
Figure 9.
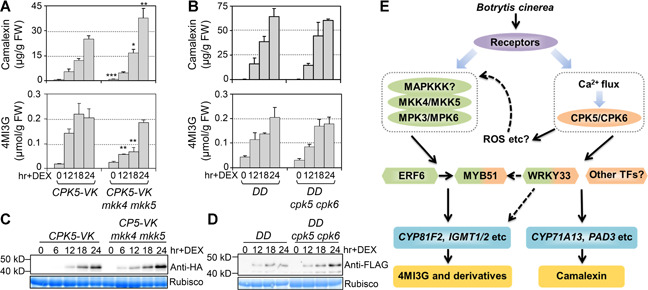
Independent functions of CPK5 and MPK3/MPK6 signaling in 4MI3G and camalexin induction
(A) Twelve‐d‐old CPK5‐VK and CPK5‐VK mkk4 mkk5 plants grown in liquid medium were treated with 5 μM DEX. Student t‐test was performed to compare the levels of camalexin or 4MI3G between CPK5‐VK mkk4 mkk5 and CPK5‐VK at each time point, *P < 0.05, **P < 0.01, ***P < 0.001. (B) DD and DD cpk5 cpk6 plants grown in liquid medium were treated with 5 μM DEX. Camalexin and 4MI3G were measured at indicated time points. Two‐way ANOVA analysis revealed that no significant group difference exists in camalexin and 4MI3G level change between different genotypes. Values are means ± SD, n = 3. FW: Fresh weight. (C) Protein levels of CPK5‐VK in CPK5‐VK and CPK5‐VK mkk4 mkk5 plants after DEX treatment were determined by immunoblot analysis using an anti‐HA antibody. (D) Protein levels of NtMEK2DD in DD and DD cpk5 cpk6 plants after DEX treatment were determined by immunoblot analysis using an anti‐FLAG antibody. Equal loading of proteins was confirmed by Coomassie brilliant blue staining. (E) A model depicts the independent and cooperative interaction of MPK3/MPK6 cascade and CPK5/CPK6 signaling in regulating 4MI3G and camalexin biosynthesis in response to B. cinerea infection. In response to B. cinerea infection, both CPK5/CPK6 and MPK3/MPK6 signaling pathways can be rapidly activated. The secondary responses/signals, such as ROS, after activation of CPK5/CPK6 could result in the change of the activity of MPK3/MPK6. The interplay between CPK5/CPK6 and MPK3/MPK6 also could converge on their common downstream targets, including the transcription factors WRKY33 and MYB51, to fine‐tune the downstream defense response.