
Figure 1.
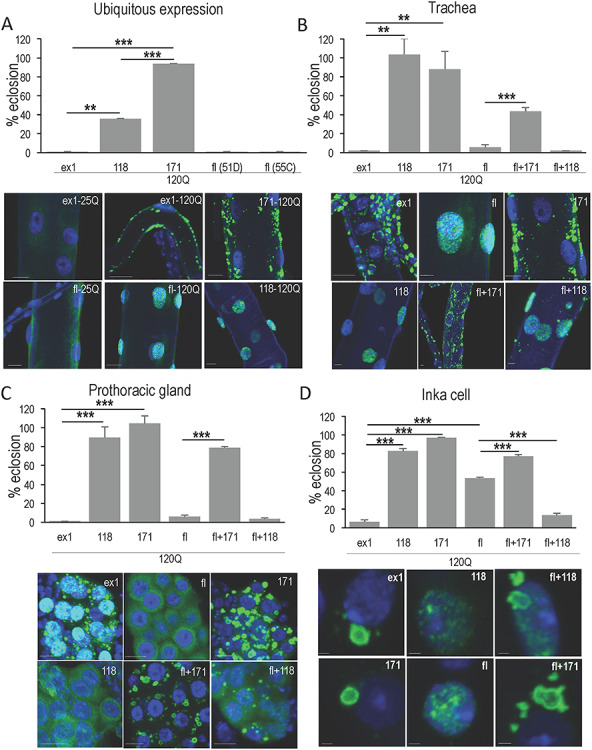
Cellular context influences subcellular behavior and pathology. (A) Ubiquitous expression with da > Gal4 reveals differences in pathogenic potential and subcellular behavior of HTT peptides. Various UAS > HTT/act > GFP males were crossed to females homozygous for the ubiquitously expressed da > Gal-4 driver, and survival to adulthood (eclosion) was monitored. Transgenes included HTT exon1 (ex1-120Q), full-length HTT (fl-120Q), HTT 118, shortstop (118-120Q) and HTT 171 (171-120Q). The percent flies eclosing as adults is calculated in reference to the internal control of non-expressing siblings (i.e. the number of UAS > HTT/+; +/da > Gal4 flies divided by UAS > HTT/+; +/act > GFP flies). Flies expressing unexpanded 25Q versions of these transgenes eclosed at a rate indistinguishable from the non-expressing controls. Peptide composition also influences the subcellular location of HTT transgenes here shown in trachea. HTTex1 with unexpanded polyQ (25 Qs) presents as diffuse cytoplasmic (ex1-25Q) while HTTex1 with 120Qs forms large cytoplasmic aggregates in tracheal cells (ex1-120Q). HTT118 with 120Qs is diffuse nuclear (118-120Q) while HTT171 forms cytoplasmic aggregates (117-120Q). Full-length HTT with 25Q is also diffuse cytoplasmic (fl-25Q) while expansion of the polyQ in full-length HTT converts it to diffuse nuclear in tracheal cells (fl-120Q). Scale bars are 10 μm. Since ubiquitous expression revealed a number of phenotypes that resembled those seen in different tissues, we tested whether multiple tissues exhibit pathology. (B) The toxicity of ex1, fl, 171 and 118 when driven exclusively in the trachea with btl > Gal4 at 25° was evaluated and the subcellular behavior determined by immunofluorescence. Expression of HTTex1-120Q and flHTT-120Q exclusively in trachea produced virtually complete toxicity similar to that seen with ubiquitous expression in (A). Many trachea in both HTTex1 and fl-mHTT expressing animals were visibly defective with gaps and constrictions evident (not shown) even though HTTex1 is clearly cytoplasmic and aggregated while fl-mHTT is diffuse nuclear implying multiple targets for pathology. In contrast, HTT118-120Q is diffuse nuclear and HTT171-120Q is cytoplasmic and aggregated and yet neither exhibits toxicity in trachea. The effect of the HTT171 fragment on fl-mHTT behavior and toxicity was examined by co-expressing both (fl + 171). Notably, the lethality of fl-mHTT is reduced when HTT171-120Q is expressed concurrently and the previously diffuse nuclear flHTT-120Q is now in cytoplasmic aggregates together with HTT171-120Q (fl + 171). (C) To further test the influence of tissue type, the toxicity of ex1, fl, 171 and 118 when driven in the prothoracic gland (PG) with spok > Gal4 at 29° was evaluated, as was the effect of the 171 fragment on fl toxicity in this tissue. In the PG flHTT-120Q is toxic as well and yet it is cytoplasmic and diffuse in these cells (fl). HTTex1-120Q is also toxic and continues to form cytoplasmic aggregates (ex1). HTT118 is relatively benign as it is in trachea although in the PG cells it exhibits a diffuse cytoplasmic distribution (118) and HTT171 is also non-toxic in the PG and occurs in cytoplasmic aggregates (171). When HTT171-120Q is co-expressed with flHTT-120Q, toxicity is significantly suppressed and the two proteins appear together in cytoplasmic aggregates (fl + 171). (D) Since ubiquitous expression produces crytptocephal phenotypes reminiscent of failure to produce edysis-triggering hormone, the toxicity of peptides driven in the 14 ETH producing INKA cells with ETH > Gal4 at 25° was evaluated. As in other tissues, HTTex1 is highly toxic in INKA cells and tends to form large cytoplasmic aggregates, while fl-mHTT exhibits only ~ 47% toxicity and is distributed as diffuse nuclear material. HTT171 exhibits no toxicity and forms large cytoplasmic aggregates similar to HTTex1, while HTT118 produces ~ 20% lethality and is distributed as diffuse nuclear material typically with a single perinucleolar punctum (118). Coexpression of fl-mHTT and HTT171 improves the survival of fl-mHTT expressing animals and recruits fl-mHTT into large cytoplasmic co-aggregates. Thus, both aggregated and diffuse HTT fragments can be toxic, and expanded polyQ (120Q) is not sufficient for either toxicity or aggregation.