
Fig. 6.
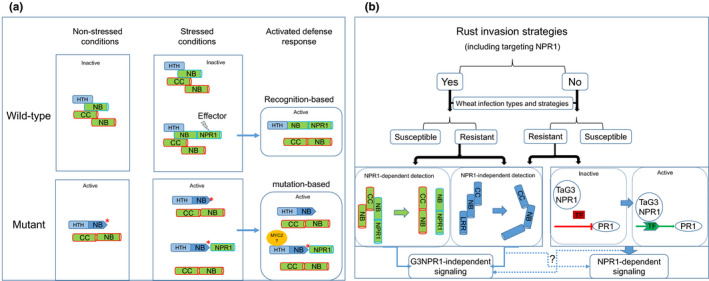
Models of TaNPR1‐induced resistance and the potential role of TaNPR1 proteins during wheat–rust interactions. (a) Model of resistance conferred by NPR1‐dependent detection in an R protein complex by a pair of head‐to‐head NB‐ARC genes. In wild‐type during the absence of rust, the two NB‐ARC genes fold together in an inactive formation. In the presence of a rust pathogen, one of the NB‐ARC loci produces an isoform protein containing NPR1 as a decoy. Two types of R protein complexes will be formed. If rust secretes an effector to attack TaNPR1, the decoy NPR1 could be mistakenly attacked, and then the NPR1‐decoy R protein complex will change to an active formation to induce a defence response. More NB‐NPR1 isoform will be produced to distract the NPR1‐attacking effector. In a mutant in which a mutation results in a change in the R protein complex to an active formation, the defence response is activated by the mutation. The mutant will be resistant to all rusts, except ones that have effectors to suppress other components of the defence signalling pathway. (b) During wheat–rust interactions, rust pathogens may apply invasion strategies that target or not target TaG3NPR1 (shown as Yes or No, respectively, on the figure). Without an ineffective detection system or compromised components for inducing defence signals, wheat will be susceptible to the pathogens. Resistant wheat in the ‘Yes’ group of rusts may use TaNPR1 as a decoy (e.g. Ta7ANPR1) in detection and have an alternative pathway to bypass TaNPR1 once the integrity of the NPR1 is compromised by the pathogens. Resistant wheat in the ‘No’ group of rusts may use either NPR1‐independent (e.g. Sr33) or NPR1‐dependent signalling through TaG3NPR1. Whether defence signalling could be transduced by both NPR1‐dependent or NPRI‐independent pathways is still a question to be resolved.