
Fig. 2. Dorsoventral polarity determines the efficiency of cell migration in confinement.
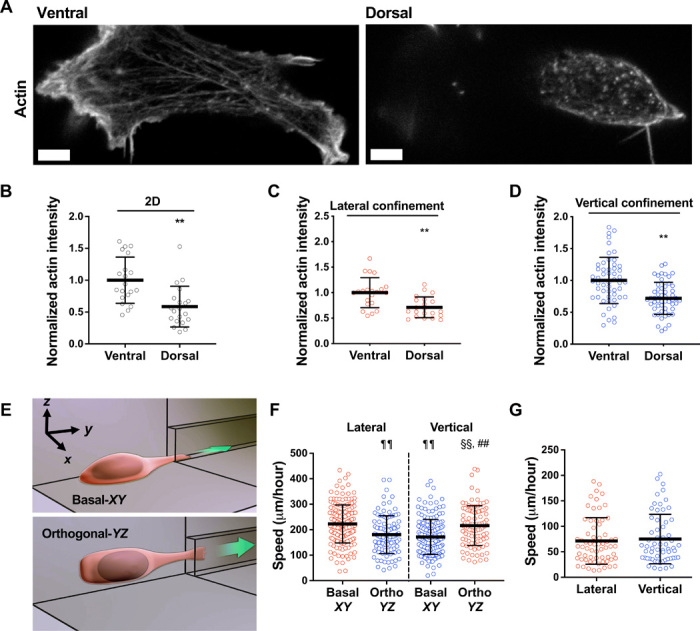
(A) Representative images of HT-1080 cell stained for actin on its ventral and dorsal surfaces on 2D. Scale bars, 5 μm. (B to D) Actin intensity on the ventral and dorsal surfaces of cells fixed and stained with actin phalloidin on 2D (n = 20 cells; two independent experiments) (B) or in lateral (n = 21 cells; two independent experiments) (C) and vertical (n = 53 cells; three or more independent experiments) (D) microchannels. Actin intensity was normalized to the ventral layer. (E) Schematic representation of a microfluidic device in which the 2D cell seeding area is orthogonal (YZ plane) to the typically used basal (XY plane) seeding region. (F) Migration speeds of HT-1080 cells in vertical and lateral microchannels, as assessed after seeding cells on a basal (XY) or orthogonal (YZ) seeding region (n ≥ 96 cells; four independent experiments). (G) Migration speeds of HT-1080 cells in lateral and vertical microchannels treated with poly-l-lysine (PLL) and methoxy poly(ethylene glycol) (mPEG) succinimidyl valerate (n ≥ 61 cells; three independent experiments). Data represent the mean ± SD; **P < 0.01 relative to ventral; ##P < 0.01 relative to vertical basal-XY; ¶¶P < 0.01 relative to lateral basal-XY; §§P < 0.01 relative to lateral orthogonal-YZ.