
Fig. 1. Analysis of early steps in the evolution of P. aeruginosa wild-type strain and antibiotic-resistant mutants in the presence of CAZ.
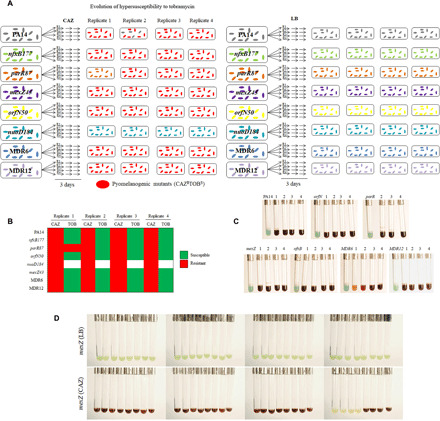
(A) Scheme of the resultant phenotype after the evolution of PA14, single (nfxB177, parR87, mexZ43, orfN50, and nuoD184) and multiple (MDR6 and MDR12) mutants, in the presence of CAZ [left part of (A)] or in the absence of antibiotic [LB; right part of (A)] for 3 days (see Materials and Methods). Pyomelanin hyperproduction was observed in 27 of 32 populations evolved in the presence of CAZ (red-colored cells). (B) Diagram showing convergence toward hypersusceptibility to TOB in the different genetic backgrounds and replicates, analyzed after short-term evolution on CAZ. In all cases, acquisition of a pyomelanogenic phenotype is associated with collateral sensitivity to TOB, irrespective of the genetic background of the evolving strain. MIC values of TOB and CAZ are included in table S2. (C) Isolation of pyomelanogenic clones from each 27 pyomelanogenic population [left part of (A); red-colored cells] obtained in the presence of CAZ. As shown in the figure, the early steps of evolution in presence of CAZ of P. aeruginosa, which lead to collateral sensitivity to TOB and pyomelanin production, are conserved among the different antibiotic-resistant mutants analyzed. (D) Pyomelanogenic phenotype of mexZ43 mutant after 1-day evolution in the presence of CAZ or in the absence of antibiotic (LB). Pyomelanin hyperproduction was observed in 28 of 32 populations evolved on CAZ. Photo credits for (C) and (D): Inés Poveda, Centro Nacional de Biotecnología. Permission for using these images is not required.