
Figure 3.
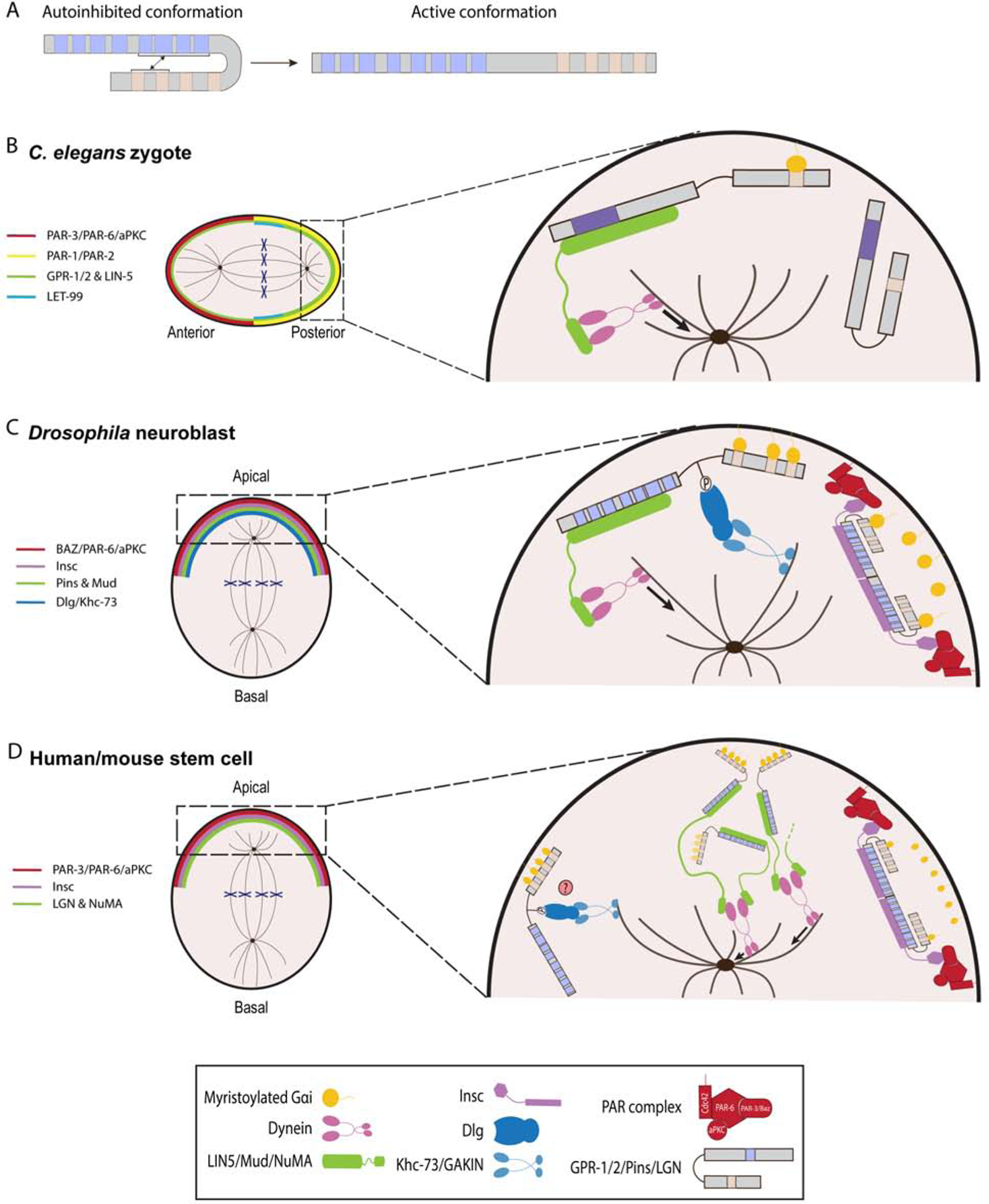
AGS-family proteins regulate spindle orientation and ACD. A, Schema depicting a transition of human LGN from a closed to an open conformation. In autoinhibited form, the GL motifs bind to the TPR motifs. B-D, Working models of AGS-family protein function in C. elegans zygote (A), Drosophila neuroblasts (B), and human/mouse stem cells (C). For each AGS-family protein, blue squares indicate TPR motifs, beige squares indicate GL motifs, and purple squares (C. elegans) indicate debated TPR motifs. A, C. elegans zygote is polarized along an anterior-posterior axis by the activity of PAR proteins complex. GPR-1/2 protein is localized to the posterior complex as LET-99 restricts GPR-1/2 to posterior-most cortex. GPR-12 is anchored at the cortex through the GL motifs that are bound to myristoylated Gαi. Pins then interacts with LIN-5 to recruit dynein and generate microtubule pulling forces on the astral microtubules. Of note, GPR-1/2 and LIN-5 are more concentrated towards the posterior cortex. B, Drosophila neuroblast is polarized along an apical-basal axis by PAR protein complex. Pins forms heterotetramers with Insc to localize to the apical cortex. Pins then recruits Mud as well as Dlg and Khc-73 proteins to orient mitotic spindle. C, Human and mouse stem cells divide asymmetrically along an apical-basal polarity axis via LGN. LGN is able to form heterotetramers with Insc for its localization and heterohexamers with NuMA, which creates a network that recruits dynein to the cortex. However, the functional interactions between such oligomers remain to be determined. The LGN/Dlg/GAKIN is predicted to be involved in spindle orientation but has yet to be shown.