

Abstract
Purpose of review:
Chronic consumption of fructose and fructose-containing sugars leads to dyslipidemia. Apolipoprotein (apo) CIII is strongly associated with elevated levels of triglycerides and cardiovascular disease risk. We reviewed the effects of fructose consumption on apoCIII levels and the role of apoCIII in fructose-induced dyslipidemia.
Recent findings:
Consumption of fructose increases circulating apoCIII levels compared with glucose. The more marked effects of fructose compared to glucose on apoCIII concentrations may involve the failure of fructose consumption to stimulate insulin secretion. The increase in apoCIII levels after fructose consumption correlates with increased postprandial serum triglyceride (TG). Further, RNAinterference of apoCIII prevents fructose-induced dyslipidemia in non-human primates. Increases in postprandial apoCIII after fructose, but not glucose consumption, are positively associated with elevated TG in large TG-rich lipoproteins and increased small dense LDL levels.
Summary:
ApoCIII might be causal in the lipid dysregulation observed after consumption of fructose and fructose-containing sugars. Decreased consumption of fructose and fructose-containing sugars could be an effective strategy for reducing circulating apoCIII and subsequently lowering TG levels.
Keywords: Apolipoprotein CIII, fructose, dyslipidemia
Introduction:
Numerous population studies demonstrate that consumption of fructose-containing sugars is associated with the development and progression of metabolic diseases, including type 2 diabetes and cardiovascular disease (CVD). Both animal and clinical studies provide evidence to suggest that the relationship between fructose and cardiometabolic disease is causal [1*]. However, as the metabolic effects of fructose consumption are numerous and extensive, a clear picture of the involved pathways and their relative contributions will require much more investigation.
Apolipoprotein (apo) CIII is mainly found in triglyceride (TG)-rich lipoproteins (TRL) and HDL particles. Epidemiological studies show an association of serum apoCIII levels with CVD [2]. These observations were corroborated by findings that loss-of-function mutations in the apoCIII gene seem protective [3,4] and gain-of-function mutations predispose [5] for CVD. These results have targeted apoCIII for drug development to treat dyslipidemia and early results using apoCIII-lowering drugs seem promising [6].
Text of review:
Data on the effects of fructose consumption on circulating levels of apoCIII is limited but convincing. In non-human primates, fructose feeding over 1, 3 and 12 months increased circulating apoCIII levels [7,8**]. In our intervention study, circulating apoCIII increased in a dose-dependent manner in healthy young adults who consumed 0%, 10%, 17.5% or 25% of their daily energy requirement as high fructose corn syrup (HFCS)-sweetened beverages over two weeks [9]. Taskinen et al. reported increased apoCIII levels in abdominally obese subjects consuming 75g of fructose/day over 12 weeks [10**]. And finally, we recently published data from a 10-week intervention in which fasting and postprandial apoCIII levels were increased in older and overweight subjects consuming 25% energy requirement as fructose compared with those consuming glucose [11**]. These data are corroborated by a 9-day feeding trial in children with obesity and metabolic syndrome whose usual diet contained 27% of daily energy as added sugar [12,13]. Providing the children with low sugar diets (10% of daily energy) decreased circulating apoCIII [12].
Fructose consumption increases apoCIII expression
ApoCIII expression is regulated through various pathways many of which involve dietary components such as fatty acids or carbohydrates. Glucose increases the expression of apoCIII through transcriptional activation of hepatocyte nuclear factor 4 alpha in combination with either carbohydrate-responsive element-binding protein 1 (ChREBP-1) [14] or sterol-regulatory-element-binding protein-1 (SREBP-1) [15]. Caron et al. speculated that this activation by glucose lowers lipolysis via lipoprotein lipase (LPL) inhibition (see below). This decreases the uptake of fatty acids in peripheral tissues, thereby promoting glucose over fatty acid metabolism [14]. However, the relationship between glucose and apoCIII is likely to be more complex than that [16] because insulin action causes downregulation of apoCIII expression via Forkhead Box O1 (FoxO1) [17]. FoxO1 is a transcription factor that resides in the nucleus. It activates hepatic apoCIII expression [17]. Upon insulin signaling, FoxO1 is phosphorylated via the PI3K/ protein kinase B (PKB/Akt) cascade. This phosphorylation causes the nuclear export and degradation of FoxO1 and hence the induction of apoCIII expression is ceased. The timing and the concentrations of glucose versus insulin that balance out their opposing effects on apoCIII expression have yet to be delineated [16]. Possibly the upregulation of apoCIII by glucose is transitory and quickly reversed by the increase in circulating insulin that follows the blood glucose spikes. Accordingly, individuals with insulin resistance or poorly managed type 1 diabetes mellitus exhibit high levels of apoCIII [18–21]. Thus failure of insulin to counter-balance the glucose-induced increase of apoCIII expression appears to be detrimental.
Fructose consumption also results in activation of the transcription factors ChREBP-1 [22,23] and SREBP-1 [24]. In rat livers, fructose-induced activation of both transcription factors exceeded the activation induced by glucose [25]. However, in contrast to glucose, fructose consumption does not stimulate insulin release [26]. Therefore the increase in apoCIII expression following fructose consumption is not counterbalanced with inhibition by insulin (Figure 1). In our study with older and overweight subjects, fructose consumption increased apoCIII levels in both the fasting and postprandial state compared with glucose consumption [11**].
Figure 1.
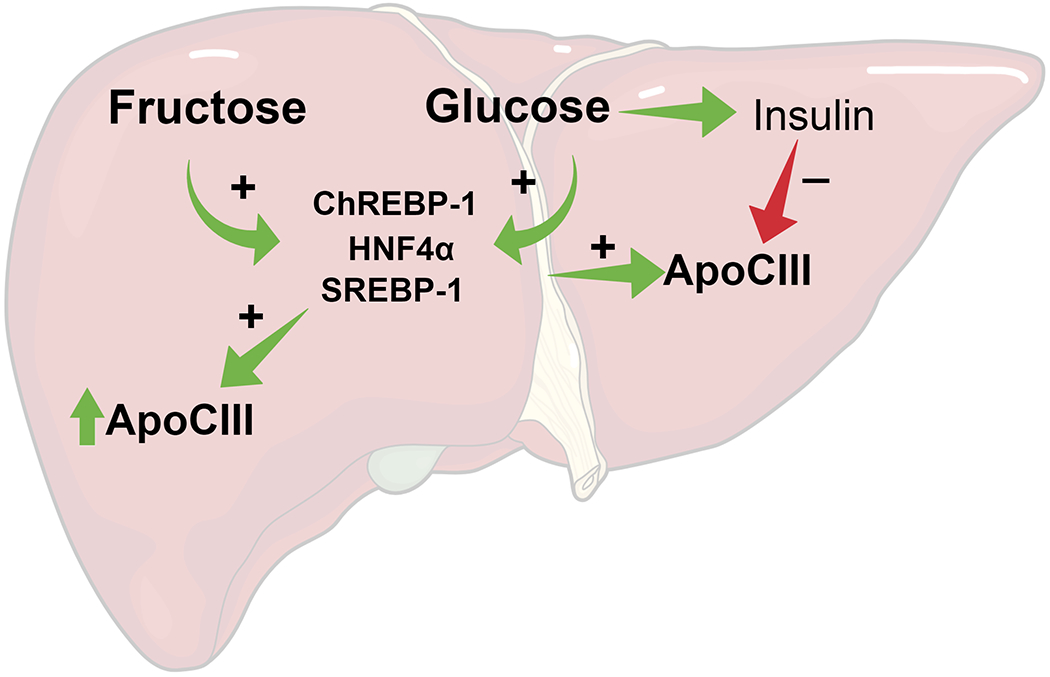
Effects of Fructose and glucose consumption on apoCIII expression
Consumption of glucose and fructose lead hepatic activation of ChREBP-1 and SREBP-1. The combined action of the individual transcription factors with HNF4α lead to increased apoCIII expression. The rise in glucose levels triggers insulin release from the pancreas. Insulin action results in the inhibition of apoCIII expression. Therefore, there is no change in apoCIII after glucose consumption but an increase after fructose.
ApoCIII contributes to fructose-induced dyslipidemia
While the evidence for a particularly detrimental role of fructose in the development of CVD and other undesirable metabolic health outcomes is growing, we still lack a firm understanding of the underlying mechanisms. Fructose consumption leads to acute and chronic changes in blood lipids. These changes include established features of dyslipidemia and the atherogenic lipoprotein phenotype – increased circulating TG and TG-rich lipoproteins (TRL), small dense LDL (sdLDL) and changes in HDL particle composition.
Triglycerides and triglyceride-rich lipoproteins
The most acute change in lipid profiles after fructose consumption is the increase in postprandial TG, which occurs during the first day of high fructose feeding [27–29]. Hepatic de novo lipogenesis (DNL) is one mechanism by which fructose consumption leads to increased TG levels [30]. However, Taskinen et al. constructed a seven-variable multivariate regression model that explained 73.7 % of the variance in TG levels after fructose consumption [10**]. DNL accounted for only 16% of the variance. The predominant contributor to the increase in TG was apoCIII, which accounted for 59% of the variance.
We recently reported the TG and cholesterol content of 13 lipoprotein subfractions fractions separated by size in the plasma collected from the overweight/obese men and women who consumed fructose- or glucose-sweetened beverages for 10 weeks. The most marked increase induced by fructose consumption during the postprandial period was the TG in large TG-rich particles (TRL) [11**]. Interestingly, the increase in postprandial apoCIII was associated with postprandial TRL TG, but not with total postprandial TG (fructose-induced effect of ΔapoCIII: on Δtotal TRL TG, r: 0.49, p=0.006; on Δtotal TG, r: 0.178 p=0.349, unpublished data).
Three mechanisms have been proposed by which apoCIII contributes to increased TG levels. Kinetic studies with humans and genetically modified mice suggested that apoCIII promotes mobilization and utilization of triglyceride during VLDL1 assembly/secretion [31]. However, there are conflicting results from additional mice and human studies and more research is needed to confirm this mechanism [32,33*]. Downstream of VLDL assembly, there are two mechanism by which apoCIII promotes retention of TG in the circulation. ApoCIII in TRL inhibits LPL, the rate-limiting enzyme that hydrolyzes TG in circulating TRL to allow for fatty acid uptake by heart, muscle and adipose. ApoCIII also interferes with TRL clearance by masking hepatic LDL receptor ( LDLR) and LDL receptor like protein 1 (LPR-1 ) [34**]. ApoCIII’s effects on TRL retention may involve apoE. As proposed by Ramms et al. [34**], in the presence of apoE on the same particle as apoCIII, apoCIII interferes with hepatic clearance of TRL; while in the absence of apoE, apoCIII inhibits LPL activity [34**]. These results raise interesting questions regarding the role of apoCIII and apoE after fructose consumption. A carbohydrate-rich diet results in an increase in circulating VLDL and IDL particles lacking apoE [35]. However, in our study [9,36] the increases in apoCIII induced by HFCS or glucose consumption were accompanied by comparable increases in apoE, however the increases in apoCIII induced by fructose were not (unpublished data).
Results from drug trials targeting apoCIII with the aim to lower triglycerides are favorable. Volanesorsen, an apoCIII anti-sense oligonucleotide reduced TG levels decreased by 69% in a 15 week trial [6]. A substantial decrease in TG (57%) that was correlated with the decrease in apocIII (r = 0.91, p = 0.0015) was also achieved in a small cohort of middle-aged subjects with high liver fat who consumed a low carbohydrate diet (5% of energy) for 2 weeks [37]. However, restriction of dietary sugars to 10% of daily energy, via replacement with complex carbohydrate, led to a 46% decrease in TG and a 49% decrease in apoCIII levels in only 9 days [12]. It would be of interest to test apoCIII lowering drugs against a restriction in dietary sugars.
High levels of TRL are a gateway to other changes in the lipoprotein metabolism. In our previous study, we assessed the effects of HFCS dose on blood lipids/lipoproteins [9]. Non-HDL-C, LDL and apoB increased with the consumed dose of HFCS. Although the changes in TG and apoCIII were strongly correlated with each other, only the change in apoCIII – not the change in TG – was a predictor independent contributor to the dose-dependent effects of HFCS on non-HDL-C, LDL and apoB [9]. This suggests that the blood concentrations of these risk factors are more affected by apoCIII’s effects on retention than any direct effects of circulating TG.
Small dense LDL
We recently reported an association between the increased levels of circulating apoCIII and the increase in sdLDL in subjects consuming fructose [11**]. It has long been hypothesized that it is the increased and sustained high concentration of TRL in the circulation that contributes to the emergence of sdLDL [11**,38]. ApoCIII is likely a necessary factor in this pathway (Figure 2). When apoCIII inhibits clearance of TRL, they become the preferred substrate of cholesterol ester transfer protein (CETP), thus promoting transfer of TG from TRL into LDL particles. Our unpublished data show that postprandial TRL TG in subjects consuming fructose was strongly associated with the TG enrichment of LDL in the postprandial period. Hepatic lipase subsequently lyses the TG from TG-enriched LDL particles generating TG-depleted, cholesterol-rich sdLDL. As we reported [11**], after 10 weeks fructose consumption, the increases in postprandial TG in large TRL and the increases in postprandial apoCIII were both associated with the increased sdLDL-C at fasting [11**]. However, the individual effects of TG and apoCIII were attenuated when they were included in the same model, indicating that their effects were mediated by a dependent pathway [11**]. Likely, as shown in Figure 2, this dependent pathway is the apoCIII-induced retention of TRL that allows for TG-enrichment of LDL. Interestingly, the reduction of apoCIII in the children with obesity and metabolic syndrome, who consumed the low sugar diet for 9 days, was also accompanied by a decrease in sdLDL particles [12]. While these results do not offer mechanistic proof, they support the possibility that apoCIII, along with TRL TG, is mediating the changes in sdLDL.
Figure 2.
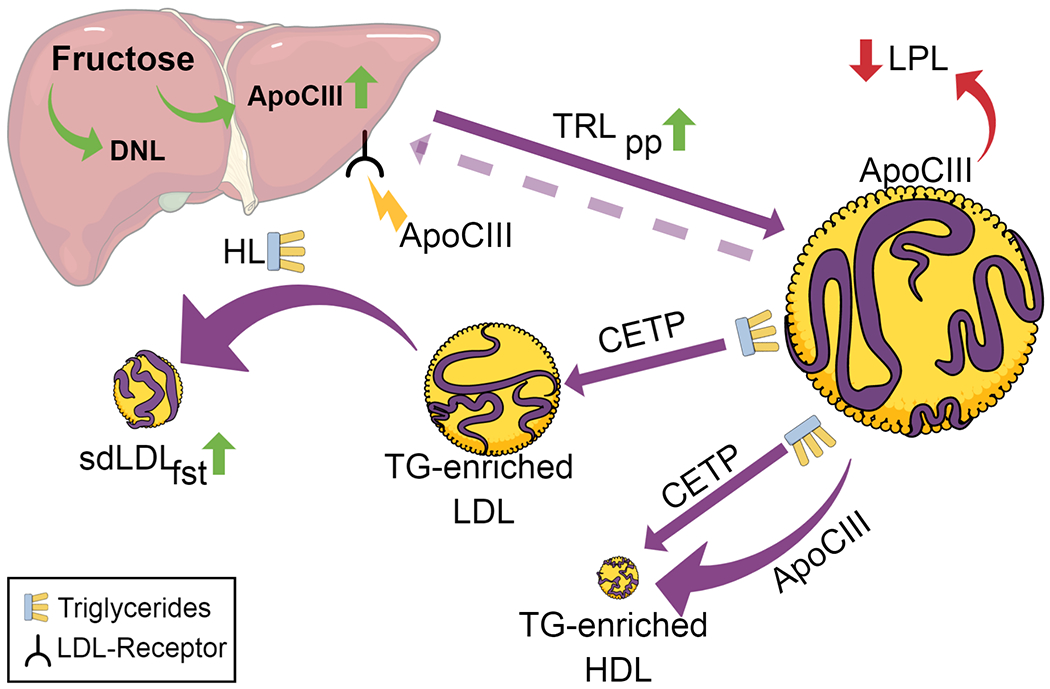
Fructose and ApoCIII on dyslipidemia – Proposed Mechanism
Hepatic fructose exposure leads to increased apoCIII expression and increased DNL, resulting in an increase in triglyceride rich particles containing apoCIII. ApoCIII blocks the clearance of TRL and lipoproteinlipase resulting in sustained high TRL in circulation. TRL then become the preferred substrate to CETP which transfers TG from TRL to LDL and HDL particles. Hepatic lipase lyses TG from TG-enriched LDL resulting in increased levels of sdLDL in the fasting state. TG-enriched HDL and HDL carrying apoCIII exhibit dysfunction like reduced reverse cholesterol transport.
Abbreviations: HL – hepatic lipase; fst – fasting; pp - postprandial
HDL
Dyslipidemia is further characterized by changes in HDL function. However, neither our studies nor clinical trials by other investigators [39] show that consumption of fructose-containing sugars decrease blood concentrations of HDL-cholesterol (HDL-C). Generally, HDL-C is considered to be the “good cholesterol”; however, recent research suggests that HDL-C is not always associated with favorable health outcomes [40,41]. While, higher HDL-C concentrations seem to protect against CVD in metabolically healthy individuals, HDL-C in individuals with unfavorable metabolic conditions associate with progression of metabolic diseases [42,43*]. The reason for these discrepant associations between HDL-C concentrations and CVD risk arise from the diversity of HDL composition, which defines the particles properties [41]. Data suggest that apoCIII on HDL particles impairs their cholesterol efflux capacity and thereby increases the risk for CVD [44,45]. In addition, the effects of apoCIII might extend to other HDL functions. HDL-containing apoCIII showed impaired anti-diabetogenic properties [46*]. We have not assessed apoCIII content in specific lipoprotein fractions in subjects with fructose-induced increases in apoCIII concentrations. However, since 10-weeks fructose consumption also decreased insulin sensitivity compared with glucose consumption [11**,47], assessing the apoCIII of the HDL subfractions would be of interest.
Individuals with hypertriglyceridemia commonly display TG-enrichment in HDL particles [48]. As shown in Figure 2, when blood concentration of TRL are high, CETP transfers TG from TRL into HDL particles [49], as well as LDL particles. These TG-enriched HDL particles exhibit decreased stability and decreased cholesterol efflux capacity [50]. We measured TG concentrations in HDL particles in the subjects consuming fructose or glucose for 10 weeks. In the postprandial stage, TG content was markedly increased in HDL fractions in subjects consuming fructose (+0.040 mmol/L (95% CI: 0.027 to 0.053 mmol/L), P=0.0015) compared to those consuming glucose (unpublished data).
These findings suggest that though fructose consumption does not decrease circulating HDL-C, it causes changes in HDL particle composition that are associated with unfavorable health outcomes. These effects might be related to apoCIII on HDL particles. However, it remains to be elucidated if dysfunctional HDL particles themselves contribute to the development of metabolic diseases or if they are just innocent bystander that reflect the overall metabolic status as proposed by Taskinen and Borén [51]. The effects of fructose consumption and increased apoCIII levels on HDL particles require further investigation.
Conclusion:
In summary, fructose consumption results in increased circulating apoCIII levels compared with glucose consumption. This difference is likely explained by the failure of fructose to induce insulin secretion, thereby allowing continued stimulation of apoCIII expression. Increased circulating apoCIII is associated with elevated triglyceride levels and other features of dyslipidemia, such as the generation of small dense LDL and TG enrichment of HDL. Future studies investigating consumption of fructose and fructose-containing sugars should include the assessment of apoCIII content, and preferably apoE content too, in individual lipoprotein fractions. Future studies should also compare the effectiveness of drug therapy versus dietary fructose restriction for decreasing apoCIII levels and other features of the atherogenic lipoprotein phenotype.
Key points:
Fructose consumption uniquely increases circulating apoCIII levels in humans and non-human primates
ApoCIII might be causal in the generation of sdLDL as seen after fructose consumption
Dietary fructose restriction might be an effective therapeutic measure to reduce apoCIII levels and associated undesirable health outcomes
Acknowledgements
Financial support and sponsorship
Havel/Stanhope clinical trials supported by the NIH/National Heart, Lung and Blood Institute 1R01HL075675 and 1R01HL09133 1R01 HL107256 and the National Center for Research Resources, a component of the NIH, and NIH Roadmap for Medical Research UL1RR024146. Bettina Hieronimus is supported by a research fellowship from the German Research Foundation (Deutsche Forschungsgemeinschaft) HI 2113/1-1.
Footnotes
Conflicts of interest
None
References
- *1.Taskinen M-R, Packard CJ, Borén J: Dietary fructose and the metabolic syndrome. Nutrients 2019, 11. [DOI] [PMC free article] [PubMed] [Google Scholar]; This review summarizes the current knowledge on the effects of excessive fructose consumption on the development and progression of metabolic diseases, with a particular emphasis on lipid metabolism and non-alcoholic fatty liver disease.
- 2.Colombo M, Looker HC, Farran B, Agakov F, Brosnan MJ, Welsh P, Sattar N, Livingstone S, Durrington PN, Betteridge DJ, et al. : Apolipoprotein CIII and N-terminal prohormone b-type natriuretic peptide as independent predictors for cardiovascular disease in type 2 diabetes. Atherosclerosis 2018, 274:182–190. [DOI] [PubMed] [Google Scholar]
- 3.Jrøgensen AB, Frikke-Schmidt R, Nordestgaard BG, Tybjræg-Hansen A: Loss-of-function mutations in APOC3 and risk of ischemic vascular disease. N Engl J Med 2014, 371:32–41. [DOI] [PubMed] [Google Scholar]
- 4.Crosby J, Peloso GM, Auer PL, Crosslin DR, Stitziel NO, Lange LA, Lu Y, Tang ZZ, Zhang H, Hindy G, et al. : Loss-of-function mutations in APOC3, triglycerides, and coronary disease. N Engl J Med 2014, 371:22–31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Sundaram M, Curtis KR, Amir Alipour M, LeBlond ND, Margison KD, Yaworski RA, Parks RJ, McIntyre AD, Hegele RA, Fullerton MD, et al. : The apolipoprotein C-III (Gln38Lys) variant associated with human hypertriglyceridemia is a gain-of-function mutation. J Lipid Res 2017, 58:2188–2196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Zhang J, Rocha N de A, McCullough PA: Contribution of ApoCIII to Diabetic Dyslipidemia and Treatment With Volanesorsen. Rev Cardiovasc Med 2018, 19:13–19. [DOI] [PubMed] [Google Scholar]
- 7.Bremer AA, Stanhope KL, Graham JL, Cummings BP, Wang W, Saville BR, Havel PJ: Fructose-fed rhesus monkeys: a nonhuman primate model of insulin resistance, metabolic syndrome, and type 2 diabetes. Clin Transl Sci 2011, 4:243–252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- **8.Butler AA, Price CA, Graham JL, Stanhope KL, King S, Hung YH, Sethupathy P, Wong S, Hamilton J, Krauss RM, et al. : Fructose-induced hypertriglyceridemia in rhesus macaques is attenuated with fish oil or ApoC3 RNA interference. J Lipid Res 2019, 60:805–818. [DOI] [PMC free article] [PubMed] [Google Scholar]; This trial in non-human primates showed increased circulating apoCIII levels after fructose feeding and provided preliminary data on lipid lowering effects of RNA interference of apoCIII. They further showed that fish oil supplementation reduced apoCIII levels in the monkeys and attenuated the fructose-induced effects on circulating TG levels.
- 9.Stanhope KL, Medici V, Bremer AA, Lee V, Lam HD, Nunez MV, Chen GX, Keim NL, Havel PJ: A dose-response study of consuming high-fructose corn syrup-sweetened beverages on lipid/lipoprotein risk factors for cardiovascular disease in young adults. Am J Clin Nutr 2015, 101:1144–1154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- **10.Taskinen M-R, Söderlund S, Bogl LH, Hakkarainen A, Matikainen N, Pietiläinen KH, Räsänen S, Lundbom N, Björnson E, Eliasson B, et al. : Adverse effects of fructose on cardiometabolic risk factors and hepatic lipid metabolism in subjects with abdominal obesity. J Intern Med 2017, 140:874–888. [DOI] [PubMed] [Google Scholar]; This is one of the three currently-published reports that show fructose feeding increases circulating apoCIII levels. Fructose feeding over 12 weeks resulted in increased TG levels in the subject. 59% of the change in TG was explained by apoCIII in a 7-variable multivariate regression model. DNL only explained 16% of the change in circulating TG.
- **11.Hieronimus B, Griffen SC, Keim NL, Bremer AA, Berglund L, Nakajima K, Havel PJ, Stanhope KL, Hieronimus B, Griffen SC, et al. : Effects of Fructose or Glucose on Circulating ApoCIII and Triglyceride and Cholesterol Content of Lipoprotein Subfractions in Humans. J Clin Med 2019, 8:913. [DOI] [PMC free article] [PubMed] [Google Scholar]; Consumption of fructose over 10 weeks led to an increase in fasting and postprandial circulating apoCIII compared with consumption of glucose. The increase in postprandial apoCIII was associated with the increase in cholesterol in small LDL particles in fructose-consuming subjects. This suggests that apoCIII might be a causal link between fructose consumption and fructose-induced dyslipidemia.
- 12.Gugliucci A, Lustig RH, Caccavello R, Erkin-Cakmak A, Noworolski SM, Tai VW, Wen MJ, Mulligan K, Schwarz J-M: Short-term isocaloric fructose restriction lowers apoC-III levels and yields less atherogenic lipoprotein profiles in children with obesity and metabolic syndrome. Atherosclerosis 2016, 253:171–177. [DOI] [PubMed] [Google Scholar]
- 13.Lustig RH, Mulligan K, Noworolski SM, Tai VW, Wen MJ, Erkin-Cakmak A, Gugliucci A, Schwarz JM: Isocaloric fructose restriction and metabolic improvement in children with obesity and metabolic syndrome. Obes (Silver Spring) 2016, 24:453–460. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Caron S, Verrijken A, Mertens I, Samanez CH, Mautino G, Haas JT, Duran-Sandoval D, Prawitt J, Francque S, Vallez E, et al. : Transcriptional activation of apolipoprotein CIII expression by glucose may contribute to diabetic dyslipidemia. Arterioscler Thromb Vasc Biol 2011, 31:513–519. [DOI] [PubMed] [Google Scholar]
- 15.Motallebipour M, Enroth S, Punga T, Ameur A, Koch C, Dunham I, Komorowski J, Ericsson J, Wadelius C: Novel genes in cell cycle control and lipid metabolism with dynamically regulated binding sites for sterol regulatory element-binding protein 1 and RNA polymerase II in HepG2 cells detected by chromatin immunoprecipitation with microarray detection. FEBS J 2009, 276:1878–1890. [DOI] [PubMed] [Google Scholar]
- 16.Ginsberg HN, Brown WV: Apolipoprotein CIII: 42 years old and even more interesting. Arterioscler Thromb Vasc Biol 2011, 31:471–473. [DOI] [PubMed] [Google Scholar]
- 17.Altomonte J, Cong L, Harbaran S, Richter A, Xu J, Meseck M, Dong HH: Foxo1 mediates insulin action on apoC-III and triglyceride metabolism. J Clin Invest 2004, 114:1493–1503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Hiukka A, Fruchart-Najib J, Leinonen E, Hilden H, Fruchart JC, Taskinen MR: Alterations of lipids and apolipoprotein CIII in very low density lipoprotein subspecies in type 2 diabetes. Diabetologia 2005, 48:1207–1215. [DOI] [PubMed] [Google Scholar]
- 19.Bharara M et al. , Rajpathak SN, Gunter MJ, Wylie-rosett J, Ho GYF, Kaplan RC, Muzumdar R, Rohan TE, Strickler HD: The role of insulin-like growth factor-I and its binding proteins in glucose homeostasis and type 2 diabetes. Diabetes Metab Res Rev 2009, 28:3–12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Briones ER, Mao SJT, Palumbo PJ, O’Fallon WM, Chenoweth W, Kottke BA: Analysis of plasma lipids and apolipoproteins in insulin-dependent and noninsulin-dependent diabetics. Metabolism 1984, 33:42–49. [DOI] [PubMed] [Google Scholar]
- 21.Blackett P, Sarale DC, Fesmire J, Harmon J, Weech P, Alaupovic P: Plasma apolipoprotein C-III levels in children with type I diabetes. South Med J 1988, 81:469–473. [DOI] [PubMed] [Google Scholar]
- 22.Koo HY, Wallig MA, Chung BH, Nara TY, Cho BHS, Nakamura MT: Dietary fructose induces a wide range of genes with distinct shift in carbohydrate and lipid metabolism in fed and fasted rat liver. Biochim Biophys Acta - Mol Basis Dis 2008, 1782:341–348. [DOI] [PubMed] [Google Scholar]
- 23.Kim M, Lai M, Herman MA, Kim M, Krawczyk SA, Doridot L, Fowler AJ, Wang JX, Trauger SA, Noh H, et al. : ChREBP regulates fructose-induced glucose production independently of insulin signaling. J Clin Invest 2016, 126:4372–4386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Haas JT, Miao J, Chanda D, Wang Y, Zhao E, Haas ME, Hirschey M, Vaitheesvaran B, Farese RV., Kurland IJ, et al. : Hepatic insulin signaling is required for obesity-dependent expression of SREBP-1c mRNA but not for feeding-dependent expression. Cell Metab 2012, 15:873–884. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Koo H-Y, Miyashita M, Cho BHS, Nakamura MT: Replacing dietary glucose with fructose increases ChREBP activity and SREBP-1 protein in rat liver nucleus. Biochem Biophys Res Commun 2009, 390:285–289. [DOI] [PubMed] [Google Scholar]
- 26.Stanhope KL, Griffen SC, Bremer AA, Vink RG, Schaefer EJ, Nakajima K, Schwarz JM, Beysen C, Berglund L, Keim NL, et al. : Metabolic responses to prolonged consumption of glucose- and fructose-sweetened beverages are not associated with postprandial or 24-h glucose and insulin excursions. Am J Clin Nutr 2011, 94:112–119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Teff KL, Elliott SS, Tschöp M, Kieffer TJ, Rader D, Heiman M, Townsend RR, Keim NL, D’Alessio D, Havel PJ: Dietary fructose reduces circulating insulin and leptin, attenuates postprandial suppression of ghrelin, and increases triglycerides in women. J Clin Endocrinol Metab 2004, 89:2963–2972. [DOI] [PubMed] [Google Scholar]
- 28.Teff KL, Grudziak J, Townsend RR, Dunn TN, Grant RW, Adams SH, Keim NL, Cummings BP, Stanhope KL, Havel PJ: Endocrine and metabolic effects of consuming fructose- and glucose-sweetened beverages with meals in obese men and women: Influence of insulin resistance on plasma triglyceride responses. J Clin Endocrinol Metab 2009, 94:1562–1569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Stanhope K, Griffen SC, Bair BR, Swarbrick M, Keim N, Havel PJ: Twenty-four-hour endocrine and metabolic profiles following consumption of high-fructose corn syrup-, sucrose-, fructose-, and glucose-sweetened beverages with meals. Am J Clin Nutr 2008, 87:1194–1203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Softic S, Cohen DE, Kahn CR: Role of Dietary Fructose and Hepatic De Novo Lipogenesis in Fatty Liver Disease. Dig Dis Sci 2016, 61:1282–1293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Yao Z, Wang Y: Apolipoprotein C-III and hepatic triglyceride-rich lipoprotein production. Curr Opin Lipidol 2012, 23:206–212. [DOI] [PubMed] [Google Scholar]
- 32.Erkin-Cakmak A, Bains Y, Caccavello R, Noworolski SM, Schwarz JM, Mulligan K, Lustig RH, Gugliucci A: Isocaloric Fructose Restriction Reduces Serum d-Lactate Concentration in Children with Obesity and Metabolic Syndrome. J Clin Endocrinol Metab 2019, 104:3003–3011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- *33.Taskinen MR, Packard CJ, Borén J: Emerging Evidence that ApoC-III Inhibitors Provide Novel Options to Reduce the Residual CVD. Curr Atheroscler Rep 2019, 21. [DOI] [PMC free article] [PubMed] [Google Scholar]; This review summarizes the mechanisms by which apoCIII contributes to undesirable cardiometabolic health outcomes. The authors further review clinical trials using apoCIII-lowering drugs.
- **34.Ramms B, Patel S, Nora C, Pessentheiner AR, Chang MW, Green CR, Golden GJ, Secrest P, Krauss RM, Metallo CM, et al. : ApoC-III ASO promotes tissue LPL activity in the absence of apoE-mediated TRL clearance. J Lipid Res 2019, 60:1379–1395. [DOI] [PMC free article] [PubMed] [Google Scholar]; This study indicates that apoE determines the function of apoCIII. In the absence of apoE, apoCIII inhibits lipoprotein lipase in the adipose tissue, while in the presence of apoE it blocks hepatic clearance of triglyceride rich lipoproteins. This study emphasizes the importance of assessing multiple apolipoproteins and the complexity of their interactions.
- 35.Zheng C, Khoo C, Furtado J, Ikewaki K, Sacks FM: Dietary monounsaturated fat activates metabolic pathways for triglyceride-rich lipoproteins that involve apolipoproteins E and C-III. Am J Clin Nutr 2008, 88:272–281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Stanhope KL, Bremer AA, Medici V, Nakajima K, Ito Y, Nakano T, Chen G, Fong TH, Lee V, Menorca RI, et al. : Consumption of fructose and high fructose corn syrup increase postprandial triglycerides, LDL-cholesterol, and apolipoprotein-B in young men and women. J Clin Endocrinol Metab 2011, 96:E1596–605. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Mardinoglu A, Wu H, Bjornson E, Zhang C, Hakkarainen A, Räsänen SM, Lee S, Mancina RM, Bergentall M, Pietiläinen KH, et al. : An Integrated Understanding of the Rapid Metabolic Benefits of a Carbohydrate-Restricted Diet on Hepatic Steatosis in Humans. Cell Metab 2018, 27:559–571. e5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Adiels M, Olofsson SO, Taskinen MR, Borén J: Overproduction of very low-density lipoproteins is the hallmark of the dyslipidemia in the metabolic syndrome. Arterioscler Thromb Vasc Biol 2008, 28:1225–1236. [DOI] [PubMed] [Google Scholar]
- 39.Te Morenga LA, Howatson AJ, Jones RM, Mann J: Dietary sugars and cardiometabolic risk: Systematic review and meta-analyses of randomized controlled trials of the effects on blood pressure and lipids. Am J Clin Nutr 2014, 100:65–79. [DOI] [PubMed] [Google Scholar]
- 40.März W, Kleber ME, Scharnagl H, Speer T, Zewinger S, Ritsch A, Parhofer KG, von Eckardstein A, Landmesser U, Laufs U: HDL cholesterol: reappraisal of its clinical relevance. Clin Res Cardiol 2017, 106:663–675. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Riwanto M, Rohrer L, Roschitzki B, Besler C, Mocharla P, Mueller M, Perisa D, Heinrich K, Altwegg L, Von Eckardstein A, et al. : Altered activation of endothelial anti-and proapoptotic pathways by high-density lipoprotein from patients with coronary artery disease: Role of high-density lipoprotein-proteome remodeling. Circulation 2013, 127:891–904. [DOI] [PubMed] [Google Scholar]
- 42.Farbstein D, Levy AP: HDL dysfunction in diabetes: Causes and possible treatments. Expert Rev Cardiovasc Ther 2012, 10:353–361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- *43.Jensen MK, Aroner SA, Mukamal KJ, Furtado JD, Post WS, Tsai MY, Tjønneland A, Polak JF, Rimm EB, Overvad K, et al. : High-Density Lipoprotein Subspecies Defined by Presence of Apolipoprotein C-III and Incident Coronary Heart Disease in Four Cohorts. Circulation 2018, 137:1364–1373. [DOI] [PMC free article] [PubMed] [Google Scholar]; Samples from four prospective cohort studies were analyzed for their apoCIII content of HDL particles. The subgroup of HDL particles containing apoCIII was associated with a higher risk of CVD, while the absence of apoCIII on HDL particles was associated with a lower risk.
- 44.Luo M, Liu A, Wang S, Wang T, Hu D, Wu S, Peng D: ApoCIII enrichment in HDL impairs HDL-mediated cholesterol efflux capacity. Sci Rep 2017, 7:1–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Morton AM, Koch M, Mendivil CO, Furtado JD, Tjønneland A, Overvad K, Wang L, Jensen MK, Sacks FM: Apolipoproteins E and CIII interact to regulate HDL metabolism and coronary heart disease risk. JCI insight 2018, 3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- *46.Christopoulou E, Tsimihodimos V, Filippatos T, Elisaf M: Apolipoprotein CIII and diabetes. Is there a link? Diabetes Metab Res Rev 2019, 35:1–8. [DOI] [PubMed] [Google Scholar]; This review outlined possible mechanisms by which apoCIII affects insulin sensitivity. As reviewed, apoCIII affects glucose homeostasis likely by enhancing insulin resistance and impairment of the anti- diabetogenic properties of HDL. In addition, they summarized the evidence of apoCIII induced apoptosis in pancreatic ß-cells.
- 47.Stanhope KL, Schwarz JM, Keim NL, Griffen SC, Bremer AA, Graham JL, Hatcher B, Cox CL, Dyachenko A, Zhang W, et al. : Consuming fructose-sweetened, not glucose-sweetened, beverages increases visceral adiposity and lipids and decreases insulin sensitivity in overweight/obese humans. J Clin Invest 2009, 119:1322–1334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Girona J, Amigó N, Ibarretxe D, Plana N, Rodríguez-Borjabad C, Heras M, Ferré R, Gil M, Correig X, Masana L: HDL triglycerides: A new marker of metabolic and cardiovascular risk. Int J Mol Sci 2019, 20:1–10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Chapman MJ, Le Goff W, Guerin M, Kontush A: Cholesteryl ester transfer protein: At the heart of the action of lipid-modulating therapy with statins, fibrates, niacin, and cholesteryl ester transfer protein inhibitors. Eur Heart J 2010, 31:149–164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Kontush A, John Chapman M: Why is HDL functionally deficient in type 2 diabetes? Curr Diab Rep 2008, 8:51–59. [DOI] [PubMed] [Google Scholar]
- 51.Taskinen MR, Borén J: New insights into the pathophysiology of dyslipidemia in type 2 diabetes. Atherosclerosis 2015, 239:483–495. [DOI] [PubMed] [Google Scholar]