Antachopulus C, Waalsh T.J, Roilides E. Fungal infections in primary immunodeficiencies. Eur. J. Pediatr. 2007;166(11):1099–1117. doi: 10.1007/s00431-007-0527-7. [DOI] [PubMed] [Google Scholar]
- 60.Guo B, Lager K.M, Henningson J.N, Miller L.C, Schlink S.N, Kappes M.A, Faaberg K.S. Experimental infection of United States swine with a Chinese highly pathogenic strain of porcine reproductive and respiratory syndrome virus. Virology. 2013;435(2):372–384. doi: 10.1016/j.virol.2012.09.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Awasthi A, Kuchroo V.K. Th17 cells:From precursors to players in inflammation and infection. Int. Immunol. 2009;21(5):489–498. doi: 10.1093/intimm/dxp021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Isailovic N, Daigo K, Mantovani A, Selmi C. Interleukin-17 and innate immunity in infections and chronic inflammation. J. Autoimmun. 2015;60(2015):1–11. doi: 10.1016/j.jaut.2015.04.006. [DOI] [PubMed] [Google Scholar]
- 63.Huffnagle G.B, Deepe G.S. Innate and adaptive determinants of host susceptibility to medically important fungi. Curr. Opin. Microbiol. 2003;6(4):344–350. doi: 10.1016/s1369-5274(03)00089-4. [DOI] [PubMed] [Google Scholar]
- 64.Sha Y, Markovic-Plese S. A role of IL-1R1 signaling in the differentiation of Th17 cells and development of autoimmune diseases. Self. Nonself. 2011;2(1):35–42. doi: 10.4161/self.2.1.15639. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Burstein V.L, Guasconi L, Beccacece I, Theumer M.G, Mena C, Prinz I, Cervi L, Herrero M, Masih D.T, Chiapello L.S. IL-17-mediated immunity controls skin infection and T helper 1 response during experimental Microsporum canis dermatophytosis. J. Invest. Dermatol. 2018;138(8):1744–1753. doi: 10.1016/j.jid.2018.02.042. [DOI] [PubMed] [Google Scholar]
- 66.Sparber F, LeibundGut-Landmann S. IL-17 takes center stage in dermatophytosis. J. Invest. Dermatol. 2018;138(8):1691–1693. doi: 10.1016/j.jid.2018.03.1518. [DOI] [PubMed] [Google Scholar]
- 67.Sparber F, LeibundGut-Landmann S. Interleukin 17-mediated host defense against Candida albicans. Pathogens. 2015;4(3):606–619. doi: 10.3390/pathogens4030606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Conti P, Tettamanti L, Mastrangelo F, Ronconi G, Frydas I, Kritas S.K, Caraffa A, Pandofi F. Impact of fungi on immune responses. Clin. Ther. 2018;4(6):885–888. doi: 10.1016/j.clinthera.2018.04.010. [DOI] [PubMed] [Google Scholar]
- 69.Schett G, Dayer J.M, Manger B. Interleukin-1 function and role in rheumatic disease. Nat. Rev. Rheumatol. 2016;12(1):14–24. doi: 10.1038/nrrheum.2016.166. [DOI] [PubMed] [Google Scholar]
- 70.Gresnigt M.S, Van de Veerdonk F.L. The role of interleukin-1 family members in the host defence against Aspergillus fumigatus. Mycopathologia. 2014;178(5-6):395–401. doi: 10.1007/s11046-014-9776-y. [DOI] [PubMed] [Google Scholar]
- 71.Smith N.L.D, Hankinson J, Simpson A, Bowyer P, Denning D.W. A prominent role for the IL1 pathway and IL15 in susceptibility to chronic cavitary pulmonary aspergillosis. Clin. Microbiol. Infect. 2014;20(8):480–8. doi: 10.1111/1469-0691.12473. [DOI] [PubMed] [Google Scholar]
- 72.Rodriguez-Cerdeira C, Carnero-Gregorio M, Lopez-Barcenas A, Fabbrocini G, Sanchez-Blanco E, Alba-Menendez A, Guzman R.A. Interleukin-2 and other cytokines in candidiasis:Expression, clinical significance, and future therapeutic targets. Acta. Dermatovenerol. Alp. Pannonica. Adriatic. 2018;27(2):91–102. [PubMed] [Google Scholar]
- 73.Sharma R, Fu S.M, Ju S.T. IL-2:A two-faced master regulator of autoimmunity. J. Autoimmun. 2011;36(2):91–97. doi: 10.1016/j.jaut.2011.01.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Zelante T, Fric J, Wong A.Y.W, Ricciardi-Castagnoli P. Interleukin-2 production by dendritic cells and its immuno-regulatory functions. Front. Immunol. 2012;3(161):1–6. doi: 10.3389/fimmu.2012.00161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Khan D.A, Ansari W.M, Khan F.A. Pro/anti-inflammatory cytokines in the pathogenesis of premature coronary artery disease. J. Int. Cytokine Res. 2011;31(7):561–567. doi: 10.1089/jir.2010.0157. [DOI] [PubMed] [Google Scholar]
- 76.Schneider C.P, Schwacha M.G, Chaudry I.H. The role of interleukin-10 in the regulation of the systemic inflammatory response following trauma-hemorrhage. Biochim. Biophys. Acta. 2004;24(1):22–32. doi: 10.1016/j.bbadis.2004.01.003. [DOI] [PubMed] [Google Scholar]
- 77.Cyktor J.C, Turner J. Interleukin-10 and immunity against prokaryotic and eukaryotic intracellular pathogens. Infect. Immun. 2011;79(8):2964–2973. doi: 10.1128/IAI.00047-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Roussey J.A, Olszewski M.A, Osterholzer J.J. Immunoregulation in fungal diseases. Microorganisms. 2016;4(47):1–22. doi: 10.3390/microorganisms4040047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Netea M.G, Sutmuller R, Hermann C, Van der Graff C.A, Van Krieken J.H, Hartug T, Adema G, Kullberg B.J. Toll-like receptor 2 suppresses immunity against Candida albicans through induction of IL-10 and regulatory T cells. J. Immunol. 2004;172(6):3712–8. doi: 10.4049/jimmunol.172.6.3712. [DOI] [PubMed] [Google Scholar]
- 80.Tercelj M, Stopinsek S, Ihan A, Salobir B, Simcic S, Rylander R. Fungal exposure and low levels of IL-10 in patients with sarcoidosis. Pulm. Med. 2014;2014 doi: 10.1155/2014/164565. Article ID 164565. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Gee K, Guzzo C, Che Mat N, Ma W, Kumar A. The IL-12 family of cytokines in infection, inflammation and autoimmune disorders. Inflamm. Allergy Drug Targets. 2009;8(1):40–52. doi: 10.2174/187152809787582507. [DOI] [PubMed] [Google Scholar]
- 82.Sun L, He C, Nair L, Yeung J, Egwuagu C.E. Interleukin 12 (IL-12) family cytokines:Role in immune pathogenesis and treatment of CNS autoimmune disease. Cytokine. 2015;75(2):249–255. doi: 10.1016/j.cyto.2015.01.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Thompson A, Orr S.J. Emerging IL-12 family cytokines in the fight against fungal infections. Cytokine. 2018;111(2018):398–407. doi: 10.1016/j.cyto.2018.05.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Romani L, Puccetti P, Bistoni F. Interleukin-12 in infectious diseases. Clin. Microbiol. Rev. 1997;10(4):611–636. doi: 10.1128/cmr.10.4.611. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Conti H.R, Shen F, Nayyar N, Stocum E, Sun J.N, Lindemann M.J, Ho A.W, Hai J.H, Yu J.J, Jung J.W, Filler S.G, Masso-Welch P, Edgerton M, Gaffen S.L. Th17 cells and IL-17 receptor signaling are essential for mucosal host defense against oral candidiasis. J. Exp. Med. 2009;206(2):299–311. doi: 10.1084/jem.20081463. [DOI] [PMC free article] [PubMed] [Google Scholar]