
Figure 3.
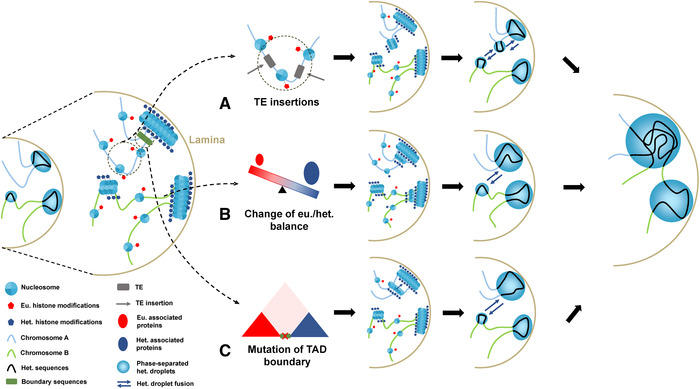
Transitions from euchromatin to heterochromatin. Three proposed models of euchromatin to heterochromatin transition. Heterochromatin is usually distributed close to the nuclear periphery and tethered to the lamina or around the nucleoli, while euchromatin is located in the nuclear interior. (A) The de novo formation of heterochromatin domains induced by TE insertions. A TE insertion into the euchromatic region may trigger heterochromatin formation mediated by small RNA pathways. This may further impact the expression of nearby genes by the spreading effect of newly formed heterochromatin domains. (B) Change in euchromatin/heterochromatin balance. The expansion of heterochromatin or ectopic formation of heterochromatin can be caused by upregulation of heterochromatin‐associated proteins (e.g., histone methyltransferase SU(VAR)3‐9, shown as dark blue circles) or downregulation of euchromatin‐associated proteins (red circles). This has been demonstrated in Drosophila and yeast. Such chromatin boundaries form without the participation of boundary elements, such as CTCF proteins, and are thus called negotiable borders. (C) Mutations of TAD boundary sequences (green bars between the two TADs) between euchromatin and heterochromatin domains. The TAD boundary sequences are usually CTCF binding sites or transcriptionally active genes or TEs. Removal or inversion of such boundary sequences may lead to the expansion of heterochromatin domains into euchromatin. The newly formed heterochromatin domains through the TE insertions, A, or expanded heterochromatin domains through B or C will convergently interact with other preexisting heterochromatin domains at the lamina through fusions of phase‐separated droplets. Such interactions may impact the nearby genes or genes on a different chromosome by reshaping the genome‐wide folding. Het, heterochromatin; Eu, euchromatin; TE, transposable element.