

Abstract

Human milk is considered the golden standard in infant nutrition. Free oligosaccharides in human milk provide important health benefits. These oligosaccharides function as prebiotics, immune modulators, and pathogen inhibitors and were found to improve barrier function in the gut. Infant formulas nowadays often contain prebiotics but lack the specific functions of human milk oligosaccharides (hMOS). Milk from domesticated animals also contains milk oligosaccharides but at much lower levels and with less diversity. Goat milk contains significantly more oligosaccharides (gMOS) than bovine (bMOS) or sheep (sMOS) milk and also has a larger diversity of structures. This review summarizes structural studies, revealing a diversity of up to 77 annotated gMOS structures with almost 40 structures fully characterized. Quantitative studies of goat milk oligosaccharides range from 60 to 350 mg/L in mature milk and from 200 to 650 mg/L in colostrum. These levels are clearly lower than in human milk (5–20 g/L) but higher than in other domesticated dairy animals, e.g., bovine (30–60 mg/L) and sheep (20–40 mg/L). Finally, the review focuses on demonstrated and potential functionalities of gMOS. Some studies have shown anti-inflammatory effects of mixtures enriched in gMOS. Goat MOS also display prebiotic potential, particularly in stimulating growth of bifidobacteria preferentially. Although functional studies of gMOS are still limited, several structures are also found in human milk and have known functions as immune modulators and pathogen inhibitors. In conclusion, goat milk constitutes a promising alternative source for milk oligosaccharides, which can be used in infant formula.
Keywords: goat milk oligosaccharides, prebiotics, health benefits, structures, quantitative analysis
Introduction
The composition of human milk is very different from that of domesticated dairy animals (Figure 1).1−6 One of the major components, human milk oligosaccharides (hMOS), constitutes a relatively large part of the carbohydrates present in mother’s milk (5–20 g/L),3,7 representing up to 20% of the total carbohydrate content. The diversity of hMOS is large, with 247 varieties observed and 162 hMOS structurally characterized.8 The composition and abundance of hMOS vary between women with the genetics of the mothers and during lactation,9−12 but also evidence emerges that gestational age, maternal health status, infant sex, and dietary habits influence hMOS composition.11,13,14 These hMOS play a role in steering a healthy development of the infant gut microbiome, acting as prebiotics, stimulating the growth of beneficial bacteria. Beneficial bacteria growing on prebiotics secrete short-chain fatty acid (SCFA) and other metabolites (postbiotics), having a beneficial effect on host health.15 Although other prebiotics are added to infant formula, such as galactooligosaccharides (GOS) and fructooligosaccharides (FOS), hMOS have well-established and specific functionalities. Oligosaccharides in human milk have been found to inhibit pathogen adhesion, stimulate the immune system of infants, modulate host receptors, and modify epithelial glycosylation as well as stimulating brain development.16,17 Some of these functions have also been observed for GOS in a few studies, particularly stimulating gut barrier function, but have been less well-established.18−21
Figure 1.

Major milk components (g/L), fat, lactose, casein, and whey, in human (blue), goat (green), cow (purple) and sheep (red) milk, compiled from available literature sources.1,2,4−6
Bovine milk is traditionally the base for infant formula, as a substitute for mother’s milk when breastfeeding is not possible or mothers choose not to breastfeed. There are fewer studies of bovine milk oligosaccharides (bMOS) than of hMOS, but the overall view is that bMOS concentrations are much lower (30–60 mg/L) compared to hMOS (5–20 g/L).3,22 Moreover, the bMOS composition with 40–50 varieties observed and 37 structures identified is much less varied23,24 than the hMOS composition (∼247 varieties with 162 structures identified).8 Goat milk is an approved protein source for infant formula and has attracted attention as a result of higher levels and diversity in milk oligosaccharides, with average concentrations reported of 60–350 mg/L in mature milk and up to 2.4 g/L in colostrum.25−30 The oligosaccharide composition of goat milk (gMOS) has been studied24,27−29,31−35 and compared to other domesticated animals.24,29 A few studies have reported variations of oligosaccharides at the goat breed level,25,26 over time during lactation,25,28,36 in parity,26 or at the individual goat level.37 Such information is considered interesting, because gMOS are more abundant than bMOS and contain certain structures that are in common with hMOS (Schemes 1 and 2). Therefore, goat milk has potential to display special functional properties that may find application in a range of commercial products in various niches.
Scheme 1. Overview of All Neutral gMOS Structures Reported in Goat Colostrum and Milk, with Relative Abundances Reported in Neutral and Acidic Pools, Respectively.

The hMOS column (Y/N) indicates whether the structure has been observed in human milk. Hypothetical interactions with galectins are indicated, according to Urashima et al.8 Possible interaction epitopes are marked green; blocked epitopes are marked red; and gray boxes indicate uncertain interactions where exact structures are not known, with relative peak intensities in the neutral and acidic pool, respectively (on the basis of the study by Albrecht et al.24).
Scheme 2. Overview of All Acidic gMOS Structures Reported in Goat Colostrum and Milk, with Relative Abundances Reported in Neutral and Acidic Pools, Respectively.
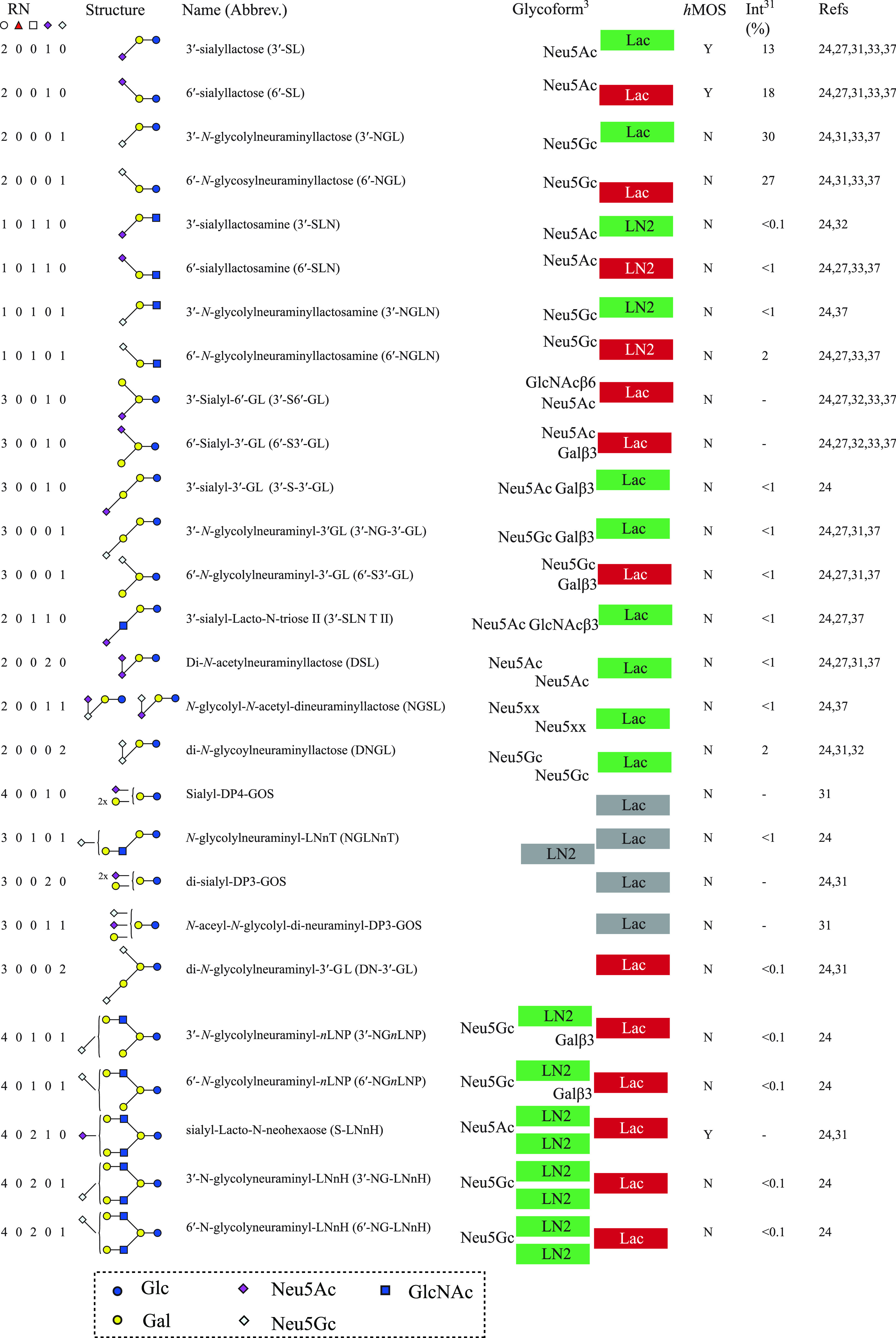
The hMOS column (Y/N) indicates whether the structure has been observed in human milk. Hypothetical interactions with galectins are indicated, according to Urashima et al.8 Possible interaction epitopes are marked green; blocked epitopes are marked red; and gray boxes indicate uncertain interactions where exact structures are not known, with relative peak intensities in the neutral and acidic pool, respectively (on the basis of the study by Albrecht et al.24).
In this literature overview, we will evaluate the current knowledge of gMOS structures and quantities, also in relation to what is known about hMOS and bMOS. An overview will be presented on functional properties of MOS in general as well as specific structure–function relationships that have been discovered thus far for hMOS, bMOS, and gMOS.
Goat Milk Oligosaccharide (gMOS) Structures
The first gMOS were separated and purified from Japanese Saanen goat colostrum.31 Detailed structural characterization identified three novel compounds, Gal(α1–3)Gal(β1–4)Glc (α3′-GL), Gal(β1–3)Gal(β1–4)Glc (3′-GL), Gal(β1–6)Gal(β1–4)Glc (6′-GL), and Fuc(α1–2)Gal(β1–4)Glc (2′-FL).31 In 1997, the same research group reported the structures of the first four acidic gMOS from Japanese Saanen, namely, Neu5Ac(α2–6)Gal(β1–4)Glc (6′-SL), Neu5Ac(α2–3)Gal(β1–4)Glc (3′-SL), Neu5Gc(α2–6)Gal(β1–4)Glc (6′-NGL), and Neu5Ac(α2–6)Gal(β1–4)GlcNAc (6′-SLN).33 This study showed non-human sialic acid N-glycolylneuraminic acid (Neu5Gc) occurring in gMOS.33 Another group published the structures of two branched acidic gMOS from mature goat milk, i.e., Gal(β1–3)[Neu5Ac(α2–6)]Gal(β1–4)Glc (6′-SHL) and Gal(β1–6)[Neu5Ac(α2–3)]Gal(β1–4)Glc (3′-SHL).32
After these initial efforts to isolate specific oligosaccharides from goat milk and to identify their structures with detailed analytical methods, more comprehensive studies were performed, identifying structural compositions, i.e., the monosaccharide makeup of the structures, based on mass spectrometry (MS) data. When these data were combined with reference structures and previous literature, comparative studies could be made, following known structures and annotating newly found oligosaccharide structures. One of the first comparative studies focused on the gMOS composition of fresh mature milk from Spanish goats from the Murciano-Granadina breed, from cows and sheep, compared to hMOS.29
A detailed and comprehensive study was conducted by Albrecht et al., in which milk samples of different domesticated animals, including goats, were analyzed for their milk oligosaccharide composition.24 Structural oligosaccharide libraries were created using a combination of analytical techniques. Purified MOS were fluorescently derivatized via reductive amination with 2-aminobenzamide (2-AB). This stoichiometric addition of one 2-AB label per oligosaccharide allowed for their relative quantification with fluorescence detection.38 Some oligosaccharides were identified on the basis of their elution compared to standards. All other structures were identified by exoglycosidase assays in combination with tandem mass spectrometry (MS/MS) data. Sialic acid determination was performed to distinguish between Neu5Gc and Neu5Ac epitopes.31
Albrecht et al. found that milk from domesticated animals contained a much larger variety in complex MOS than previously known. More than 80% of the oligosaccharides found in pools of all domesticated animals were acidic oligosaccharides. In total, 77 structures were identified in all of the milk samples, of which 29 were neutral, 45 were sialylated, and 3 were phosphorylated.24 A total of 40 structures were characterized in mature goat milk, of which 16 were neutral, 23 were acidic, and 1 was phosphorylated lactose (Schemes 1 and 2). Numerically, sheep and bovine milk are only slightly less diverse, with 37 and 32 distinct structures, respectively. However, in comparison of the total milk profile, bovine milk is dominated by one structure, i.e., 3′-SL, while sheep milk is dominated by 3′-N-glycolylneuraminyllactose (3′-NGL) and 6′-N-glycolylneuraminyllactose (6′-NGL) and goat milk shows a much more varied peak pattern in this comparative study.24 From the sialylated pool, the proportion of Neu5Gc-containing oligosaccharides was highest in sheep colostrum (94%) and mature goat milk (64%). The acidic pool of cow colostrum contained 3% Neu5Gc oligosaccharides, whereas only trace amounts were found in the milk of other domesticated animals (≤1%).24 Humans are unique in their inability to produce Neu5Gc but are able to incorporate dietary Neu5Gc in their glycosylation.39,40 Incorporation of Neu5Gc has also been observed in hMOS.41 Studies have associated Neu5Gc incorporation with several diseases, including cancer, type 2 diabetes, atherosclerosis, and autoimmune diseases.40,42 Although there is no causality proven for Neu5Gc, potential health effects of the relatively high levels of Neu5Gc in goat milk should be taken into account. In the study by Albrecht et al., 19 structures were identified for the first time in goat milk along with 3-FL, one of the more abundant hMOS constituents.24 A later study annotated 78 variations in goat colostrum but did not identify specific structures; only monosaccharide compositional data were obtained.37
Using more sensitive techniques, a comparative study between αs1-casein-producing and non-producing goats was performed.27 Previously reported structure compositions were observed and verified by MS/MS. This study annotated in total 37 oligosaccharide structures. A fucosyllactosamine structure with monosaccharide composition 1Hex, 1Fuc, 1HexNAc, 0Neu5Ac, and 0Neu5Gc was exclusively found in milk of goats unable to synthesize αs1-casein. Another fucosylated gMOS composed of 3Hex, 1Fuc, 3HexNAc, 0Neu5Ac, and 0Neu5Gc was present in both types of milk but only in trace amounts in milk of αs1-casein-producing goats. For all other structures, no significant differences in abundance were observed.
Taken together, the structural diversity of human milk is much higher than that of any domesticated dairy animal, i.e., 247 compositional variations and 162 identified structures in human milk, as compared to 78 compositions and 40 structures in goat milk, 50 compositions and 37 structures in bovine milk, and 32 structures in sheep milk; as a result of fewer studies, there is no extra data on composition variety in sheep milk at this time.8,23,24,37 With focus on the major structures that are responsible for >95% of the quantitative composition on the basis of relative abundance studies, human milk contains 22 major structures,3,43 while goat milk is defined by 9 major structures, bovine milk is defined by 6 structures, and sheep milk is defined by only 4.24 It is clear that goat milk presents a more diverse pallet of MOS structures than bovine or sheep milk.
There are some clear differences in the structural composition and number of oligosaccharides between human milk and milk from domesticated animals, e.g., goat, cow, and sheep.24 In human milk, the lacto-N-biose type 1 structural epitope predominates over those containing the N-acetyllactosamine type 2 structure (Figure 2), whereas type 1 chains are rare in milk of domesticated dairy animals. Contrary to hMOS, domesticated animal milk oligosaccharides can also have N-acetyllactosamine [Gal(β1–4)GlcNAc] at the reducing end.44 Around 70% of hMOS is neutral and often fucosylated,24 whereas the fraction of fucosylated MOS in animal milk is relatively low and most of the oligosaccharides are acidic.44,45 Domesticated animal milk can contain structural epitopes that are not present in human milk, such as GalNAc, Neu5Gc-containing oligosaccharides, and (α1–3)-linked Gal and GalNAc.24 Moreover, in goat as well as bovine milk oligosaccharides, Gal(β1–4)GlcNAc branches on 3′-galactosyllactose (lacto-N-novopentaose) have been observed as well as sialylation of both 3′-GL as well as 6′-GL (Schemes 1 and 2).
Figure 2.

Biosynthesis of neutral and acidic complex (left) human milk oligosaccharides (hMOS) and (right) goat milk oligosaccharides (gMOS). Enzymes probably involved are indicated in the human pathway. Required enzyme activities to achieve known structures are notated in the goat pathway. Iβ6GlcNAcTx refers to a potential separate enzyme as observed in marsupial lactation, capable of adding GlcNAc(β1–6) branching of 3′-GL.57 Main pathways and structure types are depicted in bold with thicker arrows. The human hypothetical pathway is compiled from Urashima et al.,8 Bode et al.,16 Blank et al.,54 and Van Leeuwen et al.53 and combined with the Kyoto Encyclopedia of Genes and Genomes (KEGG) glycosphingolipid biosynthesis: lacto and neolacto series.56
In bovine milk, N-linked glycans from glycoproteins were shown to contain N,N′-di-N-acetyllactose diamine [GalNAc(β1–4)GlcNAc (LacdiNAc)].46 Although no comprehensive studies of goat milk protein glycosylation are known to us, a recombinant human erythropoietin expressed in transgenic goat milk was highly decorated with the LacdiNAc epitope.47 In bovine milk, LacdiNAc disaccharide has been observed as well as other oligosaccharides where Gal has been substituted for GalNAc.38,48 This activity has been ascribed to bovine α-lactalbumin, which induces milk β-galactosyltransferase (B4GalT1) to accept uridine diphosphate (UDP)–GalNAc as a donor as well as UDP–Gal in the synthesis of lactose.49 It is not known whether this is also responsible for the LacdiNAc glycotope in protein glycosylation. Possibly β-N-acetylgalactosaminyltransferases B4GalNT3 and B4GalNT4 are responsible for the synthesis of these glycotopes. It should be noted that, in humans, these enzymes are also encoded in the genome and are responsible for the presence of LacdiNAc epitopes in the gastric mucosa.50 In view of the similarities between MOS and protein glycosylation patterns, such LacdiNAc glycotopes can also be expected to occur in bMOS and gMOS.
The α-Gal glycotope is found abundantly in mammalian glycosylation, except for humans, apes, and Old World monkeys.51 The responsible enzyme α-galactosyltransferase (GGTA1) is functionally encoded in the genomes of most mammals. In the human genome, two non-functional variants have been identified, GGTA1 and GGTA1P.52
From the structures known thus far (Schemes 1 and 2), it is possible to construct a hypothetical biosynthetic scheme for gMOS, as presented previously for hMOS (Figure 2).53,54 Although for gMOS the biosynthetic enzymes are less established, many of the enzymes postulated to be involved in hMOS biosynthesis have homologues in the goat genome that are also expressed in the mammary gland during lactation.55,56 There are some interesting differences, particularly in the presence of major levels of structures consisting of lactose elongated with (β1–3) galactose units. Also, the presence of the 6′ isotope of LNT (iLNT) in goat milk indicates that, contrary to the human biosynthetic pathway, the GlcNAc(β1–3)Gal epitope is not required for the attachment of a 6′-GlcNAc residue. Also, the structure Gal(β1–3)[GlcNAc(β1–6)]Gal(β1–4)Glc may be formed by transfer of Gal(β1–3) to iLNT or alternatively by transfer of GlcNAc by a separate Iβ6GlcNAcT enzyme, as observed in lactating tammar wallaby.57 The human biosynthetic pathway does not include enzymes for Gal(β1–3), with Gal(β1–4) disubstitution on a GlcNAc residue, which has been tentatively observed in goat milk.24 It should be noted that the first elucidation of this structure is tentative, because the papers identifying this structure initially contained mistakes in other structures. The goat structures indicate activity of ST8Sia, to generate Sia(α2–8)Sia epitopes, with either Neu5Ac or Neu5Gc residues. Activity of such ST8Sia transferases in the mammary gland of lactating goats was observed in a comprehensive transcriptomics study.55 Finally, the goat milk oligosaccharides contain some Gal(α1–3)Gal and GalNAc(α1–3)Gal glycotopes that are absent in human milk.
Quantitative Aspects of gMOS
Milk is a complex fluid, containing many classes of biomolecules (Figure 1). A major class is carbohydrates, of which the majority is usually lactose. The second class of carbohydrates in human milk is that of hMOS, while in bovine milk, this is monosaccharides, with bMOS constituting the third carbohydrate class.58,59 For goat milk, the concentrations of monosaccharides have not been extensively studied, but one study puts them at a comparable level with gMOS.34 Analysis of milk oligosaccharides often requires extraction of MOS prior to analysis. The extraction procedure and required level of purity depend upon the analytical methods, which are, in turn, determined by the research question. For identification of specific, unknown structures, the compounds need to be isolated in pure form for further analysis. A recent review outlines the challenges and pitfalls in analysis of hMOS.53 The most common methods for hMOS analysis have been described in detail here, and the same challenges hold for gMOS analysis. In short, many analytical methods have been employed, and particularly, comprehensive quantitative analysis is difficult. The risk of loss of MOS in the extraction phase is one major issue that has been understudied thus far. Also, the choice of analytical method determines to a large part the precision of quantitation. While for human milk many studies have been performed, a wide variation in quantitative results has been observed.3 Some of the variation can be explained by population-based differences60 or genetic variation between individuals.53 For goats, as well, the breed and geographical factors may influence the results. It is, however, also possible that differences in analytical approaches yield deviating results.
Several studies have been performed quantifying gMOS or comparing MOS quantities between domesticated animal species. In a comparative study between goat, sheep, and cow milk, quantification was based on standards using high-pressure/pH anion-exchange chromatography coupled with pulsed amperometric detection (HPAEC–PAD).29 They found that the mature Murciano-Granadina goat milk contained 250–300 mg/L gMOS, representing about 5 times the amount of bMOS measured in cow milk (30–60 mg/L) and about 10 times more than sMOS in sheep milk (20–40 mg/L) in the same study.29 Another study on this breed investigated the levels of gMOS in colostrum and mature milk of four individual goats as well as a pooled milk sample from 12 goats.37 A large variation in total gMOS levels was observed in colostrum of the four individual Murciano-Granadina goats and the pooled colostrum, ranging from 251 to 572 mg/L (Table 1).37 Total neutral gMOS levels ranged from 140 to 315 mg/L, and total acidic gMOS levels ranged from 83 to 251 mg/L. The most abundant gMOS were galactosyllactoses, with values ranging from 128.7 to 274.3 mg/L. Higher 2′-FL concentrations (2.2–31.6 mg/L) were measured in comparison to fucosyllactosamine (3.1–6.15 mg/L). Furthermore, the authors noted that, similar to human milk, all five colostrum samples contained higher 6′-SL concentrations (29–124 mg/L) than 3′-SL (3–12 mg/L).37 It should be noted that several other gMOS studies showed opposite findings, identifying predominant levels of 3′-SL instead.25,26,61 Also, in one study on human milk in Vietnam, specific individuals were identified with predominantly 3′-SL or near equal levels of 3′-SL and 6′-SL rather than a clear predominance of 6′-SL.62
Table 1. Overview of Average Levels of gMOS Determined in Quantitative Studies from Different Breeds, Murciano-Granadina (MG), New Zealand Saanen (NZS), Alpine (A), Garganica (G), Maltese (M), and Saanen (S).
| breed | gMOS (mg/L) | methoda | comments | reference |
|---|---|---|---|---|
| MG | 250–300 | HPAEC–PAD | (29) | |
| MG | 251–572 | HILIC–MS | acidic, 83–251 mg/L; neutral, 140–315 mg/L | (37) |
| NZS | 260–320 | LC–MS | colostrum, ∼320 mg/L; mature milk, ∼260 mg/L | (34) |
| A | 1110–1250 | GC–MS | monosaccharide-analysis-based method; high levels of Glc and Gal suggest significant levels of lactose | (27) |
| G | 240–700 | HPAEC–PAD | only 3′-SL, 6′-SL, and DSL; colostrum, 700 g/L; mature milk, 240 g/L | (25) |
| M | 190–625 | HPAEC–PAD | only 3′-SL, 6′-SL, and DSL; colostrum, 625 mg/L; mature milk, 190 g/L | (25) |
| S | 125–365 | HPAEC–PAD | only 3′-SL, 6′-SL, and DSL; colostrum, 365 g/L; mature milk, 125 g/L | (26) |
| MG | 112–488 | HILIC–MS | colostrum, 488 mg/L; mature milk, 112–178 mg/L | (28) |
| MG | 703–2398 | LC–MS | first 4 days of lactation; day 1, 2400 mg/L; day 4, 700 mg/L | (30) |
HPAEC–PAD, high-performance/pH anion-exchange chromatography–pulsed amperometric detection; GC–MS, gas chromatography–mass spectrometry; LC–MS, liquid chromatography–mass spectromtery; and HILIC–MS, hydrophilic interaction chromatography–mass spectrometry.
Other studies on gMOS concentrations in New Zealand Saanen goats showed ∼320 mg/L in colostrum and ∼260 mg/L in mature milk (Table 1).34 This study applied liquid chromatography with mass spectrometric detection (LC–MS), separating structures on a porous graphitized carbon (PGC) column. Quantitation was achieved in relation to a calibration curve of commercial standards (4′-GL, 3′-SL, 6′-SL, 3-FL, 2′-FL, LNH, DSL, lactose-1P, and GOS from Yakult). Only structures fitting m/z of these compounds were taken into account for the quantitation. This might result in some underestimation of gMOS, because other structural compositions were not taken into account.
Quantitation was also performed in a comparative study between Alpine goats that were either capable or incapable of producing αs1-casein.27 In the study, the milk of eight A/A goats (capable of producing αs1-casein) was compared to the milk from eight O/O goats (null mutant). The study observed a broad spread in concentrations for each group with 1110 ± 290 mg/L for A/A goats and 1250 ± 320 mg/L for O/O goats (Table 1) but no statistically significant difference between groups. These values are significantly higher than the values observed in the aforementioned studies,25,26,28,29,37 and the applied method relies on monosaccharide quantitation by gas chromatography coupled to MS, showing almost equivalent amounts (44.2–46.9 mol %) of Gal and Glc in all samples, suggesting a significant portion of remaining lactose as part of the gMOS fraction.27
A series of studies by Claps et al. analyzed the concentrations of 3′-SL, 6′-SL, and DSL in the colostrum and mature milk of Garganica (n = 15), Maltese (n = 20), and Saanen (n = 5) goats, using a HPAEC–PAD method with external standard calibration.25,26 Significant differences in the levels of the three sialyloligosaccharides were found between the breeds (Figure 3 and Table 1).
Figure 3.

Average levels (mg/L) of 3′-SL, 6′-SL, and DSL in milk of (A) Garganica, (B) Maltese, and (C) Saanen goats over time and adapted from Claps et al.25,26
Garganica colostrum and milk samples contained higher concentrations of 3′-SL and 6′-SL than Maltese and Saanen goat colostrum, whereas the DSL content was highest in colostrum from Saanen goats. In the first 24 h after kidding, a significant increase in the 3′-SL concentration in the milk of all three breeds was observed, followed by a decline toward mature milk.25,26 The same pattern was observed for 6′-SL concentrations for the three goat breeds. The DSL content in Garganica (137.9–126.4 mg/L) and Saanen (150.0–113.9 mg/L) goat colostrum dropped slightly in the first 24 h, from 137.9 to 126.4 mg/L and from 150.0 to 113.9 mg/L, respectively, but significantly increased in colostrum of Maltese goats, from 104 to 228 mg/L (Table 1). Toward mature milk, the DSL levels gradually decreased for all breeds (Figure 3). The general trend of decrease in MOS concentrations from colostrum toward mature milk is found in common also with hMOS and bMOS.22,63 A recent study comparing Saanen goats to local Guanzhong goats at 40–50 days of lactation showed similar levels of 3′-SL, 17.17 mg/L in Guanzhong and 18.51 mg/L in Saanen, while 6′-SL and DSL are significantly increased in Guanzhong goats, i.e., 33.41 mg/L 6′-SL versus 9.98 mg/L in Saanen goats and 1.16 mg/L DSL in Guanzhong versus 0.39 mg/L in Saanen.64 The Claps studies on Saanen goats showed much higher concentrations at 90 days postpartum.26
Saanen goats have a high frequency of defective alleles at the αs1-casein locus, and this is associated with a low αs1-casein content in milk (450 mg/L compared to up to 8.4 g/L).26,65 On the basis of these results, Claps et al. speculated that the defective αs1-casein production negatively affected the whole secretion process of milk constituents, including the synthesis of sialyltransferases and, thus, the production of sialylated gMOS. This is in contrast, however, with the findings from Meyrand et al., where no significant differences in acidic gMOS concentrations were observed between Alpine goats with and without αs1-casein deficiency.27
Claps and co-workers also investigated the influence of parity on the sialyl gMOS content in colostrum and milk.26 Goat milk samples from five goats in second parity and five goats in third parity were analyzed, showing only a significant difference in 3′-SL concentrations between second and third parity, with values of 203 and 125 mg/L, respectively.26 A recent study has shown that, in protein glycosylation of goat milk glycoproteins, fucosylation and sialylation increase with parity.66
Martín-Ortiz et al. examined gMOS levels in milk of Murciano-Granadina goats at different lactation stages.28 Eight major gMOS (3′-SL, 6′-SL, sialyllactosamine, three glycolylneuraminyllactose isomers, and two galactosyllactose isomers) were quantified with hydrophilic interaction chromatography–quadrupole–mass spectrometry (HILIC–Q–MS), and 49 gMOS structures were monitored.
A comparison between milk of a single goat and a pooled milk sample from eight goats showed a considerable difference in total gMOS between the individual goat (GM) and the pooled milk (GP) samples of 58.9 and 178.1 mg/L, respectively (Table 1).28 This large difference in results between the individual goat and the pooled milk shows the large variability in gMOS composition between individual goats of the same species. Similar to results from Claps et al., over time, concentrations of the most abundant gMOS were highest at the beginning of lactation and decreased with lactation time.25,26 In Murciano-Granadina goats, 6′-SL was by far the most abundant acidic gMOS in the pooled samples, whereas in the individual goat samples, 3′-SL levels almost reached those of 6′-SL. Also notably, 3′-SL in pooled milk of Saanen, Garganica, and Maltese goats was the most abundant gMOS.25,26
The concentration of neutral gMOS (265.2 mg/L) was higher than that of acidic gMOS (222.8 mg/L) in the beginning of lactation but rapidly dropped with lactation time, mainly as a result of a significant decrease in galactosyllactose isomers, the most abundant neutral gMOS. The acidic gMOS content also decreased with lactation time but not as much as that of the neutral gMOS, resulting in higher acidic (149.3 mg/L) than neutral (28.8 mg/L) gMOS concentration at 30 days of lactation. Notably, the concentrations of 3′-SL are much lower than those observed in the three other goat species, while the 6′-SL levels at day 1 are comparable to that of the Maltese goats. The levels of 6′-SL remain higher in the Murciano-Granadina goats. In this study, also three Neu5Gc–lactose isomers were monitored, without giving specific structures. These oligosaccharides were only observed at relatively low levels (together ∼25–50 mg/L) and do not seem to follow the same concentration trend over time as analogous compounds containing Neu5Ac. The concentrations of two neutral galactosyllactose structures, most likely 3′-GL and 6′-GL, were also measured, ranging from ∼30 to 270 mg/L; approximately half of the gMOS determined in this study were neutral. In the study by Albrecht et al., the analysis of acidic versus neutral structures showed 95% acidic structures in goat milk.24 Other reports also state that acidic oligosaccharides are the major component in goat milk.25,45 It is possible that the levels of acidic versus neutral gMOS are strongly affected by goat breed, suggesting the potential for a comprehensive and comparative study of goat milk from different breeds.
A recent study on goat colostrum composition analyzed lipid and oligosaccharide composition and concentration.30 This paper showed levels decreasing from 2398.4 mg/L at day 1 to 702.7 mg/L at day 4 of lactation, markedly higher than in other studies thus far (Table 1).
When all quantitative studies on gMOS are taken into account (Table 1 and Figure 4), a very broad range of concentrations has been reported. The lowest value measured in a single goat was 58.9 mg/L in mature milk and ∼200 mg/L in early colostrum.28 The highest values in mature milk were observed in a single study on Saanen goats at 1110–1250 mg/L.27 The methodology of this study, however, seems to have incorporated a significant amount of lactose into the quantitation and should therefore be discounted as an assessment of gMOS quantity. The highest colostrum values were ∼2.4 g/L in Murciano-Granadina goats.30 On average, the values reported range from ∼100 to 250 mg/L in mature milk and from ∼300 to 2400 mg/L in colostrum across different breeds. Interestingly, one study on bovine milk showed that the major oligosaccharides, i.e., 3′-SL, 6′-SL, and 6′-SLN, start at high concentrations in colostrum, i.e., 855 mg/L for 3′-SL, 117 mg/L for 6′-SL, and 141 mg/L for 6′-SLN, followed by a rapid decrease.67 An ultrafiltrate of whey permeate was shown to contain particularly high levels of bMOS, indicating that processing streams may be a promising source of dairy-derived MOS.68 A comparison of MOS composition of goat milk with human, bovine, and sheep milk shows that, in both the colostrum phase as well as mature milk phase, the quantities of gMOS are higher than those for bovine and sheep milk but still far lower than those in human milk (Figure 4). The physiological and molecular basis for these variations in gMOS concentrations remains to be elucidated. It should be noted that quantitative studies on hMOS also vary greatly, and recent recommendations were to evaluate different methods in a cross-laboratory study to determine how much of the variation is caused by experimental bias.3,53 A similar recommendation could be made for analysis of goat milk oligosaccharides.
Figure 4.

Comparison of acidic and neutral MOS concentration levels between human (blue), goat (green), cow (purple), and sheep (red) colostrum (Col) and mature milk (Mat). For sheep colostrum, there are no data (n.d.) available. The open part of the bars shows the highest concentration level reported, and the filled part of the bars represents the lowest reported concentration.3,25,26,29,30,37,60,63
Functional Analysis of Milk Oligosaccharides
Currently, increasing evidence is generated for specific functions of specific hMOS, being antiadhesive, immune-modulating, or a targeted prebiotic for specific desirable bacterial/microbiota strains in early infant development.16 At the moment, there is a lack of studies into the required concentrations of hMOS to achieve optimal functionality. Also missing in the current understanding is the level of diversity required for optimal health effects in the infant development. The addition of a single hMOS structure as a supplement to infant formula clearly has benefits, as observed for the additions of 2′-FL.69,70 When gMOS and bMOS composition is observed, it is clear that goat milk shows more similarities with hMOS composition than bMOS.24,27,29,37 It should be noted, however, that there is still a significant difference between gMOS and hMOS. One particular difference is the dominance of acidic MOS in goat milk compared to human milk (Figure 4). Moreover, the neutral fraction of hMOS is dominated with fucosylated structures, while gMOS contain only minor levels of fucosylated neutral structures (Figure 5). Figure 5 shows a comparison of structures and relative quantities based on the study by Albrecht et al. for goat milk24 to term and preterm human milk on the basis of the study by Austin et al.63 Although the composition of hMOS is significantly different from gMOS, there are sialylated, neutral non-fucosylated, and neutral fucosylated structures in common between the two. Therefore, in comparison to bMOS and sMOS, gMOS have more potential as a functional substitute for hMOS. Moreover, the levels of gMOS are significantly higher than those of bMOS and sMOS, suggesting that goat milk has more potential as a source for such functional oligosaccharides.
Figure 5.

Overview of relative intensities of gMOS derived from Albrecht et al.24 compared to hMOS in term and preterm milk derived from Austin et al.63 Clockwise from the top, first, acidic oligosaccharides are grouped together, presented according to relative abundance, followed by neutral oligosaccharides in order of abundance. Structures similar between human and goat milk are presented in bold, and corresponding segments are offset from the ring chart.
Prebiotic Potential
Thum and co-workers used a previously obtained gMOS-enriched fraction (labeled CMOF in their work)34 in a study to investigate the growth of bifidobacteria that were isolated from the feces from breast-fed infants.71 It should be noted that processing conditions and use of β-galactosidase in the enrichment of gMOS has some influence on the composition of the oligosaccharide mixture.72 A total of 17 strains of bifidobacteria were isolated from infant feces, and their growth on gMOS was analyzed in comparison to GOS, FOS, inulin, and 3′-SL or 6′-SL.71 All bifidobacteria grew faster and reached higher optical densities on the gMOS compared to the other substrates. No growth was observed on medium supplemented with inulin. Interestingly, one Bifidobacterium bifidum strain was able to use 3′-SL or 6′-SL as the sole carbon source, whereas the other B. bifidum strains were able to ferment the gMOS-enriched fraction while partially degrading 3′-SL and 6′-SL in the gMOS. Carbohydrate depletion after 36 h of incubation was evaluated by LC–MS and HPLC analysis. Two B. bifidum strains (AGR2166 and AGR2168) used more gMOS than Bifidobacterium breve or Bifidobacterium longum subspecies longum isolates, and this was related to an enhanced sialidase activity. Furthermore, for gMOS, the in vitro production of lactate and SCFAs, such as acetate, compared better to other fermentable substrates.71 These SCFAs are important substrates for gut health and gut microbiota.73,74 The production of SCFAs by beneficial bacteria has been linked to protection against several pathologies on the long term, including cardiovascular disease, inflammatory bowel disease (IBD), metabolic syndrome, and even some types cancer.75−77 Butyrate is an important SCFA and considered the preferred energy source for colonocytes.78 Incubation of B. longum subsp. infantis with gMOS has also shown increased adhesion to HT-29 cells of more than 8-fold compared to controls and more than 2.5-fold better than immunoglobulin G (IgG)-enriched bovine whey.61 The authors of that study used the same gMOS mixture, rich in 3′-SL, in a study showing enhanced SCFA production when B. longum susbsp. infantis is incubated with gMOS. Moreover, the pre-incubation with gMOS enhanced the ability of B. longum subsp. infantis to inhibit Campylobacter jejuni infection of HT-29 cells.79
The prebiotic potential of gMOS was confirmed in another study.80 Here, bifidobacteria Bifidobacterium animalis and B. longum strains were tested as well as Lactobacillus casei and Lactobacillus acidophilus strains. The growth rate of both bifidobacteria strains as well as L. casei was higher with gMOS isolated from goat-milk-derived infant formula than with the established GOS prebiotic (p < 0.05).80 It should be noted that both prebiotic studies have only tested probiotic species in monoculture and have not assessed growth of pathogens and/or commensal species nor assessed interspecies competition in mixed culture experiments. A shift toward more probiotic species in a mixed culture or in vivo is required to fully substantiate a prebiotic function.
An in vivo study by Thum et al. determined the effect of prenatal consumption of gMOS (CMOF) on the colon microbiota and milk composition of pregnant mice.81 The development of the mice offspring and whether these effects in offspring persisted after 30 days of consuming the control diet were also determined. The diet did not affect the food intake, body weight, gastrointestinal tract (GIT) length, small intestine length, or weight of stomach, colon, spleen, kidneys, brain, femur, and visceral fat of the dam. However, in comparison to negative-control-diet- and GOS-diet-fed dams, the gMOS-diet-fed dams had an increased colon length and lower liver weight at 30 days after delivery. The pups of the gMOS-fed dams had increased body weight and length at weaning, similar to GOS control pups. However, only pups of gMOS-fed dams had an increased microbial diversity and relative abundance of bifidobacteria at weaning as well as higher levels of the SCFA butyric acid in the colon. Bifidobacteria produce acetate and lactate, which can be converted into butyrate by other bacteria.82 It was concluded that consumption of gMOS by the dams during gestation and lactation improved the development of the offspring and the relative abundance of bifidobacteria in the colon at weaning. Although there is currently no data on human interventions with gMOS, studying the potential prebiotic effects, the studies thus far suggest prebiotic potential more closely resembling hMOS than GOS. This may be explained by the structural overlap between gMOS and hMOS (Figure 4). There is currently no intervention study with gMOS; a study with bMOS has been performed, using a bMOS enriched whey permeate, obtained through ultrafiltration and anion-exchange chromatography. The bMOS product is well-tolerated by healthy adults in a single-blind placebo-controlled study.83,84 The bMOS were fully digested by the intestinal microbiota but did not illicit a significant change in the microbial transcriptome nor on the exfoliated gut epithelia.84 There was only a short-term effect on bifidobacterial levels, showing an increase after 4 days of intervention but no longer after 11 days.83 The product contained mostly 3′-SL and to a lesser extent 6′-SL,85 fitting the composition observed for bovine milk.24 It should be noted that recent advances in hMOS synthesis technology have allowed for sufficient levels of 2′-FL and LNnT to be produced, leading to the first European Food Safety Authority (EFSA)-approved infant formulas supplemented with these hMOS for functionality. Tests thus far have shown that the addition of 2′-FL and LNnT shifts the microbiota more toward breast-fed compositions than formulas with GOS and FOS.86,87 Moreover, analysis of goat milk has shown the presence of both 2′-FL and LNnT, underscoring the potential of goat milk as an infant formula ingredient.
Anti-inflammatory Effects
Goat milk oligosaccharides have shown to be anti-inflammatory in a rat model of hapten-induced colitis. Daddaoua et al. investigated the role of gMOS in a model of experimental colon inflammation induced by the hapten, trinitrobenzenesulfonic acid (TNBS), in rats.88 TNBS-induced colon inflammation is characterized by anorexia with body weight loss, epithelial necrosis, bowel wall thickening, and colon shortening. For the treatment of rats, an enriched gMOS permeate of Martinez-Ferez et al. was used.89 This permeate contains ∼200–240 mg/L gMOS but also ∼2.25 g/L lactose. Rats that were fed with 500 mg of the lyophilized gMOS permeate/kg of body weight per day showed significantly less symptoms compared to control rats that did not receive the gMOS. Downregulation of colonic expression of inflammatory factors interleukin 1β (IL-1β) and inducible nitric oxide synthase (iNOS) was also observed as well as reduced cyclooxygenase 2 (COX2), which is not directly involved in inflammation but can be used as a marker for inflammation.88 Because the gMOS sample still contained a lot of lactose, it is unclear whether all of the observed effects are only caused by gMOS or if lactose also had an effect.88
Lara-Villoslada et al. also studied the effect of gMOS on colon inflammation in rats induced by dextran sodium sulfate (DSS).90 They also used the enriched gMOS permeate produced by Martinez-Ferez et al. but first removed the residual lactose from the gMOS fraction. The body weight gain and food intake of rats fed with gMOS (PF-GMO) were similar to the control rats that did not receive gMOS (PF-control). Interestingly, the colonic content of probiotic lactobacilli and bifidobacteria was significantly higher in the rats receiving gMOS but not of the potential pathogens. After DSS treatment, histological analysis showed that the gMOS rats (DSS-GMO) had no ulceration and recovered from inflammation, while the DSS control rats had significant ulceration and inflammation. Also, blood granulocyte levels were reduced in gMOS-fed rats compared to control rats. In gMOS-fed rats, the levels of myeloperoxidase activity, a proxy for neutrophil infiltration, do not increase upon DSS treatment, while in control rats, a 5-fold activity increase is observed upon DSS treatment.90
Both studies indicated that gMOS reduce intestinal inflammation and contributed to the recovery of damaged colonic mucosa and suggested gMOS as a suitable treatment of inflammatory bowel disease. Previous studies showed that GOS, which are commonly added to infant formula to substitute hMOS, were unable to reduce inflammation in a rat model for colitis.91 Similar studies have not been performed with hMOS thus far; however, anti-inflammatory effects have been well-established for hMOS.16,92
A recent mini-review showed an overview of hMOS with the potential for galectin binding and, thus, galectin-directed signaling.8 Many hMOS have potential galectin recognition epitopes, i.e., all structures with a non-reducing terminal galactose linked to a Glc, GlcNAc, or GalNAc residue, with free OH on Gal OH4 and OH6 and a free OH3 on the preceding Glc, GlcNAc, or GalNAc. Other glycotopes were shown to not fit galectin-binding sites.93 This interaction may be one of the mechanisms of hMOS immune modulation.8 Many of the structures shown in Schemes 1 and 2 have clear potential for galectin recognition, marked in green. Galectins play a pivotal role in immune modulation and inflammatory control.94 Therefore, many of the relatively abundant gMOS, such as 3′-GL, LNnT, LNnH, 3′-SL, and 3′-NGL (Schemes 1 and 2), have potential immune-modulating and anti-inflammatory effects through galectin interactions. Detailed studies on galectin specificity have shown, moreover, that some galectins have enhanced affinity when the Gal residue is sialylated on the 3 position, e.g., 3′-SL, while affinity for galectin 8-N is even further enhanced for ganglioside GD3, containing a Neu5Ac(α2–8)Neu5Ac(α2–3)Gal epitope.93 This epitope is also present on the gMOS structure DSL (Scheme 2). Specifically, a strong interaction of galectin 3-C for the Gal(α1–3)Gal and GalNAc(α1–3)Gal glycotopes has also been documented.93,95 Elevated serum levels of galectin 3 have been associated with long-term risks in cardiovascular disease.96 While human milk does not contain oligosaccharides with this glycotope, goat milk does contain a low amount of 3′-α-galactosyllactose (Scheme 1). Galectin 9-N and 9-C binding domains both interact strongly with poly-β1–3-linked Gal residues, which do not occur in human milk but are present in goat milk (Scheme 1). Where it was previously argued that hMOS might be a powerful tool to study galectin-binding kinetics and functionality,8 gMOS might fill some gaps in glycotopes that are absent in hMOS, further enhancing the understanding of galectin interactions. Similarly, gMOS might be applied to study the binding of other lectins, including siglecs, selectins, and DC-SIGN, but also bacterial and viral glycan adhesion molecules. In the gut, intestinal epithelial cell-derived galectin 9 has been implicated in GOS/FOS-associated immunomodulation.97,98 Galectins are involved in neutrophil regulation, and triggering of neutrophil response is implicated in necrosis in colitic inflammation.99 Stimulation of galectins by hMOS or gMOS in the gut might play a role in immune modulation and reducing necrotizing effects of inflammation.
Besides galectins, interactions between oligosaccharides and toll-like receptors (TLRs) in the gut have also been shown. In some studies, oligosaccharides, including GOS, hMOS, and gMOS, have been shown to mediate TLR-mediated response.100−102 In 2013, a mouse study showed that 3′-SL, which is also abundant in gMOS, interacts with TLR4, which is normally associated with LPS-mediated inflammatory response, leading to inflammation.103 Later, a study using human TLR4 was unable to reproduce the effect, concluding that differences in specificity of mouse and human TLR4 might be responsible.104
Barrier Function
Barnett et al. studied the effect of a semi-purified gMOS-enriched fraction of goat milk34 on the barrier integrity and mucin production of co-cultures of small and large intestine epithelial cells.105 They showed that the gMOS-enriched fraction enhanced the intestinal epithelial barrier function. The trans-epithelial electrical resistance (TEER) increased significantly and in a dose-dependent manner. Although the researchers did not separately investigate the effect of other components in the gMOS-enriched fraction, it is known that lactose, glucose, and galactose have no effect on TEER as a readout of barrier integrity.18,19 The gMOS did not significantly affect the metabolic flux of the epithelial cells. The expressions of the mucin-synthesizing genes for Muc2, Muc4, and Muc5AC were only slightly affected by the presence of gMOS, and total mucin production compared to controls was not different when cells were incubated with gMOS.105 The conclusion was that gMOS did increase the barrier function of intestinal cells but not through adaptation of metabolic flux or increased mucin production.
In a recent publication, different GOS formulations were investigated for their effect on mucin production and expression of mucin-related genes.100 This study observed that GOS with lactose had a more pronounced effect on the gene expression in cell lines than GOS that was purified of lactose. A further study using hMOS observed upregulation of mucin-production-related genes, MUC2, TFF3, and RETNLB, when LS174T cells were incubated with 3-FL, while MUC2 and TFF3 were upregulated by lacto-N-triose (LNT2).106 Furthermore, under challenge with tumor nectrosis factor α (TNF-α), interleukin 13 (IL-13), and tunicamycin, differential effects of 2′-FL, 3-FL, and LNT2 were observed, supporting a direct effect of these structures on barrier-function-related production of mucin.106 It should be noted that 3-FL, the most effective modulator, is not observed in goat milk, while LNT2 and 2′-FL have been found (Schemes 1 and 2); therefore, a certain level of induction of mucin production by gMOS may be expected.
A recent study using a murine NEC model observed that hMOS increased barrier function, by stimulating Muc2 production in the intestine.107 This was further verified in a human gut epithelial cell line. Furthermore, it was observed that the protective effect was mediated through chaperones, particularly protein–disulfide isomerase; suppression of protein–disulfide isomerase abolished the effect of hMOS.107 Lactose controls also had a noticeable effect on mucin-related gene expression. It seems that the effect of lowly digestible oligosaccharides on the mucin production is not straightforward.
Pathogen Inhibition
The pathogen inhibition activity of hMOS is presumed to be caused by similarity in structures between hMOS and epithelium-associated glycans.16 Pathogens present lectin-like receptors that recognize a specific glycan epitope to adhere to the epithelial layer as a first step in infection. In the presence of significant levels of hMOS with similar structural elements, hMOS will bind to these receptors and, thereby, prevent adhesion.108 In some cases, the hMOS block the epithelial receptor to which pathogen glycans adhere. For example, the HIV gp120 anchor protein presents high-mannose-type glycans that adhere to DC-SIGN as an adhesion mechanism for the pathogen.109 DC-SIGN has affinity for high-mannose as well as fucose-containing glycans, particularly glycans containing a Lex glycotope; i.e., Fuc(α1–3)[Galβ(1–4)]GlcNAc.110 Human milk contains LNFP III, with a Lex glycotope, as well as Muc1, containing glycans with Lex glycotopes that bind to DC-SIGN, thereby preventing viral adhesion.111−113 There is also evidence that 3′-SL directly interacts with epithelial cells, modulating gene expression of sialyltransferases, thereby modifying epithelial glycosylation.114 It has been hypothesized that the altered glycosylation might result in reduced binding of sialic-acid-specific pathogens.16 The large diversity of hMOS is thought to exist because of the large diversity in pathogens.16,115 It has been shown for N-glycans as well as hMOS that not only is a specific glycotope required for action, but also that the presentation of the epitope matters.116,117 The levels of hMOS are also relevant, because some interactions require multivalency.118 This can be achieved by not only highly complex, branched carbohydrates but also high levels of smaller oligosaccharides, resulting in indirect multivalency. This theory was further supported by research showing that the same functional epitope on different scaffold hMOS has different affinities for pathogen receptors.119 Well-established structures related to decoy effects against pathogens are collected in Table 2. Although it has not been established as a pathogen inhibition effect, a microbial imbalance or pathogenic species is most likely involved in the development of necrotizing enterocolitis. It is worth noting that the most profound effects have been observed with disialyl-LNT (DSLNT), which has not been observed in goat milk. However, protective effects against necrotizing enterocolitis have also been shown for 2′-FL and sialylated GOS.120 While human milk contains only minor levels of the GOS trisaccharides 3′-GL and 6′-GL43,121,122 and no sialylated GOS, goat milk does contain a wider array of GOS structures and several sialylated GOS structures (Scheme 2). These may potentially have similar effects, although the structural overlap with the sialylated GOS used in the necrotizing enterocolitis study is limited.123
Table 2. Structural MOS Epitopes with Shown Pathogen Inhibition Functionalitiesa.
| glycotopeb | pathogen | possible gMOS | reference |
|---|---|---|---|
| Fuc(α1–2)Gal glycotopes | Campylobacter jejuni | maybe | (125) |
| Candida albicans | maybe | (126) | |
| norovirus | maybe | (127) | |
| 3-FL | Escherichia coli | yes | (128) |
| Salmonella fyris | yes | (128) | |
| Lex (LNFP III)b | HIV | maybe | (111) |
| LNT | Entamoeba histolytica | no | (129) |
| Group B Streptococcus | no | (130−132) | |
| Streptococcus pneumoniae | no | (133) | |
| Pseudomonas aeruginosa | no | (130) | |
| LNFP I | Group B Streptococcus | no | (131,132) |
| LNnT | Streptococcus pneumoniae | yes | (133,134) |
| Pseudomonas aeruginosa | yes | (130) | |
| 3′-SL | Escherichia coli | yes | (128) |
| influenza B | yes | (130) | |
| Helicobacter pylori | yes | (135) | |
| Pseudomonas aeruginosa | yes | (136) | |
| 6′-SL | influenza A | yes | (130) |
| Salmonella fyris | yes | (128) | |
| Escherichia coli | yes | (128) | |
| Pseudomonas aeruginosa | yes | (136) | |
| general hMOS | Vibrio cholera | maybe | (128) |
The presence of a certain glycotope in gMOS is provided.
3-FL, 3-fucosyllactose; LNT, lacto-N-tetraose; LNFP I, lacto-N-fucopentaose I; LNnT, lacto-N-neotetraose; 3′-SL, 3′-sialyllactose; and 6′-SL, 6′-sialyllactose.
Recently, it was shown that gMOS can trigger bifidobacteria to inhibit C. jejuni infection in an in vitro study, showing a form of indirect pathogen inhibition.79 Two recent studies have investigated the direct antiadhesive capacity of isolated gMOS. One study observed reduced adhesion of Escherichia coli and Salmonella typhimurium to CACO-2 cells when pre-incubated with gMOS.80 This was observed independent of beneficial microbiota. The second study showed the same results with Salmonella by green fluorescent antibodies against the Salmonella strain used.124 Here, the effect of sialylated gMOS was shown to be stronger than that of neutral gMOS. Although not all structures in Table 2 are present in similar quantities in goat milk as in human milk, hypothetically, some functionalities may also occur with other gMOS if they present the suitable glycotopes as well as with hMOS. Further research into functionality of hMOS as well as non-human MOS would be beneficial for the development of the field.
Glossary
Abbreviations Used
- 2-AA
anthranilic acid
- 2-AB
2-aminobenzamide
- 2′-FL
2′-fucosyllactose, Fuc(α1–2)Gal(β1–4)Glc
- 3-FL
3-fucosyllactose, Gal(β1–4)[Fuc(α1–3)]Glc
- 3′-GL
3′-galactosyllactose, Gal(β1–3)Gal(β1–4)Glc
- 3′-NGL
3′-N-glycolylneuraminyllactose, Neu5Gc(α2–3)Gal(β1–4)Glc
- 3′-SHL
3′-sialylhexosyllactose, Gal(β1–6)[Neu5Ac(α2–3)]Gal(β1–4)Glc
- 3′-SL
3′-sialyllactose, Neu5Ac(α2–3)Gal(β1–4)Glc
- 4′-GL
4′-galactosyllactose, Gal(β1–4)Gal(β1–4)Glc
- 6′-GL
6′-galactosyllactose, Gal(β1–6)Gal(β1–4)Glc
- 6′-NGL
6′-N-glycolylneuraminyllactose, Neu5Gc(α2–6)Gal(β1–4)Glc
- 6′-SHL
6′-sialylhexosyllactose, Gal(β1–3)[Neu5Ac(α2–6)]Gal(β1–4)Glc
- 6′-SL
6′-sialyllactose, Neu5Ac(α2–6)Gal(β1–4)Glc
- 6′-SLN
6′-sialyllactosamine, Neu5Ac(α2–6)Gal(β1–4)GlcNAc
- B4GalNT
β-1,4-N-acetylgalactosaminyltransferase
- B4GalT
β-galactosyltransferase
- bMOS
bovine milk oligosaccharide(s)
- COX2
cyclooxygenase 2
- DC-SIGN
dentritic cell-specific intercelular adhesion molecule
- DSL
disialyllactose, Neu5Ac(α2–8)Neu5Ac(α2–3)Gal(β1–4)Glc
- DSLNT
disialyllacto-N-tetraose, Neu5Ac(α2–3)Gal(β1–3)[Neu5Ac(α2–6)]GlcNAc(β1–3)Gal(β1–4)Glc
- DSS
dextran sodium sulfate
- FOS
fructooligosaccharide(s)
- Fuc
fucose
- Gal
galactose
- GalNAc
N-acetylgalactosamine
- GGTA1
α-galactosyltransferase
- GIT
gastrointestinal tract
- Glc
glucose
- GlcNAc
N-acetylglucosamine
- gMOS
goat milk oligosaccharide(s)
- GOS
galactooligosaccharide(s)
- Hex
hexose
- HexNAc
N-acetylhexosamine
- HILIC–Q–MS
hydrophilic interaction chromatography–quadrupole–mass spectrometry
- HIV
human immunodeficiency virus
- hMOS
human milk oligosaccharide(s)
- HPAEC–PAD
high-pressure/pH anion-exchange chromatography–pulsed amperometric detection
- HPLC
high-pressure liquid chromatography
- IBD
inflammatory bowel disease
- IL-13
interleukin 13
- IL-1β
interleukin 1β
- iLNT
isolacto-N-tetraose, Gal(β1–3)GlcNAc(β1–6)Gal(β1–4)Glc
- iNOS
inducible nitric oxide synthase
- IβGlcNAcT
β-1,6-N-acetylglucosaminyltransferase
- LacdiNAc
N,N′-di-N-acetyllactose diamine, GalNAc(β1–4)GlcNAc
- LC–MS
liquid chromatography–mass spectrometry
- Lex
Lewis x blood group determinant
- LNFP III
lacto-N-fucopentaose III, Gal(β1–4)[Fuc(α1–3)]GlcNAc(β1–3)Gal(β1–4)Glc
- LNH
lacto-N-hexaose, Gal(β1–4)GlcNAc(β1–6)[Gal(β1–3)GlcNAc(β1–3)]Gal(β1–4)Glc
- LNnH
lacto-N-neohexaose, Gal(β1–4)GlcNAc(β1–6)[Gal(β1–4)GlcNAc(β1–3)]Gal,(β1–4)Glc
- LNnT
lacto-N-neotetraose, Gal(β1–4)GlcNAc(β1–3)Gal(β1–4)Glc
- LNT
lacto-N-tetraose, Gal(β1–3)GlcNAc(β1–3)Gal(β1–4)Glc
- LNT2
lacto-N-triose
- LPS
lipopolysaccharide
- MOS
milk oligosaccharide(s)
- MS
mass spectrometry
- Muc
mucin
- MUC2
mucin 2
- NEC
necrotizing enterocolitis
- Neu5Ac
N-acetylneuraminic acid
- Neu5Gc
N-glycolylneuraminic acid
- PGC
porous graphitized carbon
- SCFA
short-chain fatty acid
- sMOS
sheep milk oligosaccharide(s)
- ST8Sia
α-2,8-sialyltransferase
- TEER
trans-epithelial electrical resistance
- TFF3
trefoil factor 3
- TLR
toll-like receptor
- TNF-α
tumor nectrosis factor α
- TNSB
trinitrobenzenzesulfonic acid
- UDP
uridine diphosphate
- α3′-GL
3′α-galactosyllactose, Gal(α1–3)Gal(β1–4)Glc
Author Contributions
† Sander S. van Leeuwen and Evelien M. te Poele contributed equally to this work.
The authors thankfully acknowledge the financial support received from Ausnutria BV to the University of Groningen and CarbExplore Research BV. Part of the work was funded by the TKI Project “Influence of Human Milk Oligosaccharides in Early Life Development” (UMCG).
The authors declare the following competing financial interest(s): Eric Benjamins and Alfred Haandrikman are employed by Ausnutria BV. Evelien M. te Poele, Anastasia Chrysovalantou Chatziioannou, and Lubbert Dijkhuizen are employed by CarbExplore Research BV, which has received financial support from Ausnutria BV, at least partly used to fund the work of Sander S. van Leeuwen.
References
- Ballard O.; Morrow A. L. Human Milk Composition: Nutrients and Bioactive Factors. Pediatr. Clin. North Am. 2013, 60 (1), 49–74. 10.1016/j.pcl.2012.10.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meredith-Dennis L.; Xu G.; Goonatilleke E.; Lebrilla C. B.; Underwood M. A.; Smilowitz J. T. Composition and Variation of Macronutrients, Immune Proteins, and Human Milk Oligosaccharides in Human Milk From Nonprofit and Commercial Milk Banks. J. Hum. Lact. 2018, 34 (1), 120–129. 10.1177/0890334417710635. [DOI] [PubMed] [Google Scholar]
- Thurl S.; Munzert M.; Boehm G.; Matthews C.; Stahl B. Systematic Review of the Concentrations of Oligosaccharides in Human Milk. Nutr. Rev. 2017, 75 (11), 920–933. 10.1093/nutrit/nux044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kondyli E.; Svarnas C.; Samelis J.; Katsiari M. C. Chemical Composition and Microbiological Quality of Ewe and Goat Milk of Native Greek Breeds. Small Rumin. Res. 2012, 103 (2–3), 194–199. 10.1016/j.smallrumres.2011.09.043. [DOI] [Google Scholar]
- Park Y. W.Goat Milk—Chemistry and Nutrition. In Handbook of Milk of Non-bovine Mammals; Park Y. W., Haenlein G. F. W., Wendorff W. L., Eds.; John Wiley & Sons, Ltd.: Oxford, U.K., 2017; Chapter 2.2, pp 42–83, 10.1002/9781119110316.ch2.2. [DOI] [Google Scholar]
- Heck J. M. L.; van Valenberg H. J. F.; Dijkstra J.; van Hooijdonk A. C. M. Seasonal Variation in the Dutch Bovine Raw Milk Composition. J. Dairy Sci. 2009, 92 (10), 4745–4755. 10.3168/jds.2009-2146. [DOI] [PubMed] [Google Scholar]
- Kunz C.; Meyer C.; Collado M. C.; Geiger L.; García-Mantrana I.; Bertua-Ríos B.; Martínez-Costa C.; Borsch C.; Rudloff S. Influence of Gestational Age, Secretor, and Lewis Blood Group Status on the Oligosaccharide Content of Human Milk. J. Pediatr. Gastroenterol. Nutr. 2017, 64 (5), 789–798. 10.1097/MPG.0000000000001402. [DOI] [PubMed] [Google Scholar]
- Urashima T.; Hirabayashi J.; Sato S.; Kobata A. Human Milk Oligosaccharides as Essential Tools for Basic and Application Studies on Galectins. Trends Glycosci. Glycotechnol. 2018, 30 (172), SE51–SE65. 10.4052/tigg.1734.1SE. [DOI] [Google Scholar]
- Thurl S.; Henker J.; Siegel M.; Tovar K.; Sawatzki G. Detection of Four Human Milk Groups with Respect to Lewis Blood Group Dependent Oligosaccharides. Glycoconjugate J. 1997, 14 (7), 795–799. 10.1023/A:1018529703106. [DOI] [PubMed] [Google Scholar]
- Austin S.; de Castro C. A.; Bénet T.; Hou Y.; Sun H.; Thakkar S. K.; Vinyes-Pares G.; Zhang Y.; Wang P. Temporal Change of the Content of 10 Oligosaccharides in the Milk of Chinese Urban Mothers. Nutrients 2016, 8 (6), 346. 10.3390/nu8060346. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Samuel T. M.; Binia A.; de Castro C. A.; Thakkar S. K.; Billeaud C.; Agosti M.; Al-Jashi I.; Costeira M. J.; Marchini G.; Martínez-Costa C.; Picaud J.-C.; Stiris T.; Stoicescu S.-M.; Vanpeé M.; Domellöf M.; Austin S.; Sprenger N. Impact of Maternal Characteristics on Human Milk Oligosaccharide Composition over the First 4 Months of Lactation in a Cohort of Healthy European Mothers. Sci. Rep. 2019, 9 (1), 11767. 10.1038/s41598-019-48337-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thurl S.; Munzert M.; Henker J.; Boehm G.; Müller-Werner B.; Jelinek J.; Stahl B. Variation of Human Milk Oligosaccharides in Relation to Milk Groups and Lactational Periods. Br. J. Nutr. 2010, 104 (9), 1261–1271. 10.1017/S0007114510002072. [DOI] [PubMed] [Google Scholar]
- Tonon K. M.; de Morais M. B.; Abrão A. C. F. V.; Miranda A.; Morais T. B. Maternal and Infant Factors Associated with Human Milk Oligosaccharides Concentrations According to Secretor and Lewis Phenotypes. Nutrients 2019, 11 (6), 1358. 10.3390/nu11061358. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Azad M. B.; Robertson B.; Atakora F.; Becker A. B.; Subbarao P.; Moraes T. J.; Mandhane P. J.; Turvey S. E.; Lefebvre D. L.; Sears M. R.; Bode L. Human Milk Oligosaccharide Concentrations Are Associated with Multiple Fixed and Modifiable Maternal Characteristics, Environmental Factors, and Feeding Practices. J. Nutr. 2018, 148 (11), 1733–1742. 10.1093/jn/nxy175. [DOI] [PubMed] [Google Scholar]
- Gourbeyre P.; Denery S.; Bodinier M. Probiotics, Prebiotics, and Synbiotics: Impact on the Gut Immune System and Allergic Reactions. J. Leukocyte Biol. 2011, 89 (5), 685–695. 10.1189/jlb.1109753. [DOI] [PubMed] [Google Scholar]
- Bode L. Human Milk Oligosaccharides: Every Baby Needs a Sugar Mama. Glycobiology 2012, 22 (9), 1147–1162. 10.1093/glycob/cws074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Plaza-Díaz J.; Fontana L.; Gil A. Human Milk Oligosaccharides and Immune System Development. Nutrients 2018, 10 (8), 1038. 10.3390/nu10081038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Akbari P.; Braber S.; Alizadeh A.; Verheijden K. A. T.; Schoterman M. H. C.; Kraneveld A. D.; Garssen J.; Fink-Gremmels J. Galacto-Oligosaccharides Protect the Intestinal Barrier by Maintaining the Tight Junction Network and Modulating the Inflammatory Responses after a Challenge with the Mycotoxin Deoxynivalenol in Human Caco-2 Cell Monolayers and B6C3F1Mice. J. Nutr. 2015, 145 (7), 1604–1613. 10.3945/jn.114.209486. [DOI] [PubMed] [Google Scholar]
- Akbari P.; Fink-Gremmels J.; Willems R. H. A. M. A. M.; Difilippo E.; Schols H. A.; Schoterman M. H. C. C.; Garssen J.; Braber S. Characterizing Microbiota-Independent Effects of Oligosaccharides on Intestinal Epithelial Cells: Insight into the Role of Structure and Size. Eur. J. Nutr. 2017, 56 (5), 1919–1930. 10.1007/s00394-016-1234-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bermudez-Brito M.; Sahasrabudhe N. M.; Rösch C.; Schols H. A.; Faas M. M.; de Vos P. The Impact of Dietary Fibers on Dendritic Cell Responses in Vitro Is Dependent on the Differential Effects of the Fibers on Intestinal Epithelial Cells. Mol. Nutr. Food Res. 2015, 59 (4), 698–710. 10.1002/mnfr.201400811. [DOI] [PubMed] [Google Scholar]
- Perdijk O.; van Baarlen P.; Fernandez-Gutierrez M. M.; van den Brink E.; Schuren F. H. J.; Brugman S.; Savelkoul H. F. J.; Kleerebezem M.; van Neerven R. J. J. Sialyllactose and Galactooligosaccharides Promote Epithelial Barrier Functioning and Distinctly Modulate Microbiota Composition and Short Chain Fatty Acid Production In Vitro. Front. Immunol. 2019, 10, 10. 10.3389/fimmu.2019.00094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fong B. Y.; Ma K.; McJarrow P. Quantification of Bovine Milk Oligosaccharides Using Liquid Chromatography–Selected Reaction Monitoring–Mass Spectrometry. J. Agric. Food Chem. 2011, 59 (18), 9788–9795. 10.1021/jf202035m. [DOI] [PubMed] [Google Scholar]
- Aldredge D. L.; Geronimo M. R.; Hua S.; Nwosu C. C.; Lebrilla C. B.; Barile D. Annotation and Structural Elucidation of Bovine Milk Oligosaccharides and Determination of Novel Fucosylated Structures. Glycobiology 2013, 23 (6), 664–676. 10.1093/glycob/cwt007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Albrecht S.; Lane J. A.; Mariño K.; Al Busadah K. A.; Carrington S. D.; Hickey R. M.; Rudd P. M. A Comparative Study of Free Oligosaccharides in the Milk of Domestic Animals. Br. J. Nutr. 2014, 111 (7), 1313–1328. 10.1017/S0007114513003772. [DOI] [PubMed] [Google Scholar]
- Claps S.; Di Napoli M. A.; Sepe L.; Caputo A. R.; Rufrano D.; Di Trana A.; Annicchiarico G.; Fedele V. Sialyloligosaccharides Content in Colostrum and Milk of Two Goat Breeds. Small Rumin. Res. 2014, 121 (1), 116–119. 10.1016/j.smallrumres.2013.12.024. [DOI] [Google Scholar]
- Claps S.; Di Napoli M. A.; Caputo A. R.; Rufrano D.; Sepe L.; Di Trana A. Factor Affecting the 3′ Sialyllactose, 6’ Sialyllactose and Disialyllactose Content in Caprine Colostrum and Milk: Breed and Parity. Small Rumin. Res. 2016, 134, 8–13. 10.1016/j.smallrumres.2015.11.002. [DOI] [Google Scholar]
- Meyrand M.; Dallas D. C.; Caillat H.; Bouvier F.; Martin P.; Barile D. Comparison of Milk Oligosaccharides between Goats with and without the Genetic Ability to Synthesize As1-Casein. Small Rumin. Res. 2013, 113 (2–3), 411–420. 10.1016/j.smallrumres.2013.03.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martín-Ortiz A.; Barile D.; Salcedo J.; Moreno F. J.; Clemente A.; Ruiz-Matute A. I.; Sanz M. L. Changes in Caprine Milk Oligosaccharides at Different Lactation Stages Analyzed by High Performance Liquid Chromatography Coupled to Mass Spectrometry. J. Agric. Food Chem. 2017, 65 (17), 3523–3531. 10.1021/acs.jafc.6b05104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martinez-Ferez A.; Rudloff S.; Guadix A.; Henkel C. a.; Pohlentz G.; Boza J. J.; Guadix E. M.; Kunz C. Goats’ Milk as a Natural Source of Lactose-Derived Oligosaccharides: Isolation by Membrane Technology. Int. Dairy J. 2006, 16 (2), 173–181. 10.1016/j.idairyj.2005.02.003. [DOI] [Google Scholar]
- Marziali S.; Guerra E.; Cerdán-Garcia C.; Segura-Carretero A.; Caboni M. F.; Verardo V. Effect of Early Lactation Stage on Goat Colostrum: Assessment of Lipid and Oligosaccharide Compounds. Int. Dairy J. 2018, 77, 65–72. 10.1016/j.idairyj.2017.09.004. [DOI] [Google Scholar]
- Urashima T.; Bubb W. A.; Messer M.; Tsuji Y.; Taneda Y. Studies of the Neutral Trisaccharides of Goat (Capra hircus) Colostrum and of the One- and Two-Dimensional 1H and 13C NMR Spectra of 6′-N-Acetylglucosaminyllactose. Carbohydr. Res. 1994, 262 (2), 173–184. 10.1016/0008-6215(94)84177-2. [DOI] [PubMed] [Google Scholar]
- Viverge D.; Grimmonprez L.; Solere M. Chemical Characterization of Sialyl Oligosaccharides Isolated from Goat (Capra Hircus) Milk. Biochim. Biophys. Acta, Gen. Subj. 1997, 1336 (2), 157–164. 10.1016/S0304-4165(97)00021-4. [DOI] [PubMed] [Google Scholar]
- Urashima T.; Murata S.; Nakamura T. Structural Determination of Monosialyl Trisaccharides Obtained from Caprine Colostrum. Comp. Biochem. Physiol., Part B: Biochem. Mol. Biol. 1997, 116 (4), 431–435. 10.1016/S0305-0491(96)00269-6. [DOI] [PubMed] [Google Scholar]
- Thum C.; Cookson A.; McNabb W. C.; Roy N. C.; Otter D. Composition and Enrichment of Caprine Milk Oligosaccharides from New Zealand Saanen Goat Cheese Whey. J. Food Compos. Anal. 2015, 42, 30–37. 10.1016/j.jfca.2015.01.022. [DOI] [Google Scholar]
- Dong X.; Zhou S.; Mechref Y. LC-MS/MS Analysis of Permethylated Free Oligosaccharides and N-Glycans Derived from Human, Bovine, and Goat Milk Samples. Electrophoresis 2016, 37 (11), 1532–1548. 10.1002/elps.201500561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Sousa Y. R. F.; da Silva Vasconcelos M. A.; Costa R. G.; de Azevedo Filho C. A.; de Paiva E. P.; Queiroga R. de C. R. do E. Sialic Acid Content of Goat Milk during Lactation. Livest. Sci. 2015, 177, 175–180. 10.1016/j.livsci.2015.04.005. [DOI] [Google Scholar]
- Martín-Ortiz A.; Salcedo J.; Barile D.; Bunyatratchata A.; Moreno F. J.; Martin-García I.; Clemente A.; Sanz M. L.; Ruiz-Matute A. I. Characterization of Goat Colostrum Oligosaccharides by Nano-Liquid Chromatography on Chip Quadrupole Time-of-Flight Mass Spectrometry and Hydrophilic Interaction Liquid Chromatography-Quadrupole Mass Spectrometry. J. Chromatogr. A 2016, 1428, 143–153. 10.1016/j.chroma.2015.09.060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mariño K.; Lane J. A.; Abrahams J. L.; Struwe W. B.; Harvey D. J.; Marotta M.; Hickey R. M.; Rudd P. M. Method for Milk Oligosaccharide Profiling by 2-Aminobenzamide Labeling and Hydrophilic Interaction Chromatography. Glycobiology 2011, 21 (10), 1317–1330. 10.1093/glycob/cwr067. [DOI] [PubMed] [Google Scholar]
- Varki A. Uniquely Human Evolution of Sialic Acid Genetics and Biology. Proc. Natl. Acad. Sci. U. S. A. 2010, 107 (Supplement 2), 8939–8946. 10.1073/pnas.0914634107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bardor M.; Nguyen D. H.; Diaz S.; Varki A. Mechanism of Uptake and Incorporation of the Non-Human Sialic Acid N-Glycolylneuraminic Acid into Human Cells. J. Biol. Chem. 2005, 280 (6), 4228–4237. 10.1074/jbc.M412040200. [DOI] [PubMed] [Google Scholar]
- Quin C.; Vicaretti S. D.; Mohtarudin N. A.; Garner A. M.; Vollman D. M.; Gibson D. L.; Zandberg W. F. Influence of Sulfonated and Diet-Derived Human Milk Oligosaccharides on the Infant Microbiome and Immune Markers. J. Biol. Chem. 2020, 295 (12), 4035–4048. 10.1074/jbc.RA119.011351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alisson-Silva F.; Kawanishi K.; Varki A. Human Risk of Diseases Associated with Red Meat Intake: Analysis of Current Theories and Proposed Role for Metabolic Incorporation of a Non-Human Sialic Acid. Mol. Aspects Med. 2016, 51, 16–30. 10.1016/j.mam.2016.07.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Austin S.; Bénet T. Quantitative Determination of Non-Lactose Milk Oligosaccharides. Anal. Chim. Acta 2018, 1010, 86–96. 10.1016/j.aca.2017.12.036. [DOI] [PubMed] [Google Scholar]
- Urashima T.; Saito T.; Nakamura T.; Messer M. Oligosaccharides of Milk and Colostrum in Non-Human Mammals. Glycoconjugate J. 2001, 18 (5), 357–371. 10.1023/A:1014881913541. [DOI] [PubMed] [Google Scholar]
- Urashima T.; Taufik E.; Fukuda K.; Asakuma S. Recent Advances in Studies on Milk Oligosaccharides of Cows and Other Domestic Farm Animals. Biosci., Biotechnol., Biochem. 2013, 77 (3), 455–466. 10.1271/bbb.120810. [DOI] [PubMed] [Google Scholar]
- van Leeuwen S. S.; Schoemaker R. J. W.; Timmer C. J. a M.; Kamerling J. P.; Dijkhuizen L. Use of Wisteria Floribunda Agglutinin Affinity Chromatography in the Structural Analysis of the Bovine Lactoferrin N-Linked Glycosylation. Biochim. Biophys. Acta, Gen. Subj. 2012, 1820 (9), 1444–1455. 10.1016/j.bbagen.2011.12.014. [DOI] [PubMed] [Google Scholar]
- Montesino R.; Toledo J. R.; Sánchez O.; Sánchez A.; Harvey D. J.; Royle L.; Dwek R. A.; Rudd P. M.; Gerwig G. J.; Kamerling J. P.; Cremata J. A. Monosialylated Biantennary N-Glycoforms Containing GalNAc-GlcNAc Antennae Predominate When Human EPO Is Expressed in Goat Milk. Arch. Biochem. Biophys. 2008, 470 (2), 163–175. 10.1016/j.abb.2007.11.019. [DOI] [PubMed] [Google Scholar]
- Saito T.; Itoh T.; Adachi S. Presence of Two Neutral Disaccharides Containing N-Acetylhexosamine in Bovine Colostrum as Free Forms. Biochim. Biophys. Acta, Gen. Subj. 1984, 801 (1), 147–150. 10.1016/0304-4165(84)90223-X. [DOI] [PubMed] [Google Scholar]
- Do K.-Y.; Do S.-I.; Cummings R. D. α-Lactalbumin Induces Bovine Milk B1,4-Galactosyltransferase to Utilize UDP-GalNAc. J. Biol. Chem. 1995, 270 (31), 18447–18451. 10.1074/jbc.270.31.18447. [DOI] [PubMed] [Google Scholar]
- Ikehara Y.; Sato T.; Niwa T.; Nakamura S.; Gotoh M.; Ikehara S. K.; Kiyohara K.; Aoki C.; Iwai T.; Nakanishi H.; Hirabayashi J.; Tatematsu M.; Narimatsu H. Apical Golgi Localization of N, N′-Diacetyllactosediamine Synthase, B4GalNAc-T3, Is Responsible for LacdiNAc Expression on Gastric Mucosa. Glycobiology 2006, 16 (9), 777–785. 10.1093/glycob/cwl005. [DOI] [PubMed] [Google Scholar]
- Galili U. The α-Gal Epitope and the Anti-Gal Antibody in Xenotransplantation and in Cancer Immunotherapy. Immunol. Cell Biol. 2005, 83 (6), 674–686. 10.1111/j.1440-1711.2005.01366.x. [DOI] [PubMed] [Google Scholar]
- Shaper N. L.; Lin S.; Joziasse D. H.; Kim D.; Yang-Feng T. L. Assignment of Two Human α-1,3-Galactosyltransferase Gene Sequences (GGTA1 and GGTA1P) to Chromosomes 9q33–Q34 and 12q14–Q15. Genomics 1992, 12 (3), 613–615. 10.1016/0888-7543(92)90458-5. [DOI] [PubMed] [Google Scholar]
- van Leeuwen S. S. Challenges and Pitfalls in Human Milk Oligosaccharide Analysis. Nutrients 2019, 11 (11), 2684. 10.3390/nu11112684. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blank D.; Dotz V.; Geyer R.; Kunz C. Human Milk Oligosaccharides and Lewis Blood Group: Individual High-Throughput Sample Profiling to Enhance Conclusions From Functional Studies. Adv. Nutr. 2012, 3 (3), 440S–449S. 10.3945/an.111.001446. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crisà A.; Ferrè F.; Chillemi G.; Moioli B. RNA-Sequencing for Profiling Goat Milk Transcriptome in Colostrum and Mature Milk. BMC Vet. Res. 2016, 12 (1), 1–21. 10.1186/s12917-016-0881-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kanehisa M.; Furumichi M.; Tanabe M.; Sato Y.; Morishima K. KEGG: New Perspectives on Genomes, Pathways, Diseases and Drugs. Nucleic Acids Res. 2017, 45 (D1), D353–D361. 10.1093/nar/gkw1092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Urashima T.; Messer M.; Bubb W. A. Biosynthesis of Marsupial Milk Oligosaccharides II: Characterization of a β6-N-Acetylglucosaminyltransferase in Lactating Mammary Glands of the Tammar Wallaby, Macropus eugenii. Biochim. Biophys. Acta, Gen. Subj. 1992, 1117 (2), 223–231. 10.1016/0304-4165(92)90083-7. [DOI] [PubMed] [Google Scholar]
- Urashima T.; Asakuma S.; Leo F.; Fukuda K.; Messer M.; Oftedal O. T. The Predominance of Type I Oligosaccharides Is a Feature Specific to Human Breast Milk. Adv. Nutr. 2012, 3 (3), 473S–482S. 10.3945/an.111.001412. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Troyano E.; Olano A.; Martínez-Castro I. Presence of Free N -Acetylgalactosamine in Raw Milk. J. Dairy Res. 1995, 62 (1), 147–150. 10.1017/S0022029900033768. [DOI] [Google Scholar]
- McGuire M. K.; Meehan C. L.; McGuire M. A.; Williams J. E.; Foster J.; Sellen D. W.; Kamau-Mbuthia E. W.; Kamundia E. W.; Mbugua S.; Moore S. E.; Prentice A. M.; Kvist L. J.; Otoo G. E.; Brooker S. L.; Price W. J.; Shafii B.; Placek C.; Lackey K. A.; Robertson B.; Manzano S.; Ruíz L.; Rodríguez J. M.; Pareja R. G.; Bode L. What’s Normal? Oligosaccharide Concentrations and Profiles in Milk Produced by Healthy Women Vary Geographically. Am. J. Clin. Nutr. 2017, 105 (5), 1086–1100. 10.3945/ajcn.116.139980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Quinn E.; Slattery H.; Thompson A.; Kilcoyne M.; Joshi L.; Hickey R. Mining Milk for Factors Which Increase the Adherence of Bifidobacterium Longum Subsp. Infantis to Intestinal Cells. Foods 2018, 7 (12), 196. 10.3390/foods7120196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Leeuwen S. S.; Stoutjesdijk E.; ten Kate G. A.; Schaafsma A.; Dijck-Brouwer J.; Muskiet F. A. J.; Dijkhuizen L. Regional Variations in Human Milk Oligosaccharides in Vietnam Suggest FucTx Activity besides FucT2 and FucT3. Sci. Rep. 2018, 8 (1), 16790. 10.1038/s41598-018-34882-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Austin S.; De Castro C. A.; Sprenger N.; Binia A.; Affolter M.; Garcia-Rodenas C. L.; Beauport L.; Tolsa J.-F.; Fumeaux C. J. F. Human Milk Oligosaccharides in the Milk of Mothers Delivering Term versus Preterm Infants. Nutrients 2019, 11 (6), 1282. 10.3390/nu11061282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lu J.; Zhang Y.; Song B.; Zhang S.; Pang X.; Sari R. N.; Liu L.; Wang J.; Lv J. Comparative Analysis of Oligosaccharides in Guanzhong and Saanen Goat Milk by Using LC–MS/MS. Carbohydr. Polym. 2020, 235, 115965. 10.1016/j.carbpol.2020.115965. [DOI] [PubMed] [Google Scholar]
- Neveu C.; Riaublanc A.; Miranda G.; Chich J.-F.; Martin P. Is the Apocrine Milk Secretion Process Observed in the Goat Species Rooted in the Perturbation of the Intracellular Transport Mechanism Induced by Defective Alleles at the? S1 -Cn Locus?. Reprod., Nutr., Dev. 2002, 42 (2), 163–172. 10.1051/rnd:2002015. [DOI] [PubMed] [Google Scholar]
- Lu Y.; Jin W.; Yang Y.; Jia Y.; Sun L.; Liu J.; Wang L.; Zhang F.; Ge W.; Wang J.; Huang L.; Wang Z. Online LC-UV-ESI-MS/MS Comparative Analysis of Changes in Goat Colostrum N/O -Glycopatterns at Different Parities. J. Agric. Food Chem. 2020, 68 (7), 2174–2182. 10.1021/acs.jafc.9b07075. [DOI] [PubMed] [Google Scholar]
- Nakamura T.; Kawase H.; Kimura K.; Watanabe Y.; Ohtani M.; Arai I.; Urashima T. Concentrations of Sialyloligosaccharides in Bovine Colostrum and Milk during the Prepartum and Early Lactation. J. Dairy Sci. 2003, 86 (4), 1315–1320. 10.3168/jds.S0022-0302(03)73715-1. [DOI] [PubMed] [Google Scholar]
- Mehra R.; Barile D.; Marotta M.; Lebrilla C. B.; Chu C.; German J. B. Novel High-Molecular Weight Fucosylated Milk Oligosaccharides Identified in Dairy Streams. PLoS One 2014, 9 (5), e96040 10.1371/journal.pone.0096040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goehring K. C.; Marriage B. J.; Oliver J. S.; Wilder J. A.; Barrett E. G.; Buck R. H. Similar to Those Who Are Breastfed, Infants Fed a Formula Containing 2′-Fucosyllactose Have Lower Inflammatory Cytokines in a Randomized Controlled Trial. J. Nutr. 2016, 146 (12), 2559–2566. 10.3945/jn.116.236919. [DOI] [PubMed] [Google Scholar]
- Marriage B. J.; Buck R. H.; Goehring K. C.; Oliver J. S.; Williams J. A. Infants Fed a Lower Calorie Formula With 2′FL Show Growth and 2′FL Uptake Like Breast-Fed Infants. J. Pediatr. Gastroenterol. Nutr. 2015, 61 (6), 649–658. 10.1097/MPG.0000000000000889. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thum C.; Roy N. C.; McNabb W. C.; Otter D. E.; Cookson A. L. In Vitro Fermentation of Caprine Milk Oligosaccharides by Bifidobacteria Isolated from Breast-Fed Infants. Gut Microbes 2015, 6 (6), 352–363. 10.1080/19490976.2015.1105425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thum C.; Weinborn V.; Barile D.; C McNabb W.; C Roy N.; Maria Leite Nobrega de Moura Bell J. Understanding the Effects of Lactose Hydrolysis Modeling on the Main Oligosaccharides in Goat Milk Whey Permeate. Molecules 2019, 24 (18), 3294. 10.3390/molecules24183294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gibson G. R.; Wang X. Regulatory Effects of Bifidobacteria on the Growth of Other Colonic Bacteria. J. Appl. Bacteriol. 1994, 77 (4), 412–420. 10.1111/j.1365-2672.1994.tb03443.x. [DOI] [PubMed] [Google Scholar]
- Koh A.; De Vadder F.; Kovatcheva-Datchary P.; Bäckhed F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165 (6), 1332–1345. 10.1016/j.cell.2016.05.041. [DOI] [PubMed] [Google Scholar]
- Sanna S.; van Zuydam N. R.; Mahajan A.; Kurilshikov A.; Vich Vila A.; Võsa U.; Mujagic Z.; Masclee A. A. M.; Jonkers D. M. A. E.; Oosting M.; Joosten L. A. B.; Netea M. G.; Franke L.; Zhernakova A.; Fu J.; Wijmenga C.; McCarthy M. I. Causal Relationships among the Gut Microbiome, Short-Chain Fatty Acids and Metabolic Diseases. Nat. Genet. 2019, 51 (4), 600–605. 10.1038/s41588-019-0350-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ratajczak W.; Rył A.; Mizerski A.; Walczakiewicz K.; Sipak O.; Laszczyńska M. Immunomodulatory Potential of Gut Microbiome-Derived Short-Chain Fatty Acids (SCFAs). Acta Biochim. Polym. 2019, 66 (1), 1–12. 10.18388/abp.2018_2648. [DOI] [PubMed] [Google Scholar]
- Chambers E. S.; Preston T.; Frost G.; Morrison D. J. Role of Gut Microbiota-Generated Short-Chain Fatty Acids in Metabolic and Cardiovascular Health. Curr. Nutr. Rep. 2018, 7 (4), 198–206. 10.1007/s13668-018-0248-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen J.; Li Y.; Tian Y.; Huang C.; Li D.; Zhong Q.; Ma X. Interaction between Microbes and Host Intestinal Health: Modulation by Dietary Nutrients and Gut-Brain-Endocrine-Immune Axis. Curr. Protein Pept. Sci. 2015, 16 (7), 592–603. 10.2174/1389203716666150630135720. [DOI] [PubMed] [Google Scholar]
- Quinn E. M.; Slattery H.; Walsh D.; Joshi L.; Hickey R. M. Bifidobacterium Longum Subsp. Infantis ATCC 15697 and Goat Milk Oligosaccharides Show Synergism In Vitro as Anti-Infectives against Campylobacter Jejuni. Foods 2020, 9 (3), 348. 10.3390/foods9030348. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leong A.; Liu Z.; Almshawit H.; Zisu B.; Pillidge C.; Rochfort S.; Gill H. Oligosaccharides in Goats’ Milk-Based Infant Formula and Their Prebiotic and Anti-Infection Properties. Br. J. Nutr. 2019, 122 (04), 441–449. 10.1017/S000711451900134X. [DOI] [PubMed] [Google Scholar]
- Thum C.; McNabb W. C.; Young W.; Cookson A. L.; Roy N. C. Prenatal Caprine Milk Oligosaccharide Consumption Affects the Development of Mice Offspring. Mol. Nutr. Food Res. 2016, 60 (9), 2076–2085. 10.1002/mnfr.201600118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moens F.; Verce M.; De Vuyst L. Lactate- and Acetate-Based Cross-Feeding Interactions between Selected Strains of Lactobacilli, Bifidobacteria and Colon Bacteria in the Presence of Inulin-Type Fructans. Int. J. Food Microbiol. 2017, 241, 225–236. 10.1016/j.ijfoodmicro.2016.10.019. [DOI] [PubMed] [Google Scholar]
- Smilowitz J. T.; Lemay D. G.; Kalanetra K. M.; Chin E. L.; Zivkovic A. M.; Breck M. A.; German J. B.; Mills D. A.; Slupsky C.; Barile D. Tolerability and Safety of the Intake of Bovine Milk Oligosaccharides Extracted from Cheese Whey in Healthy Human Adults. J. Nutr. Sci. 2017, 6, e6 10.1017/jns.2017.2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Westreich S. T.; Salcedo J.; Durbin-Johnson B.; Smilowitz J. T.; Korf I.; Mills D. A.; Barile D.; Lemay D. G. Fecal Metatranscriptomics and Glycomics Suggest That Bovine Milk Oligosaccharides Are Fully Utilized by Healthy Adults. J. Nutr. Biochem. 2020, 79, 108340. 10.1016/j.jnutbio.2020.108340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee H.; Cuthbertson D. J.; Otter D. E.; Barile D. Rapid Screening of Bovine Milk Oligosaccharides in a Whey Permeate Product and Domestic Animal Milks by Accurate Mass Database and Tandem Mass Spectral Library. J. Agric. Food Chem. 2016, 64 (32), 6364–6374. 10.1021/acs.jafc.6b02039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vandenplas Y.; Berger B.; Carnielli V. P.; Ksiazyk J.; Lagström H.; Sanchez Luna M.; Migacheva N.; Mosselmans J. M.; Picaud J. C.; Possner M.; Singhal A.; Wabitsch M. Human Milk Oligosaccharides: 2′-Fucosyllactose (2′-FL) and Lacto-N-Neotetraose (LNnT) in Infant Formula. Nutrients 2018, 10 (9), 1161. 10.3390/nu10091161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berger B.; Porta N.; Foata F.; Grathwohl D.; Delley M.; Moine D.; Charpagne A.; Siegwald L.; Descombes P.; Alliet P.; Puccio G.; Steenhout P.; Mercenier A.; Sprenger N. Linking Human Milk Oligosaccharides, Infant Fecal Community Types, and Later Risk To Require Antibiotics. mBio 2020, 11 (2), e03196-19. 10.1128/mBio.03196-19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Daddaoua A.; Puerta V.; Requena P.; Martínez-Férez A.; Guadix E.; Sánchez de Medina F.; Zarzuelo A.; Suárez M. D.; Boza J. J.; Martínez-Augustin O. Goat Milk Oligosaccharides Are Anti-Inflammatory in Rats with Hapten-Induced Colitis. J. Nutr. 2006, 136 (3), 672–676. 10.1093/jn/136.3.672. [DOI] [PubMed] [Google Scholar]
- Martinez-Ferez A.; Guadix A.; Guadix E. M. Recovery of Caprine Milk Oligosaccharides with Ceramic Membranes. J. Membr. Sci. 2006, 276 (1–2), 23–30. 10.1016/j.memsci.2005.09.027. [DOI] [Google Scholar]
- Lara-Villoslada F.; Debras E.; Nieto A.; Concha A.; Gálvez J.; López-Huertas E.; Boza J.; Obled C.; Xaus J. Oligosaccharides Isolated from Goat Milk Reduce Intestinal Inflammation in a Rat Model of Dextran Sodium Sulfate-Induced Colitis. Clin. Nutr. 2006, 25 (3), 477–488. 10.1016/j.clnu.2005.11.004. [DOI] [PubMed] [Google Scholar]
- Holma R.; Juvonen P.; Asmawi M. Z.; Vapaatalo H.; Korpela R. Galacto-Oligosaccharides Stimulate the Growth of Bifidobacteria but Fail to Attenuate Inflammation in Experimental Colitis in Rats. Scand. J. Gastroenterol. 2002, 37 (9), 1042–1047. 10.1080/003655202320378239. [DOI] [PubMed] [Google Scholar]
- Triantis V.; Bode L.; van Neerven R. J. J. Immunological Effects of Human Milk Oligosaccharides. Front. Pediatr. 2018, 6, 6. 10.3389/fped.2018.00190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hirabayashi J. Oligosaccharide Specificity of Galectins: A Search by Frontal Affinity Chromatography. Biochim. Biophys. Acta, Gen. Subj. 2002, 1572 (2–3), 232–254. 10.1016/S0304-4165(02)00311-2. [DOI] [PubMed] [Google Scholar]
- Brinchmann M. F.; Patel D. M.; Iversen M. H. The Role of Galectins as Modulators of Metabolism and Inflammation. Mediators Inflammation 2018, 2018, 1–11. 10.1155/2018/9186940. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Salomonsson E.; Carlsson M. C.; Osla V.; Hendus-Altenburger R.; Kahl-Knutson B.; Öberg C. T.; Sundin A.; Nilsson R.; Nordberg-Karlsson E.; Nilsson U. J.; Karlsson A.; Rini J. M.; Leffler H. Mutational Tuning of Galectin-3 Specificity and Biological Function. J. Biol. Chem. 2010, 285 (45), 35079–35091. 10.1074/jbc.M109.098160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lopez-Andrès N.; Rossignol P.; Iraqi W.; Fay R.; Nuée J.; Ghio S.; Cleland J. G. F.; Zannad F.; Lacolley P. Association of Galectin-3 and Fibrosis Markers with Long-Term Cardiovascular Outcomes in Patients with Heart Failure, Left Ventricular Dysfunction, and Dyssynchrony: Insights from the CARE-HF (Cardiac Resynchronization in Heart Failure) Trial. Eur. J. Heart Failure 2012, 14 (1), 74–81. 10.1093/eurjhf/hfr151. [DOI] [PubMed] [Google Scholar]
- de Kivit S.; Kraneveld A. D.; Knippels L. M. J.; van Kooyk Y.; Garssen J.; Willemsen L. E. M. Intestinal Epithelium-Derived Galectin-9 Is Involved in the Immunomodulating Effects of Nondigestible Oligosaccharides. J. Innate Immun. 2013, 5 (6), 625–638. 10.1159/000350515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kivit S.; Saeland E.; Kraneveld A. D.; Kant H. J. G.; Schouten B.; Esch B. C. A. M.; Knol J.; Sprikkelman A. B.; Aa L. B.; Knippels L. M. J.; Garssen J.; Kooyk Y.; Willemsen L. E. M. Galectin-9 Induced by Dietary Synbiotics Is Involved in Suppression of Allergic Symptoms in Mice and Humans. Allergy 2012, 67 (3), 343–352. 10.1111/j.1398-9995.2011.02771.x. [DOI] [PubMed] [Google Scholar]
- Robinson B. S.; Arthur C. M.; Evavold B.; Roback E.; Kamili N. A.; Stowell C. S.; Vallecillo-Zúniga M. L.; Van Ry P. M.; Dias-Baruffi M.; Cummings R. D.; Stowell S. R. The Sweet-Side of Leukocytes: Galectins as Master Regulators of Neutrophil Function. Front. Immunol. 2019, 10, 10. 10.3389/fimmu.2019.01762. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Figueroa-Lozano S.; Ren C.; Yin H.; Pham H.; van Leeuwen S.; Dijkhuizen L.; de Vos P. The Impact of Oligosaccharide Content, Glycosidic Linkages and Lactose Content of Galacto-Oligosaccharides (GOS) on the Expression of Mucus-Related Genes in Goblet Cells. Food Funct. 2020, 11 (4), 3506–3515. 10.1039/D0FO00064G. [DOI] [PubMed] [Google Scholar]
- Cheng L.; Kiewiet M. B. G.; Groeneveld A.; Nauta A.; de Vos P. Human Milk Oligosaccharides and Its Acid Hydrolysate LNT2 Show Immunomodulatory Effects via TLRs in a Dose and Structure-Dependent Way. J. Funct. Foods 2019, 59 (May), 174–184. 10.1016/j.jff.2019.05.023. [DOI] [Google Scholar]
- Ortega-González M.; Ocón B.; Romero-Calvo I.; Anzola A.; Guadix E.; Zarzuelo A.; Suárez M. D.; Sánchez de Medina F.; Martínez-Augustin O. Nondigestible Oligosaccharides Exert Nonprebiotic Effects on Intestinal Epithelial Cells Enhancing the Immune Response via Activation of TLR4-NFκB. Mol. Nutr. Food Res. 2014, 58 (2), 384–393. 10.1002/mnfr.201300296. [DOI] [PubMed] [Google Scholar]
- Kurakevich E.; Hennet T.; Hausmann M.; Rogler G.; Borsig L. Milk Oligosaccharide Sialyl(2,3)Lactose Activates Intestinal CD11c+ Cells through TLR4. Proc. Natl. Acad. Sci. U. S. A. 2013, 110 (43), 17444–17449. 10.1073/pnas.1306322110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perdijk O.; van Neerven R. J. J.; Meijer B.; Savelkoul H. F. J.; Brugman S. Induction of Human Tolerogenic Dendritic Cells by 3′-Sialyllactose via TLR4 Is Explained by LPS Contamination. Glycobiology 2018, 28 (3), 126–130. 10.1093/glycob/cwx106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barnett A. M.; Roy N. C.; McNabb W. C.; Cookson A. L. Effect of a Semi-Purified Oligosaccharide-Enriched Fraction from Caprine Milk on Barrier Integrity and Mucin Production of Co-Culture Models of the Small and Large Intestinal Epithelium. Nutrients 2016, 8 (5), 267. 10.3390/nu8050267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheng L.; Kong C.; Walvoort M. T. C.; Faas M. M.; Vos P. Human Milk Oligosaccharides Differently Modulate Goblet Cells Under Homeostatic, Proinflammatory Conditions and Under ER-Stress. Mol. Nutr. Food Res. 2020, 64, 1900976. 10.1002/mnfr.201900976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu R. Y.; Li B.; Koike Y.; Määttänen P.; Miyake H.; Cadete M.; Johnson-Henry K. C.; Botts S. R.; Lee C.; Abrahamsson T. R.; Landberg E.; Pierro A.; Sherman P. M. Human Milk Oligosaccharides Increase Mucin Expression in Experimental Necrotizing Enterocolitis. Mol. Nutr. Food Res. 2018, 63 (3), 1800658. 10.1002/mnfr.201800658. [DOI] [PubMed] [Google Scholar]
- Lis-Kuberka J.; Orczyk-Pawiłowicz M. Sialylated Oligosaccharides and Glycoconjugates of Human Milk. The Impact on Infant and Newborn Protection, Development and Well-Being. Nutrients 2019, 11 (2), 306. 10.3390/nu11020306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gringhuis S. I.; den Dunnen J.; Litjens M.; van der Vlist M.; Geijtenbeek T. B. H. Carbohydrate-Specific Signaling through the DC-SIGN Signalosome Tailors Immunity to Mycobacterium Tuberculosis, HIV-1 and Helicobacter Pylori. Nat. Immunol. 2009, 10 (10), 1081–1088. 10.1038/ni.1778. [DOI] [PubMed] [Google Scholar]
- van Liempt E.; Bank C. M. C.; Mehta P.; García-Vallejo J. J.; Kawar Z. S.; Geyer R.; Alvarez R. A.; Cummings R. D.; van Kooyk Y.; van Die I. Specificity of DC-SIGN for Mannose- and Fucose-Containing Glycans. FEBS Lett. 2006, 580 (26), 6123–6131. 10.1016/j.febslet.2006.10.009. [DOI] [PubMed] [Google Scholar]
- Hong P.; Ninonuevo M. R.; Lee B.; Lebrilla C.; Bode L. Human Milk Oligosaccharides Reduce HIV-1-Gp120 Binding to Dendritic Cell-Specific ICAM3-Grabbing Non-Integrin (DC-SIGN). Br. J. Nutr. 2009, 101 (4), 482–486. 10.1017/S0007114508025804. [DOI] [PubMed] [Google Scholar]
- Kuhn L.; Kim H.-Y.; Hsiao L.; Nissan C.; Kankasa C.; Mwiya M.; Thea D. M.; Aldrovandi G. M.; Bode L. Oligosaccharide Composition of Breast Milk Influences Survival of Uninfected Children Born to HIV-Infected Mothers in Lusaka, Zambia. J. Nutr. 2015, 145 (1), 66–72. 10.3945/jn.114.199794. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saeland E.; de Jong M. A. W. P.; Nabatov A. A.; Kalay H.; Geijtenbeek T. B. H.; van Kooyk Y. MUC1 in Human Milk Blocks Transmission of Human Immunodeficiency Virus from Dendritic Cells to T Cells. Mol. Immunol. 2009, 46 (11–12), 2309–2316. 10.1016/j.molimm.2009.03.025. [DOI] [PubMed] [Google Scholar]
- Angeloni S. Glycoprofiling with Micro-Arrays of Glycoconjugates and Lectins. Glycobiology 2004, 15 (1), 31–41. 10.1093/glycob/cwh143. [DOI] [PubMed] [Google Scholar]
- Newburg D. S. Glycobiology of Human Milk. Biochemistry 2013, 78 (7), 771–785. 10.1134/S0006297913070092. [DOI] [PubMed] [Google Scholar]
- Wang Z.; Chinoy Z. S.; Ambre S. G.; Peng W.; McBride R.; de Vries R. P.; Glushka J.; Paulson J. C.; Boons G.-J. A General Strategy for the Chemoenzymatic Synthesis of Asymmetrically Branched N-Glycans. Science (Washington, DC, U. S.) 2013, 341 (6144), 379–383. 10.1126/science.1236231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prudden A. R.; Liu L.; Capicciotti C. J.; Wolfert M. A.; Wang S.; Gao Z.; Meng L.; Moremen K. W.; Boons G.-J. Synthesis of Asymmetrical Multiantennary Human Milk Oligosaccharides. Proc. Natl. Acad. Sci. U. S. A. 2017, 114 (27), 6954–6959. 10.1073/pnas.1701785114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lu W.; Du W.; Somovilla V. J.; Yu G.; Haksar D.; de Vries E.; Boons G.-J.; de Vries R. P.; de Haan C. A. M.; Pieters R. J. Enhanced Inhibition of Influenza A Virus Adhesion by Di- and Trivalent Hemagglutinin Inhibitors. J. Med. Chem. 2019, 62 (13), 6398–6404. 10.1021/acs.jmedchem.9b00303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klamer Z.; Staal B.; Prudden A. R.; Liu L.; Smith D. F.; Boons G.; Haab B. Mining High-Complexity Motifs in Glycans: A New Language To Uncover the Fine Speci Fi Cities of Lectins and Glycosidases. Anal. Chem. 2017, 89 (22), 12342–12350. 10.1021/acs.analchem.7b04293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Autran C. A.; Schoterman M. H. C.; Jantscher-Krenn E.; Kamerling J. P.; Bode L. Sialylated Galacto-Oligosaccharides and 2′-Fucosyllactose Reduce Necrotising Enterocolitis in Neonatal Rats. Br. J. Nutr. 2016, 116 (2), 294–299. 10.1017/S0007114516002038. [DOI] [PubMed] [Google Scholar]
- Sumiyoshi W.; Urashima T.; Nakamura T.; Arai I.; Nagasawa T.; Saito T.; Tsumura N.; Wang B.; Brand-Miller J.; Watanabe Y.; Kimura K. Galactosyllactoses in the Milk of Japanese Women: Changes in Concentration during the Course of Lactation. J. Appl. Glycosci. 2004, 51 (4), 341–344. 10.5458/jag.51.341. [DOI] [Google Scholar]
- Huang X.; Zhu B.; Jiang T.; Yang C.; Qiao W.; Hou J.; Han Y.; Xiao H.; Chen L. Improved Simple Sample Pretreatment Method for Quantitation of Major Human Milk Oligosaccharides Using Ultrahigh Pressure Liquid Chromatography with Fluorescence Detection. J. Agric. Food Chem. 2019, 67 (44), 12237–12244. 10.1021/acs.jafc.9b03445. [DOI] [PubMed] [Google Scholar]
- Pham H. T. T.; ten Kate G. A.; Dijkhuizen L.; van Leeuwen S. S. Synthesis and Characterization of Sialylated Lactose and Lactulose Derived Oligosaccharides by Trypanosoma cruzi Trans-sialidase. J. Agric. Food Chem. 2019, 67 (12), 3469–3479. 10.1021/acs.jafc.8b06974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Urakami H.; Saeki M.; Watanabe Y.; Kawamura R.; Nishizawa S.; Suzuki Y.; Watanabe A.; Ajisaka K. Isolation and Assessment of Acidic and Neutral Oligosaccharides from Goat Milk and Bovine Colostrum for Use as Ingredients of Infant Formulae. Int. Dairy J. 2018, 83, 1–9. 10.1016/j.idairyj.2018.03.004. [DOI] [Google Scholar]
- Ruiz-Palacios G. M.; Cervantes L. E.; Ramos P.; Chavez-Munguia B.; Newburg D. S. Campylobacter Jejuni Binds Intestinal H(O) Antigen (Fucα1, 2Galβ1, 4GlcNAc), and Fucosyloligosaccharides of Human Milk Inhibit Its Binding and Infection. J. Biol. Chem. 2003, 278 (16), 14112–14120. 10.1074/jbc.M207744200. [DOI] [PubMed] [Google Scholar]
- Brassart D.; Woltz A.; Golliard M.; Neeser J. R. In Vitro Inhibition of Adhesion of Candida Albicans Clinical Isolates to Human Buccal Epithelial Cells by Fuc Alpha 1----2Gal Beta-Bearing Complex Carbohydrates. Infect. Immun. 1991, 59 (5), 1605–1613. 10.1128/IAI.59.5.1605-1613.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morrow A. L.; Ruiz-Palacios G. M.; Jiang X.; Newburg D. S. Human-Milk Glycans That Inhibit Pathogen Binding Protect Breast-Feeding Infants against Infectious Diarrhea. J. Nutr. 2005, 135 (5), 1304–1307. 10.1093/jn/135.5.1304. [DOI] [PubMed] [Google Scholar]
- Coppa G. V.; Zampini L.; Galeazzi T.; Facinelli B.; Ferrante L.; Capretti R.; Orazio G. Human Milk Oligosaccharides Inhibit the Adhesion to Caco-2 Cells of Diarrheal Pathogens: Escherichia Coli, Vibrio Cholerae, and Salmonella Fyris. Pediatr. Res. 2006, 59 (3), 377–382. 10.1203/01.pdr.0000200805.45593.17. [DOI] [PubMed] [Google Scholar]
- Jantscher-Krenn E.; Lauwaet T.; Bliss L. a.; Reed S. L.; Gillin F. D.; Bode L. Human Milk Oligosaccharides Reduce Entamoeba Histolytica Attachment and Cytotoxicity in Vitro. Br. J. Nutr. 2012, 108 (10), 1839–1846. 10.1017/S0007114511007392. [DOI] [PubMed] [Google Scholar]
- Kunz C.; Rudloff S.; Baier W.; Klein N.; Strobel S. Oligosaccharides in Human Milk: Structural, Functional, and Metabolic Aspects. Annu. Rev. Nutr. 2000, 20 (1), 699–722. 10.1146/annurev.nutr.20.1.699. [DOI] [PubMed] [Google Scholar]
- Lin A. E.; Autran C. A.; Szyszka A.; Escajadillo T.; Huang M.; Godula K.; Prudden A. R.; Boons G.-J.; Lewis A. L.; Doran K. S.; Nizet V.; Bode L. Human Milk Oligosaccharides Inhibit Growth of Group B Streptococcus. J. Biol. Chem. 2017, 292 (27), 11243–11249. 10.1074/jbc.M117.789974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ackerman D. L.; Craft K. M.; Doster R. S.; Weitkamp J.-H.; Aronoff D. M.; Gaddy J. A.; Townsend S. D. Antimicrobial and Antibiofilm Activity of Human Milk Oligosaccharides against Streptococcus Agalactiae, Staphylococcus Aureus, and Acinetobacter Baumannii. ACS Infect. Dis. 2018, 4 (3), 315–324. 10.1021/acsinfecdis.7b00183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Andersson B.; Porras O.; Hanson L. A.; Lagergard T.; Svanborg-Eden C. Inhibition of Attachment of Streptococcus Pneumoniae and Haemophilus Influenzae by Human Milk and Receptor Oligosaccharides. J. Infect. Dis. 1986, 153 (2), 232–237. 10.1093/infdis/153.2.232. [DOI] [PubMed] [Google Scholar]
- Andersson B; Dahmén J; Frejd T; Leffler H; Magnusson G; Noori G; Edén C S Identification of an Active Disaccharide Unit of a Glycoconjugate Receptor for Pneumococci Attaching to Human Pharyngeal Epithelial Cells. J. Exp. Med. 1983, 158 (2), 559–570. 10.1084/jem.158.2.559. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mysore J. V.; Wigginton T.; Simon P. M.; Zopf D.; Heman-Ackah L. M.; Dubois A. Treatment of Helicobacter Pylori Infection in Rhesus Monkeys Using a Novel Antiadhesion Compound. Gastroenterology 1999, 117 (6), 1316–1325. 10.1016/S0016-5085(99)70282-9. [DOI] [PubMed] [Google Scholar]
- Thomas R.; Brooks T. Common Oligosaccharide Moieties Inhibit the Adherence of Typical and Atypical Respiratory Pathogens. J. Med. Microbiol. 2004, 53 (9), 833–840. 10.1099/jmm.0.45643-0. [DOI] [PubMed] [Google Scholar]