
Figure 1.
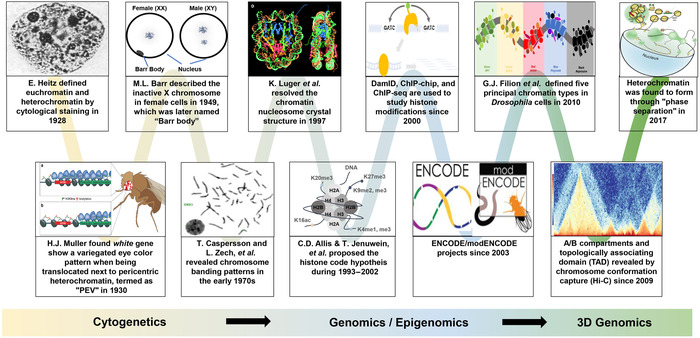
The concept of heterochromatin has been evolving since its first description in 1928 by Emil Heitz. Shown is an incomplete list of events that led to changes in the concept of heterochromatin during research developments from the cytogenetic era until very recently in the 3D genomics era. The characterization of proteins (e.g., HP1) regulating constitutive heterochromatin was largely attributed to the genetic screens in Drosophila for mutants affecting the position‐effect variegation (PEV) phenotype. The studies into facultative heterochromatin were first marked by the discovery of the Barr body, that is, the female‐specific inactivated X chromosome. After the discovery of nucleosome structure as the chromatin unit for gene regulation, many PEV‐related genes were found to be the reader or writer of histone modifications within the chromatin unit. With the development of chromatin immunoprecipitation (ChIP) technology targeting these histone modifications, combined with either microarray (ChIP‐chip) or sequencing analyses (ChIP‐seq), it became clear that heterochromatin is usually associated with highly repetitive regions and the histone methylation marks H3K9me2/3 and H3K27me3. This facilitated the definition of the heterochromatin region at the base pair resolution in the genomic and epigenomic era. Recently, it has been shown in Drosophila and humans that heterochromatin forms within phase‐separated droplets of HP1 protein and forms distinct topologically associated domains from those of euchromatin, as detected by Hi‐C technology.