

Abstract
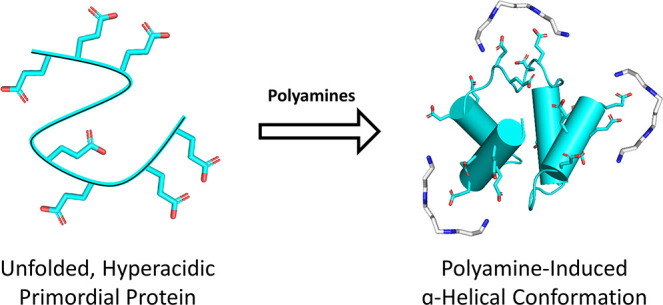
Polyamines are known to mediate diverse biological processes, and specifically to bind and stabilize compact conformations of nucleic acids, acting as chemical chaperones that promote folding by offsetting the repulsive negative charges of the phosphodiester backbone. However, whether and how polyamines modulate the structure and function of proteins remain unclear. In particular, early proteins are thought to have been highly acidic, like nucleic acids, due to a scarcity of basic amino acids in the prebiotic context. Perhaps polyamines, the abiotic synthesis of which is simple, could have served as chemical chaperones for such primordial proteins? We replaced all lysines of an ancestral 60-residue helix-bundle protein with glutamate, resulting in a disordered protein with 21 glutamates in total. Polyamines efficiently induce folding of this hyperacidic protein at submillimolar concentrations, and their potency scaled with the number of amine groups. Compared to cations, polyamines were several orders of magnitude more potent than Na+, while Mg2+ and Ca2+ had an effect similar to that of a diamine, inducing folding at approximately seawater concentrations. We propose that (i) polyamines and dications may have had a role in promoting folding of early proteins devoid of basic residues and (ii) coil–helix transitions could be the basis of polyamine regulation in contemporary proteins.
Natural polyamines contain two or more amino groups separated by aliphatic hydrocarbon chains and can be either linear or branched. At neutral pH, polyamines are polycations that—unlike metal ions, which are effectively point charges—encode patterns of regularly spaced positively charged sites. Low-molecular weight, linear polyamines (see Figure 1 for the most common natural examples) are present in all living organisms1 and essential for cell survival.2 The functional profile of these and other polyamines is diverse, and these simple metabolites mediate a variety of biological processes, including chromatin structure remodeling,3 gene transcription and translation,4,5 cell proliferation,6 and circadian clock regulation.7 Although the intracellular concentrations of polyamines are not well characterized, current evidence suggests it can be quite high. The concentrations of spermidine and putrescine in Escherichia coli, for example, are estimated to be ∼6 and ∼20 mM, respectively,8 though the fraction of bound versus free polyamine is unknown. Intracellular polyamine concentrations have also been shown to vary significantly with the cellular state, thus suggesting a regulatory role for polyamines.9 Mechanistically, polyamines operate on nucleic acids through interactions with the negatively charged phosphodiester backbone, resulting in changes in DNA conformation,10,11 tRNA stabilization,12 and even phase separation.13 In the laboratory, polyamines have been employed to promote the folding and stability of nucleic acids in vitro.14−16 However, with respect to proteins, modulation of structure and/or function by polyamines has centered largely on amyloidogenesis;17−19 consequently, the mechanisms by which protein structure and function can be modulated by polyamines are largely unknown.
Figure 1.

Natural polyamines used in this study. Polyamines are polycations at neutral pH, with pKas that range from 8 to 11.51,52
A second interesting aspect of polyamines is their potential role at the dawn of life. Given the ubiquity, essentiality, and relatively simple chemical structure of polyamines, it is likely that they are ancient metabolites that emerged early in life’s history, perhaps as chemical chaperones for early nucleic acids, including ancient ribozymes.20 Consistent with this view, primordial synthesis of polyamines has been observed.21 Here, we ask whether polyamines might have had a role in the emergence of the early proteins. Modern proteins are comprised of both positively and negatively charged amino acids, and the charge of most proteins is either moderately negative or moderately positive to maximize foldability and solubility around neutral pH. While solubility may be enhanced by significant negative charge, foldability will be hindered. In the absence of positively charged residues, salt bridges—which are critical for protein stability, as reflected in thermophilic proteins22—are lacking, leaving only the repulsive interactions between adjacent negative charges that destabilize compact structures. This may not be an issue in stable, modern proteins; their surface can be rendered highly acidic by either chemical modification23 or site-directed mutagenesis.24 However, repulsive interactions would become a severely limiting factor in the absence of a large well-packed hydrophobic core, as expected at the early stages of protein evolution. In early life contexts, the availability of the contemporary basic amino acids was likely poor.25,26 Alternative basic amino acids such ornithine could have been a substitute,27,28 but the possibility that basic amino acids were depleted in, or even entirely absent from, the first proteins cannot be ruled out.29−31 Thus, understanding whether and how highly acidic proteins could fold or function remain major unsolved problems in early protein evolution. Furthermore, folding of acidic proteins is fundamentally the same problem faced by the ribozyme, which employs chemical chaperones, most frequently dications, to enable globular packing. Perhaps the earliest proteins were similarly dependent on chemical chaperones?
To determine if polyamines can operate as efficient chemical chaperones for acidic proteins, we constructed a hyperacidic version of a previously described reconstructed ancestral protein comprising a tandem duplication of a helix–hairpin–helix (HhH) motif (Figure 2).27,32 This 60-residue protein, dubbed acidic-(HhH)2, is devoid of arginine and lysine and has a total of 21 glutamate residues. As expected, acidic-(HhH)2 is unfolded in low-salt buffer and required near-molar concentrations of NaCl to achieve complete folding. Addition of submillimolar concentrations of polyamines, however, induced formation of α-helical structure. Our results inspire the hypothesis that polyamines served as chemical chaperones of ancient proteins, and that this feature may exist in polyamine-regulated proteins today.
Figure 2.
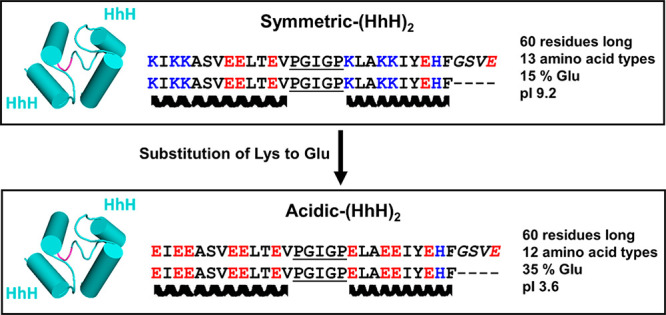
Design of acidic-(HhH)2. Symmetric-(HhH)2 is a fully symmetric protein constructed in a previous study to understand the properties of ancient protein forms.27 Derived from a family of dsDNA binding proteins, it is positively charged at neutral pH. To generate a hyperacidic model protein for this study, all lysine residues in symmetric-(HhH)2 were substituted with glutamate (see Results for more details). The conserved loop residues between the two helices of the HhH motif (PGIGP) are underlined, and the linker residues (GSVE) between the two HhH motifs are rendered in italics in the sequence and colored magenta in the structural model. The C-terminus of acidic-(HhH)2 bears a 6xHis tag connected by a two-residue Leu-Glu linker (not shown).
Materials and Methods
Protein Expression and Purification
Synthetic genes were ordered from Twist Bioscience (www.twistbioscience.com) and cloned into a pET21a vector, yielding variants with a C-terminal 6x-His tag. All constructs were verified by Sanger sequencing. Transformed E. coli BL21(DE3) cells were induced at an OD600 of ∼0.6 with 1 mM isopropyl β-d-1-thiogalactopyranoside. Induced cells were shake-incubated at 20 °C overnight. Cell pellets were collected by centrifugation and frozen at −20 °C for storage. Cell pellets from 250 mL cultures were resuspended in 30 mL of 100 mM NaCl, 50 mM Tris-HCl (pH 7.5), 2.5 units/mL Benzonase (Merck Millipore), 0.2 unit/mL rDNase1 (Thermo Fisher Scientific), 0.25× Protease Inhibitor Cocktail for histidine-tagged proteins (Sigma-Aldrich), and 0.3 mg/mL lysozyme and shake-incubated at 37 °C for 90 min. The lysates were then cooled on ice for 15 min and sonicated for 5 min, with a 30 s cooling period after every 30 s of sonication. After cell lysis, the samples were spiked with 0.5 M NaCl and 5 mM imidazole and gently rocked for 30 min at room temperature (moderate concentrations of NaCl decreased the level of unspecific binding to Ni-NTA and also promoted solubility of the parent protein27). Pellets were clarified by centrifugation and passed through a 0.45 μm sterile filter. Clarified lysates were applied to 3 mL of packed Ni-NTA resin pre-equilibrated in 5 mM imidazole, 0.5 M NaCl, and 50 mM Tris-HCl (pH 8.0) (“purification buffer”). After sample application, the resin was washed with 25 mL of purification buffer. Nonspecifically bound proteins were removed by washing with purification buffer spiked with 25 mM imidazole. The bound protein was eluted with 20 mL of 500 mM imidazole, 50 mM Tris-HCl (pH 8.0), and 500 mM NaCl. In samples used for nuclear magnetic resonance (NMR) analysis, an additional wash step of 20 mL of 6 M GuHCl was performed. The eluted protein fractions were dialyzed against 20 mM bis-Tris (pH 5.5) and 50 mM NaCl using a 3.5 kDa molecular weight cutoff (MWCO) SnakeSkin Dialysis Tube (Thermo Fisher Scientific, Waltham, MA). Next, the dialyzed sample was loaded on an anion exchange HiTrap Q HP 5 mL column (GE healthcare life sciences, Boston, MA) pre-equilibrated with 20 mM bis-Tris (pH 5.5) and 50 mM NaCl at a flow rate of 1 mL/min using an FPLC AKTA-prime plus chromatography system (GE). The protein was eluted by a linear gradient of NaCl [0.05–1 M NaCl in 20 mM bis-Tris (pH 5.5)] over the course of 20 column volumes at a flow rate of 2 mL/min. The fractions containing acidic-(HhH)2 were combined and dialyzed against 5 mM Tris (pH 7.5) and 25 mM NaCl. Finally, the sample was concentrated using a 3 kDa MWCO centrifugal filtration unit (Thermo Fisher Scientific), and the protein concentration was measured with a BCA kit (Thermo Fisher Scientific).
Circular Dichroism (CD) Spectroscopy
CD spectra were recorded on a Chirascan circular dichroism spectrometer (Applied Photophysics). Samples containing 5 μM protein in 5 mM Tris-HCl (pH 7.5) and 25 mM NaCl (unless otherwise stated) were placed in a 1 mm path length quartz cuvette and equilibrated at 25 °C. Tris buffer, although not ideal for CD studies, was chosen because phosphate buffer precipitated in the presence of polyamines. Spectra were recorded in the range of 195–260 nm with a data pitch of 1 nm and a slit width of 1.5 nm. The data points exceeding 700 V applied to the photomultiplier tube were discarded. Titrations were performed by sequential additions of stock polyamine solutions or salt solutions in 5 mM Tris-HCl and 25 mM NaCl adjusted to pH 7.5. The reported spectra were buffer-subtracted and corrected for dilution caused by added titrant.
Nuclear Magnetic Resonance (NMR) Spectroscopy
1H NMR spectra were recorded for 100 μM acidic-(HhH)2 in 25 mM fully deuterated Tris (Tris-d11) (pH 7.5) and 25 mM NaCl in a 90% H2O/10% D2O mixture and upon addition of various spermine concentrations. Titrations were performed by sequential additions of a pH-adjusted, 1 M spermine solution in 25 mM NaCl and a 90% H2O/10% D2O mixture. NMR experiments were conducted at 293 K on a Bruker AVANCE NEO 600 MHz NMR spectrometer equipped with a 5 mm cryogenic triple-resonance HCN TCI probe (triple-axis X, Y, and Z gradients). Data were processed and analyzed using TOPSPIN 4.0 (Bruker BioSpin). The one-dimensional 1H NMR spectrum was acquired using excitation sculpting33 to suppress the solvent signal; the two-dimensional (2D) TOCSY spectrum (total correlation spectroscopy34) at the spermine saturating concentration (1:250 protein:spermine molar ratio) was recorded using a dipsi2 mixing time of 120 ms.
Results
Design of Acidic-(HhH)2
To determine if polyamines can induce folding of a primordial, hyperacidic protein, we designed acidic-(HhH)2 (Figure 2) on the basis of our recently published symmetric-(HhH)2 protein.27 Briefly, symmetric-(HhH)2 was the result of ancestral inference and targeted simplification of the ancient and widely distributed (HhH)2 protein fold. The (HhH)2 protein fold is an α-helix bundle formed by two symmetrically juxtaposed helix–hairpin–helix (HhH) motifs.35 The HhH motif is a pre-LUCA structural element32 that interacts with nucleic acids. It is part of numerous proteins, including ribosomal proteins and polymerases, but when duplicated can form a stand-alone domain. The sequence of symmetric-(HhH)2 is derived from the symmetrization of the reconstructed ancestor of all known (HhH)2 protein families. As the name implies, the sequences of the first and second HhH subdomains of symmetric-(HhH)2 are identical. Symmetric-(HhH)2 therefore represents an intermediate along the trajectory leading from a primordial single HhH polypeptide to a contemporary (HhH)2 protein domain. Although the more simplified primordial-(HhH)2 construct was the preferred background for an acidic variant, as it is comprised of only prebiotic amino acids, the resulting acidic variant responded weakly to the presence of polyamines, as detailed below.
To generate acidic-(HhH)2, all of the lysine residues in symmetric-(HhH)2 were mutated to glutamate, an abiotic, early-emerging amino acid.31 Glutamate was chosen over aspartate because it has a higher α-helix propensity36 and because this mutation is preferred in the BLOSSOM62 substitution matrix.37 Twelve positions were exchanged in symmetric-(HhH)2 to give acidic-(HhH)2. The resulting protein is 60 residues long, has 100% sequence identity between the two HhH domains, and is comprised of just 12 amino acid types. In total, acidic-(HhH)2 has 21 glutamate residues, no arginine or lysine, and just two histidine residues. Consequently, acidic-(HhH)2 has a pI of 3.6 and bears a significant negative charge at neutral pH. In buffer at pH 7.5, acidic-(HhH)2 is unfolded, as demonstrated by the random coil signal in the CD spectrum (Figure 3) and the poor peak dispersion in the one-dimensional (1D) 1H NMR spectrum (Figure 4, top).
Figure 3.
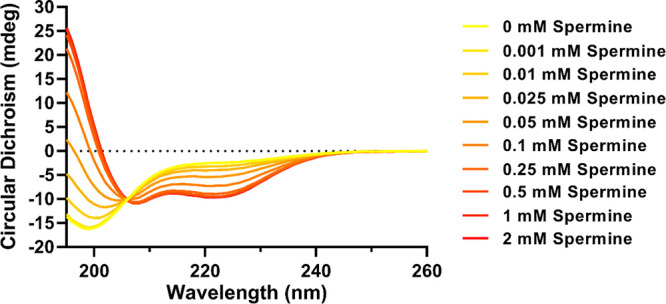
Folding of acidic-(HhH)2 upon spermine addition monitored by circular dichroism (CD). Shown are CD spectra of 5 μM acidic-(HhH)2 with spermine at varying concentrations. Each curve represents the average of two scans after buffer subtraction [5 mM Tris (pH 7.5) and 25 mM NaCl] and correction for dilution by added titrant. The development of a peak at <195 nm and negative bands at ∼208 and ∼222 nm suggests folding into a predominantly α-helical conformation. An isodichroic point at ∼206 nm suggests a two-state transition.
Figure 4.

Titration of acidic-(HhH)2 with spermine monitored by NMR. 1D 1H NMR spectra of 100 μM acidic-(HhH)2 in varying concentrations of spermine (0, 1, 5, and 25 mM). The increasing peak dispersion and observation of new peaks upon addition of spermine (highlighted with a gray background) both indicate that spermine is promoting the acquisition of structure. (A) Close-up of the spectral region reporting amide and aromatic protons. (B) Spectral region corresponding to aliphatic protons, predominantly methyl groups.
Spermine Induces α-Helix Formation
The effect of spermine, a natural polyamine with four amino groups separated by alkyl chains (Figure 1), on the structure of acidic-(HhH)2 was first monitored by CD spectroscopy. Upon addition of submillimolar concentrations of spermine, signals associated with α-helical structure rapidly developed in the CD spectrum during the dead time of the experiment (Figure 3 and Figure S1). Titration of acidic-(HhH)2 with spermine yielded a midpoint of the folding transition of ∼70 μM (in Figure 5, see the legend). The spectra from the spermine titration also revealed the presence of an isodichroic point at ∼206 nm, consistent with a simple two-state transition between a random coil conformation and an α-helical conformation. The low concentration at which we observed the folding transition indicates that the effect of spermine is not simply from screening negative charge. The circular dichroism buffer includes 25 mM NaCl (ionic strength of 25 mM), and addition of 0.1 mM spermine increased the ionic strength by just 1 mM (assuming spermine is completely protonated and neutralized by chloride ions).
Figure 5.
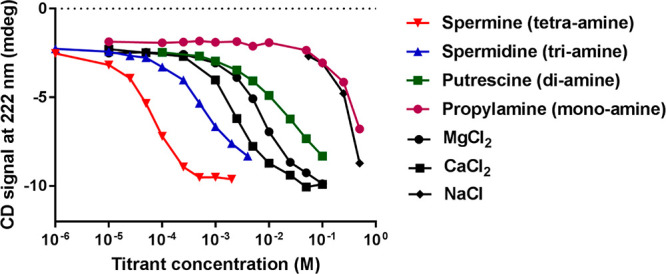
Titration of acidic-(HhH)2 with various polyamines and salts. Circular dichroism spectra of 5 μM acidic-(HhH)2 upon addition of various polyamines and salts were recorded (Figure S5). Plotted here is the CD signal at 222 nm, a reporter of α-helical structure. Empirical midpoint concentrations are 0.07 mM spermine, 0.7 mM spermidine, 21 mM putrescine, 420 mM propylamine, 2.6 mM CaCl2, 7.4 mM MgCl2, and 300 mM NaCl. The midpoint concentrations were estimated from a linear interpolation between points and assuming a folded signal of approximately −10 mdeg at 222 nm. Instead of a technical duplicate, we performed a second polyamine titration experiment with an independent protein preparation at twice the protein concentration, presented in Figure S4. The midpoint concentrations between the two titrations are within 20%, and the order of potency is the same.
In contrast to acidic-(HhH)2, the acidified variant of primordial-(HhH)2 responded only weakly to spermine, with concentrations of >1 mM spermine inducing only modest conformational changes (Figure S2A). However, the failure of this construct to fold upon polyamine addition seems to stem from the loss of specific interactions rather than the perturbation of surface charges. Symmetric-(HhH)2 retains two His-Glu interaction pairs that bridge the two HhH subdomains and were previously proven to be critical. Upon simplification to give primordial-(HhH)2, these histidine residues had to be replaced by arginine to maintain folding and function. We accordingly constructed an acidified form of primordial-(HhH)2 in which the positions corresponding to the two histidines were kept as arginine, but the other 12 basic residues were replaced with glutamate . The resulting protein behaved like acidic-(HhH)2: it was unfolded in buffer yet assumed an α-helical structure upon the addition of spermine (Figure S2B).
To cross validate our results from circular dichroism, the titration of acidic-(HhH)2 with spermine was followed by 1H NMR. In the absence of spermine, acidic-(HhH)2 exhibits poor chemical shift dispersion (Figure 4, top). This observation is indicative of an unfolded state, as the dispersion of the 1H resonances, and specifically of the 1HN, 1Hα, and methyl signals, is much greater in folded proteins, reflecting the well-known dependence of the environment of proton nuclei on secondary and tertiary structure.38 Titration of acidic-(HhH)2 with spermine caused an increase in the 1H chemical shift dispersion (Figure 4). The observation of new 1H signals upon the addition of spermine at >8.7 ppm, < 0.6 ppm, and between 7.3 and 7.6 ppm (Figure 4, peaks highlighted in gray) is in agreement with folding upon spermine binding. The appearance of methyl resonances at chemical shifts outside the “random coil” range (Figure 4B) and the increased dispersion of amide proton resonances in particular (Figure 4A) indicate a change in the chemical environment of these nuclei, most likely due to changes in structure. Chemical shifts of backbone protons tend to be shifted upfield for α-helices, by an average of −0.35 ppm relative to random coil values, versus downfield for β-sheets, by an average of 0.40 ppm.39,40 Thus, the lack of new signals upon spermine addition in the 4.85–5.90 ppm region is consistent with the absence of β-sheet structure, while the observation of 1Hα signals at 3.8–4.4 ppm supports the presence of an α-helical structure (Figure S3). In the so-called “fingerprint region” of the 2D TOCSY spectrum (marked box in Figure S3), one would expect to observe one signal per residue for all non-proline residues (excluding the N-terminal residue). However, the presence of 21 glutamic acid residues, a duplicated sequence, and some spectral broadening—perhaps due to transient associations between protein molecules at the high protein concentration used for this experiment and in the presence of spermine—led to high 1HN resonance overlap, thus precluding the assignment of the 1H NMR peaks with this unlabeled protein sample. Nevertheless, the NMR data qualitatively support the chemical chaperone effects of spermine, where, in agreement with the CD analysis presented above, the structure of acidic-(HhH)2 becomes more ordered and adopts a predominantly α-helical conformation upon addition of spermine.
Other Polyamines and Dications Mediate Folding of Acidic-(HhH)2
To better understand the chemical chaperone properties of polyamines, we performed titrations with natural polyamines of different length, as well as a monoamine (propylamine) (Figure 5 and Figure S4). Propylamine and NaCl had similar effects on the folding of acidic-(HhH)2, both exhibiting a folding midpoint concentration at ∼0.4 M and complete folding at ∼1 M. At approximately molar concentrations of salt, both charge masking and excluded volume effects can be significant41 (Figure S5). In contrast, the chemical chaperone effect of polyamines is achieved at significantly lower concentrations (Figure 5). The strength of the chemical chaperone effect scaled with the number of amines per molecule, with spermine (a tetraamine) exhibiting the strongest chemical chaperone and putrescine (a diamine) exhibiting the weakest effect. The transition between putrescine (a diamine) and spermidine (a triamine) resulted in the greatest change in the apparent folding midpoint with an ∼30-fold increase in potency. Finally, although ∼100-fold less potent than the most potent polyamine (spermine), the dications Mg2+ and Ca2+ were far more active than either NaCl or propylamine, and even slightly more active than putrescine.
Discussion
Acidic-(HhH)2 is a model primordial protein with a highly acidic surface. The ability of polyamines to efficiently fold acidic-(HhH)2 suggests a possible role for polyamines as a chemical chaperone early in protein evolution, before basic amino acids could be readily incorporated into proteins. We note that other solutions to the basic amino acid problem have been reported, including the use of ornithine as an alternative basic amino acid27 or a reliance on high concentrations of salt to drive protein folding,42 an effect we observe here, as well. Similarly, phosphate binding sites are generally enriched for basic amino acids in modern proteins; however, in the most ancient protein lineages, phosphate binding is mediated by backbone interactions at the N-termini of α-helices,43 a feature also seen in the (HhH)2 fold.44 Dications, and specifically Mg2+ and Ca2+, also comprise an alternative solution, and although they were far less potent than spermine, they induced folding of acidic-(HhH)2 at approximately seawater concentrations.45 The prebiotic soup, which may have some resemblance to modern seawater, may have contained multiple chemical species that support the folding of simple, hyperacidic proteins and nucleic acids. Indeed, evolution in general, and protein evolution in particular, is renowned for leveraging multiple solutions to tackle the same challenge. One need only to consider the great wealth of unrelated proteases that adopt different folds and employ different nucleophiles.46 In our view, an abundance of accessible solutions, which are complementary rather than contradictory, may well be a prerequisite for the emergence of life.
While a highly acidic surface seems to be well tolerated if neutralized by polyamines, specific electrostatic interactions may not be replaceable. The His-Glu interactions in acidic-(HhH)2 and the corresponding Arg-Glu interactions in the acidified version of primordial-(HhH)2 are crucial for (HhH)2 folding and cannot be substituted by polyamines. In fact, a similar phenomenon was observed by Hollecker and Creighton where, although chemical acidification of lysine side chains was generally well tolerated, the modification of certain positions resulted in complete unfolding.23 Furthermore, the fact that spermine-induced folding does occur in an acidic version of primordial-(HhH)2 that retains the interacting arginine residues supports our interpretation that this interaction is crucial for the stability of the (HhH)2 fold and also that acidic-(HhH)2 does indeed adopt the (HhH)2 fold upon addition of spermine.
Although framed and tested from the perspective of an acidic, primordial protein, these results lead us to hypothesize that polyamine-induced folding could be retained in modern biology, especially because the chemical chaperone benefits are observed within a biologically relevant concentration range, particularly for spermine (1–2 mM in mammalian cells47) and spermidine (6–7 mM in E. coli(8)), though it is unknown what fraction of molecules are bound. In mammalian cells, the concentrations of various polyamines oscillate in accordance with the circadian rhythm,7 suggesting a role for polyamines in signaling and regulation.9,48 Polyamines have also been shown to regulate individual proteins, such as a viral serine kinase49 and the circadian regulation protein PER2.7 However, the mechanistic and structural basis of these dependencies is unknown. Finally, we note that the high concentration of polyamines in bacteria is compatible with, and potentially related to, their generally acidic proteomes;50 hence, polyamines may have proteome-wide effects. We demonstrate that, mechanistically, polyamines can operate at concentrations with negligible ionic strength or excluded volume effects. Hence, specific binding interactions between structurally adjacent acidic residues are likely to be responsible for the chemical chaperone effects. Protein regions with high α-helicity that are enriched for acidic amino acids may therefore be a hallmark of a polyamine-responsive protein element.
Further studies are needed to support the hypotheses described above, and also to examine whether hyperacidic proteins could not only fold in the presence of polyamines, as demonstrated here, but also exert function. We could not observe dsDNA binding by acidic-(HhH)2 in the presence of polyamines (data not shown), but we note that such measurements are technically challenging because polyamines bind nucleic acids on their own and the functional output of highly simplified proteins is often weak. Nonetheless, other model proteins may provide indications for function, thus lending further support for a role of polyamines at the earliest stages of protein evolution.
Supporting Information Available
The Supporting Information is available free of charge at https://pubs.acs.org/doi/10.1021/acs.biochem.0c00800.
Supporting figures (PDF)
Author Contributions
D.D. and L.M.L. contributed equally to this work. I.G.-S. proposed the idea of polyamines as substitutes to basic amino acids in the early proteins. D.D., L.M.L., and D.S.T. designed the experiments. D.D., L.M.L., E.A., and A.K. produced protein and performed CD and other biophysical experiments. T.S. performed NMR measurements. D.D., L.M.L., and D.S.T. analyzed data. D.D., L.M.L., and D.S.T. wrote the paper.
This work was funded by Israel Science Foundation Grant 980/14 to D.S.T. D.S.T. is the Nella and Leon Benoziyo Professor of Biochemistry. I.G.-S. acknowledges support by the Croatian Science Foundation (Grant IP-2016-06-6272). T.S. is the incumbent of the Monroy-Marks Research Fellow Chair.
The authors declare no competing financial interest.
Supplementary Material
References
- Michael A. J. (2016) Polyamines in Eukaryotes, Bacteria, and Archaea. J. Biol. Chem. 291, 14896–14903. 10.1074/jbc.R116.734780. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kusano T.; Berberich T.; Tateda C.; Takahashi Y. (2008) Polyamines : Essential Factors for Growth and Survival. Planta 228, 367–381. 10.1007/s00425-008-0772-7. [DOI] [PubMed] [Google Scholar]
- Pasini A.; Caldarera C. M.; Giordano E. (2014) Chromatin Remodeling by Polyamines and Polyamine Analogs. Amino Acids 46, 595–603. 10.1007/s00726-013-1550-9. [DOI] [PubMed] [Google Scholar]
- Nilsson J.; Koskiniemi S.; Persson K.; Grahn B.; Holm I. (1997) Polyamines Regulate Both Transcription and Translation of the Gene Encoding Ornithine Decarboxylase Antizyme in Mouse. Eur. J. Biochem. 250, 223–231. 10.1111/j.1432-1033.1997.0223a.x. [DOI] [PubMed] [Google Scholar]
- Matsufuji S.; Matsufuji T.; Miyazaki Y.; Murakami Y.; Atkins J. F; Gesteland R. F; Hayashi S.-i. (1995) Autoregulatory Frameshifting in Decoding Mammalian Ornithine Decarboxylase Antizyme. Cell 80, 51–60. 10.1016/0092-8674(95)90450-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Landau G.; Ran A.; Bercovich Z.; Feldmesser E.; Horn-Saban S.; Korkotian E.; Jacob-Hirsh J.; Rechavi G.; Ron D.; Kahana C. (2012) Expression Profiling and Biochemical Analysis Suggest Stress Response as a Potential Mechanism Inhibiting Proliferation of Polyamine-depleted Cells. J. Biol. Chem. 287, 35825–35837. 10.1074/jbc.M112.381335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zwighaft Z.; Aviram R.; Shalev M.; Rousso-Noori L.; Kraut-Cohen J.; Golik M.; Brandis A.; Reinke H.; Aharoni A.; Kahana C.; Asher G. (2015) Circadian Clock Control by Polyamine Levels through a Mechanism that Declines with Age Article Circadian Clock Control by Polyamine Levels through a Mechanism that Declines with Age. Cell Metab. 22, 874–885. 10.1016/j.cmet.2015.09.011. [DOI] [PubMed] [Google Scholar]
- Tabor C. W.; Tabor H. (1985) Polyamines in Microorganisms. Microbiol. Rev. 49, 81–99. 10.1128/MMBR.49.1.81-99.1985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pegg A. E. (2009) Mammalian Polyamine Metabolism and Function. IUBMB Life 61, 880–894. 10.1002/iub.230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hasan R.; Alam M. K.; Ali R. (1995) Polyamine Induced Z-Conformation of Native Calf Thymus DNA. FEBS Lett. 368, 27–30. 10.1016/0014-5793(95)00591-V. [DOI] [PubMed] [Google Scholar]
- Feuerstein B. G.; Pattabiraman N.; Marton L. J. (1990) Molecular Mechanics of the Interactions of Spermine with DNA: DNA Bending as a Result of Ligand Binding. Nucleic Acids Res. 18, 1271–1282. 10.1093/nar/18.5.1271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kućan Ž., Naranda T., Plohl M., Nöthig-Laslo V., and Weygand-Durašević I. (1988) in Effect of Spermine on Transfer RNA and Transfer RNA-Ribosome Interactions BT - Progress in Polyamine Research: Novel Biochemical, Pharmacological, and Clinical Aspects (Zappia V., and Pegg A. E., Eds.) pp 525–533, Springer US, Boston. [DOI] [PubMed] [Google Scholar]
- Aumiller W. M.; Pir Cakmak F.; Davis B. W.; Keating C. D. (2016) RNA-Based Coacervates as a Model for Membraneless Organelles: Formation, Properties, and Interfacial Liposome Assembly. Langmuir 32, 10042–10053. 10.1021/acs.langmuir.6b02499. [DOI] [PubMed] [Google Scholar]
- Flink I.; Pettijohn D. E. (1975) Polyamines Stabilise DNA Folds. Nature 253, 62–63. 10.1038/253062a0. [DOI] [PubMed] [Google Scholar]
- Koculi E.; Lee N.-K.; Thirumalai D.; Woodson S. A. (2004) Folding of the Tetrahymena Ribozyme by Polyamines: Importance of Counterion Valence and Size. J. Mol. Biol. 341, 27–36. 10.1016/j.jmb.2004.06.008. [DOI] [PubMed] [Google Scholar]
- Lightfoot H. L.; Hall J. (2014) Endogenous Polyamine Function—the RNA Perspective. Nucleic Acids Res. 42, 11275–11290. 10.1093/nar/gku837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kabir A.; Jash C.; Payghan P. V.; Ghoshal N.; Kumar G. S. (2020) Polyamines and Its Analogue Modulates Amyloid Fibrillation in Lysozyme: A Comparative Investigation. Biochim. Biophys. Acta, Gen. Subj. 1864, 129557. 10.1016/j.bbagen.2020.129557. [DOI] [PubMed] [Google Scholar]
- Simoni E.; Caporaso R.; Bergamini C.; Fiori J.; Fato R.; Miszta P.; Filipek S.; Caraci F.; Giuffrida M. L.; Andrisano V.; Minarini A.; Bartolini M.; Rosini M. (2016) Polyamine Conjugation as a Promising Strategy To Target Amyloid Aggregation in the Framework of Alzheimer’s Disease. ACS Med. Chem. Lett. 7, 1145–1150. 10.1021/acsmedchemlett.6b00339. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lewandowski N. M.; Ju S.; Verbitsky M.; Ross B.; Geddie M. L.; Rockenstein E.; Adame A.; Muhammad A.; Vonsattel J. P.; Ringe D.; Cote L.; Lindquist S.; Masliah E.; Petsko G. A.; Marder K.; Clark L. N.; Small S. A. (2010) Polyamine Pathway Contributes to the Pathogenesis of Parkinson Disease. Proc. Natl. Acad. Sci. U. S. A. 107, 16970–16975. 10.1073/pnas.1011751107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baeza I.; Ibanez M.; Wong C.; Chavez P.; Gariglio P.; Oro J. (1992) Possible Prebiotic Significance of Polyamines in the Condensation, Protection, Encapsulation, and Biological Properties of DNA. Origins Life Evol. Biospheres 21, 225–242. 10.1007/BF01809858. [DOI] [PubMed] [Google Scholar]
- Wong C.; Santiago J. C.; Rodriguez-Paez L.; Ibanez M.; Baeza I.; Oro J. (1991) Synthesis of Putrescine Under Possible Primitive Earth Conditions. Origins Life Evol. Biospheres 21, 145–156. 10.1007/BF01809443. [DOI] [PubMed] [Google Scholar]
- Kumar S.; Tsai C.-J.; Ma B.; Nussinov R. (2000) Contribution of Salt Bridges Toward Protein Thermostability. J. Biomol. Struct. Dyn. 17, 79–85. 10.1080/07391102.2000.10506606. [DOI] [PubMed] [Google Scholar]
- Hollecker M.; Creighton T. E. (1982) Effect on protein stability of reversing the charge on amino groups. Biochim. Biophys. Acta, Protein Struct. Mol. Enzymol. 701, 395. 10.1016/0167-4838(82)90243-6. [DOI] [PubMed] [Google Scholar]
- Lawrence M. S.; Phillips K. J.; Liu D. R. (2007) Supercharging Proteins Can Impart Unusual Resilience. J. Am. Chem. Soc. 129, 10110–10112. 10.1021/ja071641y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnson A. P.; Cleaves H. J.; Dworkin J. P.; Glavin D. P.; Lazcano A.; Bada J. L. (2008) The Miller Volcanic Spark Discharge Experiment. Science 322, 404. 10.1126/science.1161527. [DOI] [PubMed] [Google Scholar]
- Miller S. L. (1953) A Production of Amino Acids Under Possible Primitive Earth Conditions. Science 117, 528–529. 10.1126/science.117.3046.528. [DOI] [PubMed] [Google Scholar]
- Longo L. M.; Despotović D.; Weil-Ktorza O.; Walker M. J.; Jabłońska J.; Fridmann-Sirkis Y.; Varani G.; Metanis N.; Tawfik D. S. (2020) Primordial Emergence of a Nucleic Acid Binding Protein via Phase Separation and Statistical Ornithine-to-Arginine Conversion. Proc. Natl. Acad. Sci. U. S. A. 117, 15731–15739. 10.1073/pnas.2001989117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hartman H. (1975) Speculations on the Origin and Evolution of Metabolism. J. Mol. Evol. 4, 359–370. 10.1007/BF01732537. [DOI] [PubMed] [Google Scholar]
- Longo L. M.; Blaber M. (2014) Prebiotic Protein Design Supports a Halophile Origin of Foldable Proteins. Front. Microbiol. 10.3389/fmicb.2013.00418. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McDonald G. D.; Storrie-Lombardi M. C. (2010) Biochemical Constraints in a Protobiotic Earth Devoid of Basic Amino Acids: The “BAA(−) World. Astrobiology 10, 989–1000. 10.1089/ast.2010.0484. [DOI] [PubMed] [Google Scholar]
- Trifonov E. N. (2000) Consensus Temporal Order of Amino Acids and Evolution of the Triplet Code. Gene 261, 139–151. 10.1016/S0378-1119(00)00476-5. [DOI] [PubMed] [Google Scholar]
- Alva V.; Söding J.; Lupas A. N. (2015) A Vocabulary of Ancient Peptides at the Origin of Folded Proteins. eLife 4, e09410. 10.7554/eLife.09410. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hwang T. L.; Shaka A. J. (1995) Water Suppression That Works. Excitation Sculpting Using Arbitrary Wave-Forms and Pulsed-Field Gradients. J. Magn. Reson., Ser. A 112, 275–279. 10.1006/jmra.1995.1047. [DOI] [Google Scholar]
- Braunschweiler L.; Ernst R. R. (1983) Coherence Transfer by Isotropic Mixing: Application to Proton Correlation Spectroscopy. J. Magn. Reson. 53, 521–528. 10.1016/0022-2364(83)90226-3. [DOI] [Google Scholar]
- Brennan R. G.; Matthews B. W. (1989) The Helix-Turn-Helix DNA Binding Motif. J. Biol. Chem. 264, 22–25. [PubMed] [Google Scholar]
- Nick Pace C.; Martin Scholtz J. (1998) A Helix Propensity Scale Based on Experimental Studies of Peptides and Proteins. Biophys. J. 75, 422–427. 10.1016/S0006-3495(98)77529-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Henikoff S.; Henikoff J. G. (1992) Amino Acid Substitution Matrices From Protein Blocks. Proc. Natl. Acad. Sci. U. S. A. 89, 10915–10919. 10.1073/pnas.89.22.10915. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Case D. A. (2000) Interpretation of Chemical Shifts and Coupling Constants in Macromolecules. Curr. Opin. Struct. Biol. 10, 197–203. 10.1016/S0959-440X(00)00068-3. [DOI] [PubMed] [Google Scholar]
- Dalgarno D. C.; Levine B. A.; Williams R. J. P. (1983) Structural Information From NMR Secondary Chemical Shifts of Peptide α C-H Protons in Proteins. Biosci. Rep. 3, 443–452. 10.1007/BF01121955. [DOI] [PubMed] [Google Scholar]
- Jiménez M. A.; Nieto J. L.; Herranz J.; Rico M.; Santoro J. (1987) 1H NMR and CD Evidence of the Folding of the Isolated Ribonuclease 50–61 Fragment. FEBS Lett. 221, 320–324. 10.1016/0014-5793(87)80948-1. [DOI] [PubMed] [Google Scholar]
- Gibb B. C. (2019) Hofmeister’s Curse. Nat. Chem. 11, 963–965. 10.1038/s41557-019-0355-1. [DOI] [PubMed] [Google Scholar]
- Longo L. M.; Lee J.; Blaber M. (2013) Simplified Protein Design Biased for Prebiotic Amino Acids Yields a Foldable, Halophilic Protein. Proc. Natl. Acad. Sci. U. S. A. 110, 2135–9. 10.1073/pnas.1219530110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Longo L. M.; Petrović D.; Kamerlin C. L.; Tawfik D. S. (2020) Short and Simple Sequences Favored the Emergence of N-Helix Phospho- Ligand Binding Sites in the First Enzymes. Proc. Natl. Acad. Sci. U. S. A. 117, 5310–5318. 10.1073/pnas.1911742117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Doherty A. J.; Serpell L. C.; Ponting C. P. (1996) The Helix-Hairpin-Helix DNA-Binding Motif: A Structural Basis for Non-Sequence-Specific Recognition of DNA. Nucleic Acids Res. 24, 2488–2497. 10.1093/nar/24.13.2488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Besson P.; Degboe J.; Berge B.; Chavagnac V.; Fabre S.; Berger G. (2014) Calcium, Na, K and Mg Concentrations in Seawater by Inductively Coupled Plasma-Atomic Emission Spectrometry: Applications to IAPSO Seawater Reference Material, Hydrothermal Fluids and Synthetic Seawater Solutions. Geostand. Geoanal. Res. 38, 355–362. 10.1111/j.1751-908X.2013.00269.x. [DOI] [Google Scholar]
- Neurath H. (1984) Evolution of Proteolytic Enzymes. Science 224, 350–357. 10.1126/science.6369538. [DOI] [PubMed] [Google Scholar]
- Watanabe S.; Kusama-Eguchi K.; Kobayashi H.; Igarashi K. (1991) Estimation of Polyamine Binding to Macromolecules and ATP in Bovine Lymphocytes and Rat Liver. J. Biol. Chem. 266, 20803–20809. [PubMed] [Google Scholar]
- Miller-Fleming L.; Olin-Sandoval V.; Campbell K.; Ralser M. (2015) Remaining Mysteries of Molecular Biology: The Role of Polyamines in the Cell. J. Mol. Biol. 427, 3389–3406. 10.1016/j.jmb.2015.06.020. [DOI] [PubMed] [Google Scholar]
- Kenyon T. K.; Lynch J.; Hay J.; Ruyechan W.; Grose C. (2001) Varicella-Zoster Virus ORF47 Protein Serine Kinase : Characterization of a Cloned, Biologically Active Phosphotransferase and Two Viral Substrates, ORF62 and ORF63. J. Virol. 75, 8854–8858. 10.1128/JVI.75.18.8854-8858.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kozlowski L. P. (2017) Proteome-pI: Proteome Isoelectric Point Database. Nucleic Acids Res. 45, D1112–D1116. 10.1093/nar/gkw978. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blagbrough I. S., Metwally A. A., and Geall A. J. (2011) Measurement of Polyamine pKa Values. In Polyamines: Methods and Protocols (Pegg A. E., and Casero R. A. Jr., Eds.) pp 493–503, Humana Press, Totowa, NJ. [DOI] [PubMed] [Google Scholar]
- Kimberly M. M.; Goldstein J. H. (1981) Determination of pKa Values and Total Proton Distribution Pattern of Spermidine by Carbon-13 Nuclear Magnetic Resonance Titrations. Anal. Chem. 53, 789–793. 10.1021/ac00229a011. [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.