
Figure 4. Chromatin dynamics in regeneration.
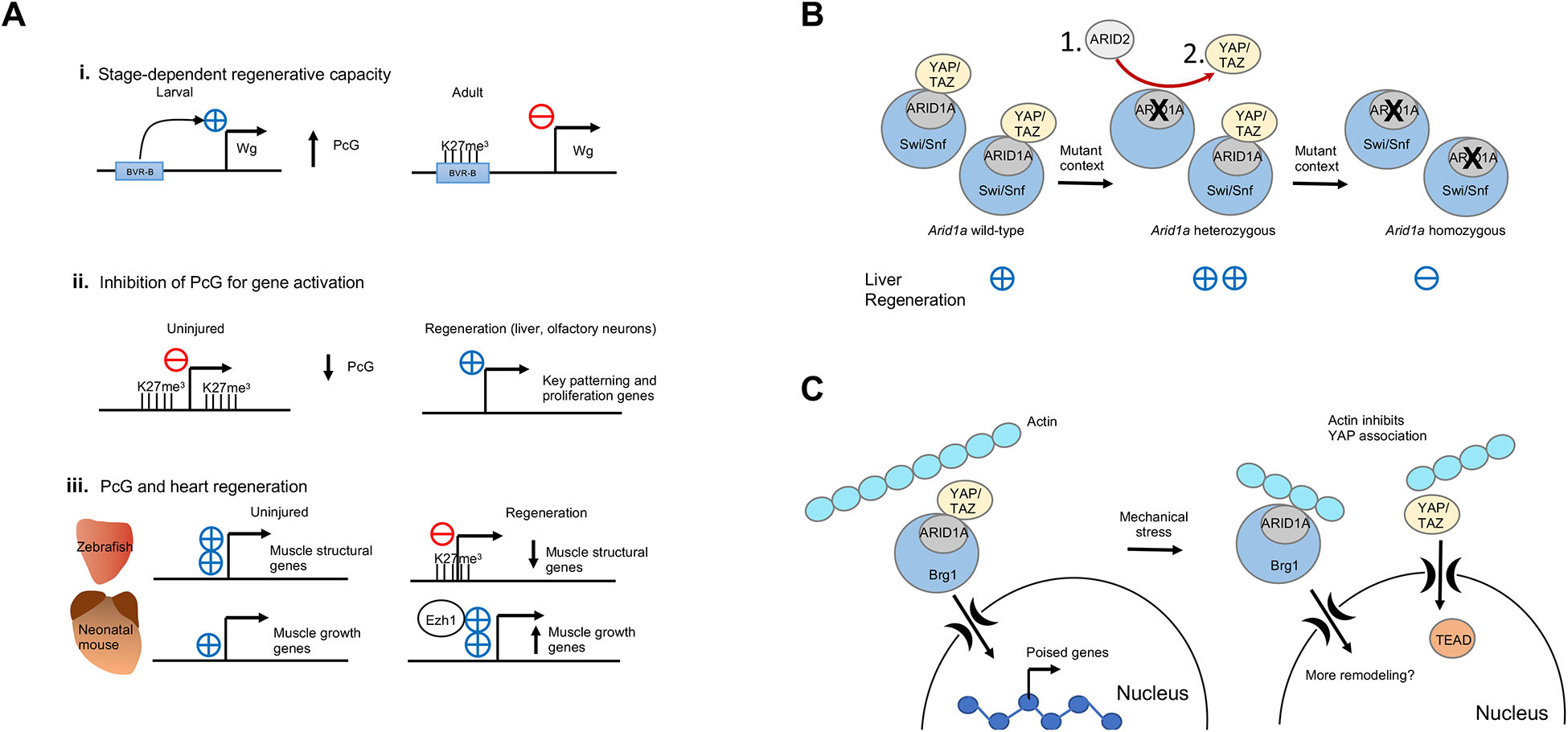
A. The role of Polycomb regulation in regeneration. Polycomb signaling can silence pro-regenerative enhancers, restricting regeneration, as shown here for the BRV-B enhancer of the wingless gene during imaginal disc regeneration in Drosophila. Polycomb silencing is sometimes inhibited in order to activate pro-regeneration genes, such as cell-cycle genes, in the case of regeneration of liver or olfactory neurons. Polycomb has different functions during heart regeneration depending on the model organism; in adult zebrafish, Polycomb silences highly expressed muscle structural genes that encode central components of the sarcomere such as titin and the myosin heavy and light chains (cmlc1, ttna, ttnb, vmhcl). Alternatively, in neonatal mice, Polycomb subunit Ezh1 acts as a transcriptional activator for genes involved in tissue morphogenesis, muscle structure and metabolism such as Bmp7, Irx2 and Tnni1.
B. Heterozygosity of Arid1a promotes liver regeneration. Some possible mechanisms include exchange of Arid1a subunits with Arid1b or Arid107 (1), a decrease in Hippo signaling via YAP/TAZ (2), or the fine tuning of gene expression to restrict antagonistic pathways (not pictured).
C. ATP-dependent chromatin remodeling complex Swi/Snf promotes regeneration after mechanical stress. In the uninjured setting, the ARID1A subunit of Swi/Snf sequesters YAP/TAZ and prevents it from binding its co-factor, TEAD-1. Swi/Snf remodels chromatin to poise pro-regeneration genes for activation in the liver. Upon mechanical stress, actin binds to ARID1A and releases YAP/TAZ, which can then associate with its active cofactor TEAD-1 and activate regeneration. Swi/Snf itself may then also further remodel chromatin102.