

Abstract
Older adults comprise the fastest growing global demographic and are at increased risk of poor mental health outcomes. Although aerobic exercise and sleep are critical to the preservation of emotional well-being, few studies have examined their combined mood-enhancing effects, or the potential neural mechanisms underlying these effects. Here, we used a randomized crossover design to test the impact of acute exercise on mood and the intrinsic functional connectivity (iFC) of the cingulo-opercular network in physically healthy older adults. Wrist actigraphy provided objective indices of sleep. Results revealed that 30 min of moderate-intensity aerobic exercise acutely enhanced positive affect (PA) and reduced iFC between the cingulo-opercular network and the hippocampus. Both effects were magnified among older adults with greater sleep disturbance. Exercise-induced changes in hippocampal iFC mediated relations between sleep disturbance and exercise-induced increases in PA. These findings provide evidence that aerobic exercise enhances mood, that it does so by altering connectivity between the anterior insula—a key hub in the cingulo-opercular network—and the hippocampus and that lower sleep quality is a stronger predictor of these effects among older adults. These observations underscore the benefits of moderate-intensity exercise—a safe and scalable behavioral intervention—and provide new clues about the neural circuitry underlying the interactive effects of sleep and exercise on mood.
Keywords: sleep actigraphy, acute exercise, affect, cingulo-opercular network
Introduction
Adults over the age of 65 years constitute the fastest growing demographic of the global population (Wahl et al., 2019). By 2050, this segment of society is expected to exceed two billion people, effectively doubling its current size (Newgard and Sharpless, 2013). While greater longevity affords longer periods of productive and meaningful life, older age confers increased risk for a range of negative health outcomes (Kennedy et al., 2014). Poor emotional well-being—including lower levels of positive affect (PA) and elevated depression, the leading cause of disability worldwide (Friedrich, 2017)—is common among older adults (Rodda et al., 2011) and has far-reaching impacts on health by exacerbating chronic disease [i.e., cardiovascular disease (Dhar and Barton, 2016), cancer (Spiegel and Giese-Davis, 2003), diabetes (Mezuk et al., 2008), arthritis (Dickens et al., 2002) and Alzheimer’s disease (Ownby et al., 2006)].
Growing evidence indicates that exercise is critically important for the preservation of emotional well-being among older adults (Rhyner and Watts, 2016). Studies have shown that both aerobic exercise and resistance training can diminish depression (Gordon et al., 2018; Morres et al., 2019) and that a single 10- to 35-min session of light-to-moderate intensity exercise can acutely amplify PA (Ekkekakis and Petruzzello, 1999; Reed and Ones, 2006). Moreover, several studies have shown that acute and chronic aerobic exercises improve sleep in midlife to later life (King et al., 1997; Kline et al., 2011; Santos et al., 2012).
Like exercise, sleep is implicated in emotion regulation and mental health outcomes across the life span (Steptoe et al., 2008), and changes in sleep are common with aging (Mander et al., 2017; Li et al., 2018). On average, middle-aged and older adults have shorter sleep duration, lower sleep efficiency (i.e., percentage of time in bed asleep) and more sleep complaints (Mander et al., 2017; Li et al., 2018) than their younger counterparts. Among older women, lower sleep efficiency and more awake time after sleep onset (WASO) are associated with elevated anxiety (Spira et al., 2009), and older adults with insomnia are more likely to report negative emotions (Buysse, 2004). From a mechanistic perspective, acute sleep deprivation has been shown to increase loneliness (Simon and Walker, 2018), amplify amygdala reactivity (Krause et al., 2017) and alter the activity of the cingulo-opercular network—a key mediator of emotion regulation and detection of salient stimuli (Krause et al., 2017).
Despite compelling evidence linking sleep and exercise to emotional well-being, few studies have examined their combined effects. Although existing work provides preliminary evidence for interactive effects, for example by showing that the salubrious impact of chronic exercise on post-traumatic stress symptoms are magnified among those with poor sleep (Babson et al., 2015), little is known about whether or not these interactions occur in response to single sessions of exercise, or the neural circuitry that may underlie these interactions. Converging lines of evidence point to the importance of cingulo-opercular (‘salience’) network intrinsic functional connectivity (iFC) as a potential modifiable target. Anchored by the anterior insula (AI)/frontal operculum and midcingulate cortex (MCC), the cingulo-opercular network is a distinct network that detects and filters sensory information and is thought to play a critical role in emotion regulation and salience detection (Menon and Uddin, 2010). Other works highlight the potential relevance of this network to mood and anxiety disorders (Furman et al., 2011; Andreescu et al., 2015; Koch et al., 2016) and insomnia (Chen et al., 2014). The cingulo-opercular network projects to several subcortical brain regions that are sensitive to exercise, including afferent projections to the hippocampus (Erickson et al., 2011; Arszovszki et al., 2014; Alfini et al., 2016). Collectively, these observations motivate the hypotheses that the mood-enhancing effects of acute exercise are magnified among those with disturbed sleep and that these effects might be mediated by alterations in cingulo-opercular network iFC.
Here, we used a novel combination of approaches to rigorously evaluate the joint effects of sleep and acute exercise on affect and cingulo-opercular network iFC in older adults. Wrist actigraphy provided objective indices of sleep disturbance (WASO) and duration [total sleep time (TST)], which were selected because they are two key dimensions of sleep that can be derived from wrist actigraphy and are associated with emotional well-being (Faubel et al., 2009; Lemola et al., 2013; Wrzus et al., 2014; Tazawa et al., 2019). A randomized crossover design enabled us to test the impact of a 30-min aerobic exercise intervention on mood and cingulo-opercular network connectivity, relative to a well-established sedentary control condition. Integrating these data streams enabled us to determine whether (i) acute exercise enhances PA, (ii) this emotional ‘boost’ is magnified among older adults with greater sleep disturbance and (iii) these emotional effects emerge from alterations in the functional connectivity of the cingulo-opercular network.
To assess specificity, we also examined the impact of sleep quality and acute exercise on negative affect (NA) and on frontoparietal (‘executive’) network connectivity; particularly because prior research has demonstrated an effect of exercise on frontoparietal network iFC and the dorsal lateral prefrontal cortex (dlPFC)—one of its primary hubs (Fang et al., 2016; Prehn et al., 2019). Moreover, like the cingulo-opercular network, frontoparietal network iFC has been implicated in executive control and emotion regulation (Golkar et al., 2012; Prehn et al., 2019). Addressing these questions has the potential to inform the development of safe, scalable and low-cost behavioral interventions for improving the health and well-being of older adults, and to provide novel targets for mechanistic work in humans or animals.
Method
Participants
Thirty-two physically active, cognitively normal older adults (75% women, age 55–81 years) participated in the current study (Table 1). Recruitment strategies included in-person informational sessions at local recreation centers, regional newspaper advertisements and study fliers posted on university listservs. Participants were screened using a structured telephone interview to determine health history and screen for potential contraindications. Qualified individuals obtained their physicians’ approval for moderate-intensity exercise and participated in a laboratory screening session, where they were familiarized with the procedures and provided informed written consent (Figure 1). This study was approved by the Institutional Review Board of the University of Maryland in accordance with the Helsinki Declaration.
Table 1.
Participant characteristics
| Characteristics | Mean SD SD |
|---|---|
| Age (years) | 66.3 ± 7.3 |
| Female | 24 (75.0) |
| Education >12 years | 30 (93.8) |
| White | 25 (78.1) |
| Body mass index (kg/m2) | 25.6 ± 4.2 |
| 7-Day physical activity (kcal/kg/day) | 221.9 ± 28.4 |
| BDI-II | 1.9 ± 1.9 |
| STAI-Trait | 46.6 ± 3.8 |
| MMSE | 29.2 ± 1.1 |
| STOP-BANG | 1.6 ± 0.7 |
| Total sleep time (min) | 406.3 ± 70.4 |
| Wake after sleep onset (min) | 34.4 ± 23.0 |
| Actigraphic data collection (days) | 8.1 ± 3.5 |
Note. Female, education and white are expressed as n (%).
Abbreviations. BDI-II; Beck Depression Inventory-II; STAI-T, Spielberger State and Trait Anxiety Inventory, Trait version; MMSE, Mini-Mental State Exam; STOP-BANG, Snoring, Tired, Observed, Pressure-Body, Age, and Gender questionnaire.
Fig. 1.
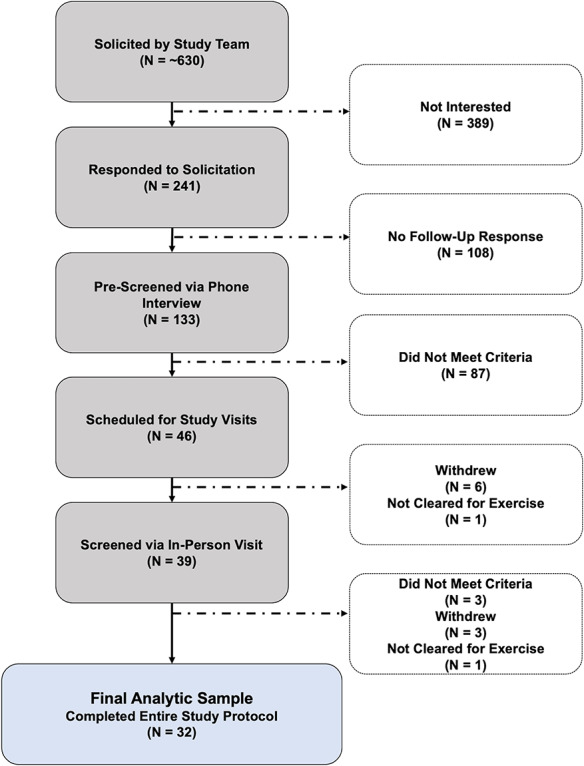
Participant recruitment, screening, exclusion and enrollment between 2016 and 2018.
Eligibility criteria
Individuals were excluded if they reported a history of heart attack, stroke, transient ischemic attack, seizures, epilepsy, brain tumor, closed head injury, alcohol/substance abuse, psychosis; current visual/auditory limitations; or a current diagnosis of Alzheimer’s disease, atrial fibrillation, cardiovascular disease, coronary artery disease, diabetes, hypertension or Parkinson’s disease. We attempted to exclude individuals with probable obstructive sleep apnea using the Snoring Tiredness Observed Pressure-Body Age Neck Gender questionnaire (scores >4) (Chung et al., 2016), elevated depression using the Beck Depression Inventory-II (>13) (Beck et al., 1996), elevated trait anxiety using the State Trait Anxiety Inventory (>54) (Spielberger et al., 1970), significant cognitive impairment using the Mini-Mental Status Examination (<24) (Folstein et al., 1983), left-handedness using the Edinburgh Handedness Inventory (<0) (Oldfield, 1971), severe obesity (body mass index ≥40 kg/m2), low physical activity (<30 min of physical activity three times per week during the past 6 months), limited English language proficiency and magnetic resonance imaging (MRI) contraindications.
Sleep assessment
Prior to the first imaging assessment, participants were fitted with and instructed to wear an actigraph (Motionlogger Watch; Ambulatory Monitoring, Inc., Ardsley, New York) on their non-dominant wrist for seven consecutive 24-h intervals. We collected actigraphic data in 1-min epochs and required a minimum of 3 nights of usable data from each participant. To facilitate data processing, participants were instructed to press an event-marker button on the actigraph when they went to bed (with the intention of sleeping) and again after rising. Daily sleep diaries were used as an ancillary source of information, and served as a means of identifying the in-bed interval, from which the actigraphic sleep measures were derived. Participants were instructed to log the time in the evening they got into bed with the intention of sleeping, and the time in the morning they arose with the intention of beginning their day (Alfini et al., 2020). Participants were also instructed to log actigraph removals (e.g., due to bathing or swimming) each evening. Actigraphic data and diary information were visually inspected by two (nonblinded) raters to identify invalid data (i.e., non-wear periods and periods of device malfunction). Inter-rater discrepancies were resolved via consensus. Action-W software (Ambulatory Monitoring, Inc.) was used to automatically quantify sleep disturbance (WASO; total number of minutes awake after sleep onset) and duration (TST; total number of minutes asleep while in bed), which were averaged across nights of actigraphy.
Acute exercise and sedentary control procedures
Immediately prior to the imaging assessment, participants engaged in either 30 min of moderate-intensity cycling exercise or seated rest (intersession interval, M = 12.1, SD = ±17.0 days). PA and NA were assessed immediately before and after the acute exercise and sedentary control interventions. Sessions were randomized and occurred on separate days. During both interventions, valence (=0.96) and arousal (=0.96) were assessed using the Self-Assessment Manikin (Bradley and Lang, 1994), with higher scores indicating greater valence (emotional feelings of attractiveness/goodness) and arousal. Participants also wore a heart rate (HR) monitor (Polar RS800CX, Polar Electro, Oy, Kempele, Finland) during both interventions. Valence, arousal, HR and rating of perceived exertion (RPE)—indexed using the Borg’s 6–20 RPE scale (Borg, 1990)—were recorded at 5-min intervals, before, during and after each intervention. Participants were provided water ad libitum.
Exercise was performed on a mechanically braked cycle ergometer (Monark 828 E, Monark Exercise AB, Vansbro Sweden) in a testing room at the neuroimaging center. As described in prior publications from our group and others (Rajab et al., 2019b; Won et al., 2019a; Won et al., 2019b), we used an established exercise protocol consisting of 20 min of steady-state exercise flanked by a 5-min self-paced warm-up and cool-down. During exercise, participants were instructed to achieve and sustain an RPE of 15 (i.e., verbal anchor ‘hard’) and a pedal cadence of 60–80 revolutions per minute. Participants were instructed to adjust the ergometer’s resistance to control exercise intensity. After the cool-down, a towel and dry clothing were provided.
Seated rest was completed in a chair in the same testing room. The sedentary rest intervention included 30 min of wakeful rest, during which participants refrained from reading, writing, talking, or using digital devices.
Affect assessment
PA and NA were assessed using the state version of the Positive and Negative Affect Schedule (Watson et al., 1988), a psychometrically sound, 20-item index of PA (=0.91) and NA (=0.69).
MRI assessment
Participants were instructed to remain still, keep their eyes open, and focus on a fixation cross. MRI data were acquired using a Siemens Trio Tim 3.0 Tesla MRI scanner (Erlangen, Germany) and 32-channel head-coil. Sagittal T1-weighted anatomical images were acquired ∼10 min after each intervention using a magnetization prepared rapid gradient echo (MPRAGE) imaging sequence (TR = 1900 ms, TE = 2.3 ms, TI = 900 ms, flip angle = 9°, slice thickness = 0.9 mm, in-plane resolution = 0.9 × 0.9 mm, matrix = 300 × 320, field-of-view = 230 × 256 mm, duration = 4 min 26 s). Echo planar imaging (EPI) volumes were acquired ∼10 min after the completion of the MPRAGE scan (interim scans included field mapping and arterial spin labeling) using a multi-band sequence (TR/TE = 2000/24 ms, flip angle = 70, slice thickness = 3.0 mm, 36 slices, in-plane resolution = 3.0 × 3.0 mm, matrix = 64 × 64, volumes = 240, duration = 8 min 06 s). EPI images were acquired in the oblique axial plane (i.e., 20 relative to the anterior commissure–posterior commissure line) to reduce potential susceptibility artifact.
MRI data processing
T1-weighted images were processed using the FreeSurfer (v. 5.3.0) automated pipeline, which includes tissue segmentation and intensity-based surface deformation. FreeSurfer-rendered anatomical maps were used for co-registration, gray matter isolation and nuisance regression (Khan et al., 2008; Fischl, 2012). The first four volumes of the EPI time-series were discarded to allow for T2* equilibration effects. The resulting time-series was realigned using slice-oriented motion correction (Beall and Lowe, 2014) to mitigate motion artifact. Further processing was performed using AFNI (v. 19.2.06) (Cox, 1996). EPI volumes were de-spiked (3dDespike), slice-time corrected to the beginning of each TR (3dTshift) and motion corrected using the (reference) volume with the least motion (i.e., the minimum outlier fraction). EPI volumes with an outlier fraction >10% or a volume-wise displacement >0.2 mm were censored. The percentage of volumes censored following the exercise condition (2.2%) was not significantly different from the percentage of volumes censored following the rest condition (2.1%) (Z = 0.1, P = 0.918, n = 32). The reference volume was co-registered to the corresponding T1-weighted image (align_epi_anat.py). The T1-weighted image and anatomical followers (FreeSurfer rendered gray matter, white matter and ventricular segmentations) were normalized to the MNI atlas (ICBM 2009c template; 3dQwarp) (Cox and Glen, 2013). The resulting transformation matrix was used to normalize and resample (2 mm3) the co-registered EPI data. To further mitigate artifact, the motion parameters, white matter signal and cerebral spinal fluid signals were included as nuisance regressors, and the residual time-series was bandpass filtered to retain frequencies between 0.01 and 0.1 Hz (3dDeconvolve). After each processing step, data were visually inspected for quality assurance.
First-level fMRI modeling
A seed-based analysis was used to interrogate the cingulo-opercular network. Guided by previous studies (Yeo et al., 2011; Choi et al., 2012), we seeded the AI, one of its primary hubs using 10-mm spheres (left = −34, 22, −2, right = 34, 22, −2). To compute iFC, whole-brain voxelwise regressions were computed separately for each arm of the trial (exercise/control) using the average seed time-series as a predictor (3dfim + and 3dcalc). Voxelwise correlations were normalized using Fisher’s r-to-z transformation. To assess specificity, we also interrogated another intrinsically connected task-positive brain network—the frontoparietal network—using seeds placed in the dlPFC (left = −50, 20, 34, right = 50, 20, 34; Figure 2) (Vincent et al., 2008; Menon and Uddin, 2010; Yeo et al., 2011; Choi et al., 2012; Menon, 2015). The central voxel for both seed masks was determined using the Harvard–Oxford atlas.
Fig. 2.
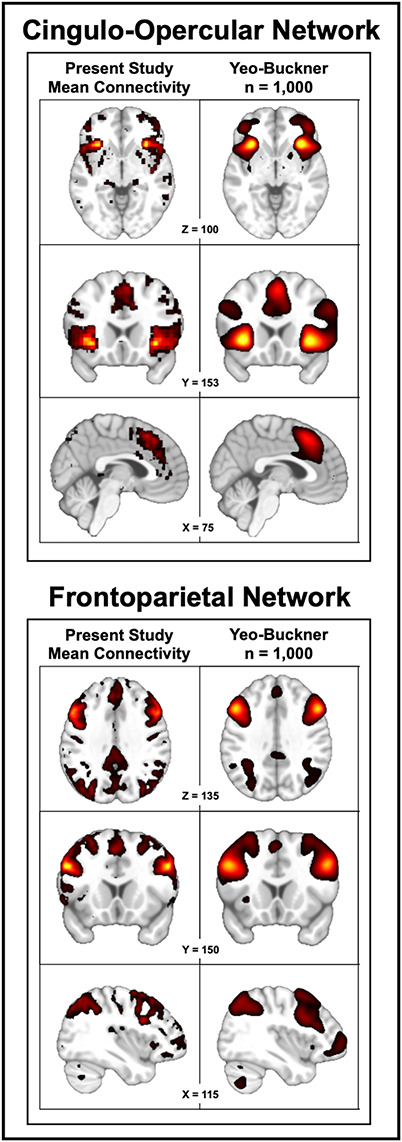
Cingulo-opercular and frontoparietal networks. Upper panel depicts the cingulo-opercular network. Lower panel depicts the frontoparietal network. Left column depicts the networks revealed by the present study (n = 32), averaged across the two assessments. For comparison purposes, the right column depicts the same networks derived using the ‘gold-standard’ Yeo–Buckner data set [n = 1000; (Yeo et al., 2011)]. Visual inspection suggests that the networks are similar, with most differences attributable to the lower spatial resolution of the Yeo–Buckner data set. ‘Gold-standard’ networks were generated using the seed coordinates chosen for the present study in NeuroSynth (Yarkoni et al., 2011). For illustrative purposes, results were arbitrarily thresholded to show the top 12.5% of voxels in each map.
Analytic strategy
Whole-brain tests of voxelwise mean differences.
Voxelwise analyses enabled us to identify the effects of acute exercise on cingulo-opercular network iFC. We merged each participant’s normalized FreeSurfer-rendered GM map to create a group-level intersection mask, within which spatial smoothing, voxelwise analyses and Monte Carlo simulations were performed. We used 3dttest++ to spatially smooth (8 mm) the z-transformed iFC maps and determine the effects of exercise (vs. rest) on cingulo-opercular network iFC. Data were thresholded at P < 0.05, whole-GM corrected for cluster extent (≥1464 mm3) using 3dClustSim and a P = 0.001 cluster-defining threshold (Eklund et al., 2016). To assess specificity, identical procedures were used to assess the impact of the acute exercise intervention on frontoparietal network iFC. Significant voxelwise results were averaged and carried forward to ‘off-line’ regression tests in order to protect against non-independence error.
Brain–behavior relations.
Exploratory data analyses were used to identify potential outliers and evaluate the distributional characteristics of the outcome variables (Tukey, 1977). Analyses were conducted using Stata software (v. 15.1; StataCorp, College Station, TX). Unless otherwise stated, we used mixed-effects models with restricted maximum likelihood and unstructured covariance. Only one sleep measure and one MRI or affective variable was included in each model, and all analyses controlled for potential nuisance variance in mean-centered age and sex. Missing PA data for one participant was imputed using the sample mean. First, we evaluated the Condition × Time interaction on PA. We repeated this analysis for NA to assess specificity. Second, guided by the results of our first objective, we assessed whether variation in objective sleep disturbance (WASO) predicted the effects of exercise on PA. To assess specificity, we repeated this analysis for sleep duration (TST). Third, based on the results of our whole-brain voxelwise analysis, we extracted the average connectivity signal (3dROIstats) from the clusters that showed a significant mean difference between the exercise and rest conditions and used this to compute an iFC change score for these regions (post-exercise − post-rest). Fourth, we used a standard mediation framework (Baron and Kenny, 1986) to test whether exercise-induced differences in iFC statistically explained relations between sleep disturbance (WASO) and exercise-induced changes in PA [(post-exercise − pre-exercise) − (post-rest − pre-rest)]. Here, we used linear regression to test three specific pathways: (i) relations between sleep disturbance (WASO) and brain function; (ii) relations between brain function and exercise-induced changes in PA and (iii) relations between sleep disturbance (WASO) and exercise-induced changes in PA. To minimize the overall number of comparisons, we used a ‘go/no-go’ analytic strategy; only those iFC clusters that demonstrated significant relations for a particular pathway were examined in subsequent tests. We also used the Šidák test to correct for multiple comparisons (on a familywise basis) in multivariable analyses that included more than one test (Šidák, 1967). Because two predictors were included in our first family of multivariable models, a more stringent threshold was enforced for statistical significance (P = 0.0253). Finally, using R (v. 3.6.1), we implemented a well-established nonparametric bootstrapping procedure to formally test mediation (Tingley et al., 2014).
Results
Preliminary analyses confirmed the validity of the acute exercise intervention
As a precursor to hypothesis testing, we confirmed between-condition differences in perceived exertion, valence, arousal and HR (Table 2).
Table 2.
Experimental intervention manipulation check
| Condition | |||
|---|---|---|---|
| Outcome | Exercise | Rest | P (d) |
| Borg’s RPE, mean ± SD | 14.6 ± 1.3 | 6.1 ± 0.3 | <0.001 (0.88) |
| SAM, Valence, mean ± SD | 5.9 ± 2.0 | 6.9 ± 2.1 | 0.008 (0.50) |
| SAM, Arousal, mean ± SD | 5.4 ± 2.1 | 4.3 ± 2.0 | 0.027 (0.41) |
| Heart rate (bpm), mean ± SD | 137.3 ± 19.1 | 66.6 ± 8.7 | <0.001 (3.26) |
Note. RPE, rating of perceived exertion 6–20 scale; SAM, Self-Assessment Manikin; bpm, beats per minute; d, Cohen’s d. Values reflect the scores for each measure averaged over the final 10 min of exercise and rest, respectively. P-values are from paired t-tests or Wilcoxon signed-rank tests.
Acute aerobic exercise enhances PA
Results revealed a significant Condition × Time interaction for PA (B = 2.90, 95% confidence interval [CI] = 0.30–5.50, P = 0.029, n = 32). Acute exercise intervention amplified PA (B = 1.22, 95% CI = 0.27–2.17, P = 0.012, n = 32), whereas rest diminished PA (B = −1.68, 95% CI = −3.22 to −0.14, P = 0.033, n = 32). Planned comparisons revealed higher PA after exercise compared to rest (B = 2.88, 95% CI = 0.96–4.80, P = 0.003, n = 32), and no differences in PA were found at baseline (B = −0.02, 95% CI = −2.07 to 2.04, P = 0.988, n = 32; Figure 3A). None of these effects were significant for NA (P > 0.122), suggesting a degree of specificity.
Fig. 3.
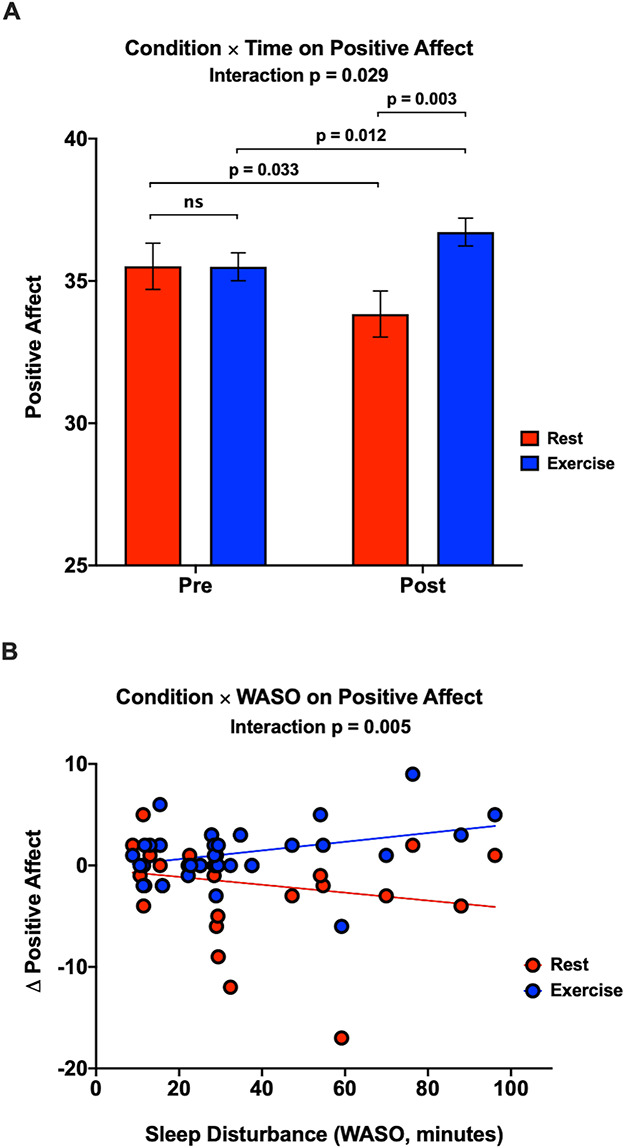
Exercise boosts PA to a greater degree among individuals with greater sleep disturbance. A. Condition × Time interaction for PA. Planned comparisons demonstrated that exercise significantly increased PA, whereas rest significantly decreased PA. Results also revealed higher PA after exercise compared to rest. PA did not differ across intevention arms at baseline. Erros bars represent the standard error. B. Condition × WASO interaction for PA. Individuals with greater sleep disturbance (WASO) exhibited greater exercise-induced improvements in positive affect [(post-exercise  pre-exercise)
pre-exercise)  (post-rest
(post-rest  pre-rest)].
pre-rest)].
The positive impact of exercise is magnified among individuals with greater sleep disturbance
As expected, we found a significant Condition × Sleep interaction, such that individuals with greater sleep disturbance (WASO) showed magnified exercise-induced improvements in PA (B = −0.08, 95% CI = −0.14 to −0.02, P = 0.005, n = 32; Figure 3B). Comparable effects were not evident for sleep duration (TST), P = 0.148.
Acute exercise decreases cingulo-opercular iFC with the hippocampus and cerebellum
Voxelwise analyses indicated that acute exercise significantly decreased iFC between the cingulo-opercular network (AI hubs) and several regions, including the left hippocampus (anterior/posterior; 18, 31, −5) and left cerebellum (subregion VIII; −18, 31, −5), (Figures 4A & 4B). Acute exercise did not significantly affect iFC of the frontoparietal network (not depicted), suggesting that this effect is specific to the cingulo-opercular network.
Fig. 4.
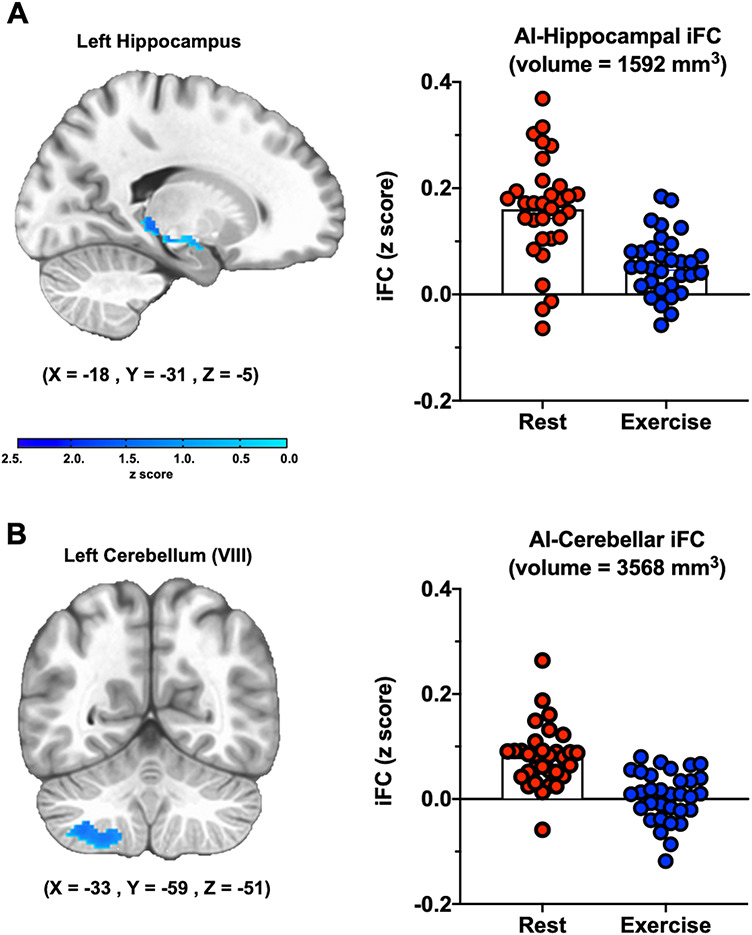
Acute aerobic exercise reduces iFC between the cingulo-opercular network (AI hubs) and the hippocampus and cerebellum. A. Hippocampus. B. Cerebellum. For illustrative purposes, bar graphs depict the mean iFC for each arm of the crossover intervention.
Alterations in cingulo-opercular network iFC mediate the link between sleep disturbance and exercise-induced PA
Paralleling the behavioral results, individuals with greater sleep disturbance (WASO) showed greater exercise-induced decreases in iFC between the cingulo-opercular network (AI hubs) and the left hippocampus (B = −0.0020, 95% CI = −0.0040 to −0.0004, P = 0.018, n = 32). The same pattern was evident, but failed to reach significance, for the left cerebellum (B = −0.00099, 95% CI = −0.00198 to 0.00001, P = 0.052, n = 32). Thus, the AI-cerebellum functional connection was excluded from subsequent analyses. Individuals evincing greater exercise-induced decreases in hippocampal iFC also showed greater exercise-induced increases in PA (B = −16.58, 95% CI = −29.69 to −3.46, P = 0.015, n = 32). Moreover, those with greater sleep disturbance (WASO) showed larger exercise-induced improvements in PA (B = 0.067, 95% CI = 0.004–0.129, P = 0.038, n = 32). A nonparametric bootstrapping procedure revealed that exercise-induced decreases in iFC between the cingulo-opercular network (AI hubs) and the left hippocampus significantly mediated the association between sleep disturbance (WASO) and exercise-induced increases in PA (B = 0.025, 95% CI = 0.004–0.070, P = 0.018, n = 32) (Figure 5).
Fig. 5.
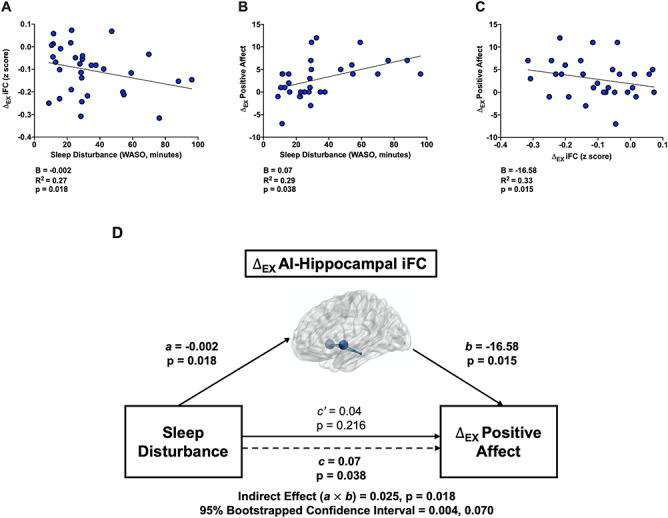
Exercise-induced changes ( EX) in iFC between the cingulo-opercular network (AI hubs) and left hippocampus mediate
EX) in iFC between the cingulo-opercular network (AI hubs) and left hippocampus mediate  EX in positive affect (PA) among elders with disturbed sleep (WASO). A. Relations between objective sleep disturbance and
EX in positive affect (PA) among elders with disturbed sleep (WASO). A. Relations between objective sleep disturbance and 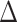 EX in iFC (post-exercise
EX in iFC (post-exercise  post-rest) between the cingulo-opercular network (AI hubs) and left hippocampus (path a in panel D). B. Relations between
post-rest) between the cingulo-opercular network (AI hubs) and left hippocampus (path a in panel D). B. Relations between 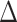 EX in hippocampal iFC and
EX in hippocampal iFC and 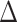 EX in PA [(post-exercise
EX in PA [(post-exercise  pre-exercise)
pre-exercise)  (post-rest
(post-rest  pre-rest)] (path b in panel D). C. Relations between sleep disturbance and
pre-rest)] (path b in panel D). C. Relations between sleep disturbance and 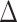 EX in PA (path c in panel D). D. Statistical mediation framework. A non-parametric bootstrap analysis confirmed a significant mediation effect, indicating that
EX in PA (path c in panel D). D. Statistical mediation framework. A non-parametric bootstrap analysis confirmed a significant mediation effect, indicating that 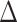 EX in AI-left hippocampal iFC partially explain relations between objective sleep disturbance and
EX in AI-left hippocampal iFC partially explain relations between objective sleep disturbance and 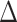 EX in PA (path c’ and the significant indirect effect of a
EX in PA (path c’ and the significant indirect effect of a 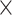 b). Path.
b). Path.
Discussion
Older adults are at increased risk for exacerbation of chronic medical conditions through poor emotional well-being, which can be further compounded by poor sleep. Exercise represents a viable intervention, yet there is little known about how the affective changes elicited by a single dose of exercise interact with sleep quality, or the neural networks that may drive these effects. In this study of physically active, cognitively normal older adults, 30 min of moderate aerobic exercise increased PA and reduced iFC between the cingulo-opercular network (AI hubs) and several regions, including the hippocampus and cerebellum. Both effects were magnified for older adults with objectively greater sleep disturbance (WASO). Mediation analyses demonstrated that exercise-induced changes in cingulo-opercular (AI-hippocampal) iFC statistically explained relations between sleep disturbance (WASO) and exercise-induced increases in PA. In contrast, significant effects were not evident for NA, total sleep duration (TST), or the frontoparietal network, suggesting a degree of specificity. Taken together, these findings provide compelling new evidence that aerobic exercise acutely enhances mood, that it does so by altering connectivity between the AI—a key hub in the cingulo-opercular (or ‘salience’) network—and the hippocampus, and that lower sleep quality is a stronger predictor of these effects among older adults.
Our findings linking cingulo-opercular network (AI-hippocampal) iFC with PA are supported by studies that examined functional connectivity in the context of mood disorders, and psychological stress (see Menon (2011) for an extensive review). One such study (Sheline et al., 2010) examined the connectivity patterns of three large-scale brain networks (the cognitive control, default mode, and affective networks) in 18 young adults with major depression, compared to 17 age-matched controls. Individuals with depression exhibited increased connectivity in the bilateral dorsal medial prefrontal cortex in all three networks. This brain region, the dorsal nexus, overlaps with the ACC, which along with the AI, is a primary hub of the cingulo-opercular network (Menon and Uddin, 2010). These findings suggest a central role for this region in the etiology of depression, and further propose that iFC reductions in regions related to the AI may help mitigate depressive symptoms. A second study (van Marle et al., 2010) investigated functional connectivity patterns immediately after acute psychological stress, compared to a no-stress control, in 26 young women. Results revealed that those exposed to psychological stress exhibited greater connectivity between prominent corticolimbic regions of the cingulo-opercular network, including the amygdala, MCC, AI and locus coeruleus, compared to controls, suggesting that cingulo-opercular iFC may represent a measurable target for interventions aiming to reduce psychological distress. Our findings are internally consistent with the interpretation that decreased cingulo-opercular network iFC after exercise is a driver of the exercise-related increase in subjectively rated PA, particularly among those who experience more disrupted sleep (greater WASO), in whom mood disorders are known to be more prevalent (Krystal, 2012). Additionally, while another study found that exercise altered frontoparietal network iFC (Prehn et al., 2019), we identified no such effect following an acute bout of exercise (vs. rest), which may reflect differences in experimental designs. A key challenge for future research will be to conduct larger-scale studies that are conducive to parametric analyses—including individuals of varying fitness levels (e.g., untrained vs trained; normal weight vs. obese), exercise intensity, exercise mode, and clinical status (e.g., individuals diagnosed with or at risk of developing clinically significant depressive symptoms) to determine the reproducibility of such effects (Poldrack et al., 2017).
Sleep deprivation and restriction are linked to irritability and lower PA (Kamphuis et al., 2012; Goldstein et al., 2013; Minkel et al., 2014; Pires et al., 2016; Ben Simon et al., 2020), and recent findings suggest that sleep loss and sleep disruption consistently reduce PA to a greater degree than NA (Finan et al., 2015 2017; Ben Simon et al., 2020). Such discrepancies may relate, in part, to sleep’s role in mood disorders, particularly depression, in which anhedonia (or the inability to experience PA) is a hallmark feature (Ben Simon et al., 2020). Our participants were not sleep deprived, yet we found that greater sleep disruption (WASO) predicted the extent to which exercise increased PA. Considering the strong link between poor sleep and mood disorders (Peterson and Benca, 2006; Baglioni et al., 2011), it is possible that individuals with greater sleep disturbances may exhibit an affective (behavioral and neuronal) phenotype similar to those with depression (Triantafillou et al., 2019; Ben Simon et al., 2020). Further, numerous studies have demonstrated the mood-enhancing effects of aerobic exercise in patients with depression (Blumenthal et al., 1999 2007; Brush et al., 2020), but conflicting data suggest that such effects may be dampened and perhaps inconsequential in people with normal (non-depressed) moods (Lennox et al., 1990). If individuals with sleep disruptions and mood disorders do, in fact, share similar affective characteristics, it would help explain why exercise might preferentially improve emotional well-being more among those with poor sleep.
Our findings were particularly strong in terms of the reduced connectivity between the AI and left hippocampus. Exercise is known to strongly impact hippocampal function (Pereira et al., 2007; Erickson et al., 2011; Won et al., 2019b). In our previously published work, acute exercise increased the activation of the hippocampus during semantic memory retrieval (Won et al., 2019b). Together, these findings suggest that acute exercise may tune hippocampal neurons for adaptive responses that promote positive affective experience and memory retrieval. Future studies should evaluate whether acute exercise impacts other connections within the cingulo-opercular network, namely those connected to the dorsal ACC.
Our findings concerning the effects of acute exercise on PA are consistent with a large body of prior research (Ekkekakis and Petruzzello, 1999; Reed and Ones, 2006). Previous studies have shown that exercise performed at low-to-moderate intensity (Berger and Motl, 2000; Ekkekakis et al., 2000) for at least 10 min (Jakicic et al., 1995) increases PA and that PA can remain elevated for up to 30 min after exercise (Steptoe and Bolton, 1988). To date, very few studies have examined the combined effects of sleep and exercise on acute PA. Results from one such investigation suggest that aerobic exercise training reduces symptoms of post-traumatic stress disorder among veterans reporting poor baseline sleep, compared to those reporting good baseline sleep (Babson et al., 2015). Notably, the current study is broadly consistent with these observations and specifically implicates decreased AI-hippocampal connectivity as a possible mechanism.
Potential mechanisms
The mood-enhancing effects of acute exercise may be linked to the upregulation of several signaling pathways, including those stimulated by stress hormones (Kirschbaum and Hellhammer, 1989; Budde et al., 2015), endocannabinoids (Dietrich and McDaniel, 2004) and growth factors (Cappon et al., 1994). Moderate-intensity exercise activates the hypothalamic–pituitary–adrenal (HPA) axis by stimulating the secretion of corticotropin-releasing hormone (CRH) from the hypothalamus, which leads to the release of the adrenocorticotropic hormone from the anterior pituitary gland, and the secretion of cortisol from the adrenal cortex (Mastorakos and Pavlatou, 2005). As an integral factor in the fight or flight response, cortisol acts not only to liberate stored energy by stimulating gluconeogenesis and glycogenesis but also exerts feedback inhibition on the hypothalamus to stop the secretion of CRH (Goncharova et al., 2019). Additionally, cortisol crosses the blood–brain barrier and binds to receptors located on several cingulo-opercular network-related brain regions (i.e., AI, hippocampus), where it may help regulate affect and emotion processing (Thomason et al., 2011; Banks, 2012). However, cortisol exerts differential effects on brain regions depending on the receptor to which it binds (Lupien et al., 2007). Animal studies have shown that voluntary exercise reduces mineralocorticoid receptor binding, and increases glucocorticoid receptor binding in the hippocampus, facilitating hippocampal modulation of the stress response (Stranahan et al., 2008). Nonetheless, despite receptor affinity, short-term release of cortisol is likely beneficial, while prolonged release of cortisol can be deleterious and inhibitive of the stress response (Lupien et al., 2007). Additional experimental studies are necessary to understand how the HPA axis may mediate exercise-induced improvements in PA. Several lines of research suggest that exercise—a predictable, controllable stressor—may modulate HPA activity by enhancing cellular function and promoting hippocampal negative feedback (on the HPA axis), thereby shutting down the release of cortisol (Stranahan et al., 2008). As such, the effects of exercise could potentially normalize, or protect against, the HPA axis hyperactivation observed in both poor sleep and mood disorders (Watson and Mackin, 2006).
Other animal studies have suggested that exercise may improve mood states by increasing galanin (a neuropeptide implicated in the circadian regulation of sleep/wake patterns and feeding) and suppressing the stress-related neurotransmitter norepinephrine (Sciolino and Holmes, 2012).
Similarly, growth factors, including brain-derived neurotrophic factor, insulin-like growth factor-1 and vascular endothelial growth factor are upregulated in response to acute exercise and have been linked to adult hippocampal neurogenesis (Ding et al., 2006; Cotman et al., 2007). Although widely known for their effects on cognition (Cotman et al., 2007), neurotrophic factors bind to receptors on the hippocampus and prefrontal cortex, and have been shown to improve mood and depressive symptoms (Castrén and Kojima, 2017). Similarly, in terms of network iFC, both sleep loss and affective disorders have been associated with elevated cingulo-opercular network iFC (Sheline et al., 2010; van Marle et al., 2010; Simon et al., 2015; Ben Simon et al., 2020), and prior reports have demonstrated the salutary effects of exercise on network iFC in both healthy and clinical populations (Voss et al., 2010; Flodin et al., 2015; Chirles et al., 2017). Although more research is warranted, it is possible that the acute exercise intervention in the current study improved PA through a normalization of AI-hippocampal iFC.
Future challenges
Our study is not without limitations. MRI scans occurred after both conditions, but were not conducted prior to the exercise or control interventions, precluding us from evaluating ‘passage of time’ effects on functional MRI (fMRI). This represents an important challenge for future research. The use of larger and more diverse samples—including individuals with a greater range of age-related impairments—would enhance the reproducibility and generalizability of such work (Munafò et al., 2017; Fox et al., 2018), as would demographic information about work status and schedule, which has the potential to impact sleep timing and duration. An argument can also be made that sedentary behavior, alone, may reduce PA; however, we used a true ‘no treatment’ control condition, in which the only aspect that differed between the two conditions was pedaling the ergometer. We found no evidence that participants had a negative affective experience during either condition. In fact, valence ratings were significantly greater during seated rest compared to exercise (Table 2). Further evaluations comparing the effects of different types of sedentary conditions (e.g., ‘no treatment’ vs ‘reading’) on affect are warranted. Additionally, future, larger-scale studies with greater statistical power should consider implementing more sophisticated analytic techniques (e.g., moderated mediation), which could help identify the potential causal effects of exercise on PA and determine whether sleep moderates some or all of these pathways. Such strategies would improve our understanding of sleep’s role in the relationship between exercise and PA. Like most fMRI studies, our results are observational and, excepting the impact of exercise on mood and iFC, do not license causal claims. Moreover, while subjective and objective measures of sleep do not always align, both provide important and likely complementary information. Future large-scale studies would afford a critical opportunity to compare the value of objective measures of sleep vs self-reported sleep quality, which has also been linked to affect, well-being and psychosocial risk factors (Steptoe et al., 2008). Another key avenue for future research will be to manipulate sleep (e.g., via acute deprivation) and cingulo-opercular (AI-hippocampal) iFC (e.g., via neurofeedback) to clarify their mechanistic importance.
Conclusions
Compared to sedentary rest, 30 min of aerobic exercise acutely enhanced PA and reduced iFC between the AI hubs of the cingulo-opercular network and the hippocampus. Both effects were amplified among healthy older adults with objectively greater sleep disturbance. Mediation analyses demonstrated that decreased AI-hippocampal iFC following acute exercise accounted for a significant proportion of the greater exercise-induced increase in PA in those with greater sleep disturbance (WASO). These observations underscore the benefits of moderate intensity exercise—a safe and scalable behavioral intervention—and provide new clues about the neural circuitry underlying the interactive effects of sleep and exercise on mood in older age. The use of a randomized crossover design and objective actigraphic measures of sleep quality enhances our confidence in the robustness and translational relevance of these results in later life.
Contributor Information
Alfonso J Alfini, PhD, Department of Mental Health, Johns Hopkins Bloomberg School of Public Health, Baltimore, MD 21205, USA.
Junyeon Won, MA, Department of Kinesiology, University of Maryland School of Public Health, College Park, MD 20742, USA.
Lauren R Weiss, PhD, Department of Kinesiology, University of Maryland School of Public Health, College Park, MD 20742, USA; Neuroscience and Cognitive Science Program, University of Maryland, College Park, MD 20742, USA.
Casandra C Nyhuis, Department of Mental Health, Johns Hopkins Bloomberg School of Public Health, Baltimore, MD 21205, USA.
Alexander J Shackman, PhD, Neuroscience and Cognitive Science Program, University of Maryland, College Park, MD 20742, USA; Department of Psychology, University of Maryland, College Park, MD, USA.
Adam P Spira, PhD, Department of Mental Health, Johns Hopkins Bloomberg School of Public Health, Baltimore, MD 21205, USA; Department of Psychiatry and Behavioral Sciences, Johns Hopkins School of Medicine, Baltimore, MD 21205, USA; Center on Aging and Health, Johns Hopkins School of Medicine and Bloomberg School of Public Health, Baltimore, MD 21205, USA.
J Carson Smith, PhD, Department of Kinesiology, University of Maryland School of Public Health, College Park, MD 20742, USA; Neuroscience and Cognitive Science Program, University of Maryland, College Park, MD 20742, USA.
Author Contributions
J.C.S developed the study idea and overall study protocol. A.J.A. and J.W. managed participant recruitment and data collection. A.J.A, L.R.W. and J.W. collected data. L.R.W. oversaw data management. C.C.N. and A.J.A. scored actigraphy data. A.J.A., J.W. and L.R.W. processed and analyzed imaging data. A.J.A. analyzed psychometric data. A.J.A, J.C.S, A.P.S. and A.J.S. interpreted the data. J.C.S., A.J.A, A.P.S. and A.J.S. collectively developed the analytic strategy. A.J.A. drafted the paper and J.W., L.R.W., C.C.N., A.P.S., A.J.S and J.C.S edited the paper. A.J.A. created the figures. All authors reviewed, revised and approved the final manuscript.
Funding
This work was supported by the National Institutes of Health [AG27668-13] and University of Maryland, College Park. We thank the participants for their time and dedication to this study.
Resource Sharing
Key statistical maps and regions-of-interest have been uploaded to NeuroVault.org.
Conflict of Interest
A.P.S. received an honorarium from Springer Nature Switzerland AG for Guest Editing a Special Issue of Current Sleep Medicine Reports. All other authors have no conflicts of interest.
References
- Alfini A.J., Schrack J.A., Urbanek J.K., et al. (2020). Associations of actigraphic sleep parameters with fatigability in older adults. The Journals of Gerontology: Series A , 75, e95–e102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alfini A.J., Weiss L.R., Leitner B.P., Smith T.J., Hagberg J.M., Smith J.C. (2016). Hippocampal and cerebral blood flow after exercise cessation in master athletes. Frontiers in Aging Neuroscience, 8, doi: 10.3389/fnagi.2016.00184 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Andreescu C., Sheu L.K., Tudorascu D., et al. (2015). Emotion reactivity and regulation in late-life generalized anxiety disorder: functional connectivity at baseline and post-treatment. The American Journal of Geriatric Psychiatry, 23(2), 200–14. doi: 10.1016/j.jagp.2014.05.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arszovszki A., Borhegyi Z., Klausberger T. (2014). Three axonal projection routes of individual pyramidal cells in the ventral CA1 hippocampus. Frontiers in Neuroanatomy, 8, doi: 10.3389/fnana.2014.00053 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Babson K.A., einz A.J., Ramirez G., et al. (2015). The interactive role of exercise and sleep on veteran recovery from symptoms of PTSD. Mental Health and Physical Activity, 8, 15–20. doi: 10.1016/j.mhpa.2014.12.002 [DOI] [Google Scholar]
- Baglioni C., Battagliese G., Feige B., et al. (2011). Insomnia as a predictor of depression: a meta-analytic evaluation of longitudinal epidemiological studies. Journal of Affective Disorders, 135(1), 10–19. doi: 10.1016/j.jad.2011.01.011 [DOI] [PubMed] [Google Scholar]
- Banks W.A. (2012). Brain meets body: the blood-brain barrier as an endocrine interface. Endocrinology, 153(9), 4111–19. doi: 10.1210/en.2012-1435 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baron R.M., Kenny D.A. (1986). The moderator-mediator variable distinction in social psychological research: conceptual, strategic, and statistical considerations. Journal of Personality and Social Psychology, 51(6), 1173–82. doi: 10.1037//0022-3514.51.6.1173 [DOI] [PubMed] [Google Scholar]
- Beall E.B., Lowe M.J. (2014). SimPACE: generating simulated motion corrupted BOLD data with synthetic-navigated acquisition for the development and evaluation of SLOMOCO: a new, highly effective slicewise motion correction. NeuroImage, 101, 21–34. doi: 10.1016/j.neuroimage.2014.06.038 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beck A.T., Steer R.A., Brown G.K. (1996). Beck Depression Inventory-ii (Bdi-ii). San Antonio, TX: Psychological Corporation. [Google Scholar]
- Ben Simon E., Vallat R., Barnes C.M., Walker M.P. (2020). Sleep loss and the socio-emotional brain. Trends in Cognitive Sciences, 24(6), 435–50. doi: 10.1016/j.tics.2020.02.003 [DOI] [PubMed] [Google Scholar]
- Berger B.G., Motl R.W. (2000). Exercise and mood: a selective review and synthesis of research employing the profile of mood states. Journal of Applied Sport Psychology, 12(1), 69–92. doi: 10.1080/10413200008404214 [DOI] [Google Scholar]
- Blumenthal J.A., Babyak M.A., Doraiswamy P.M., et al. (2007). Exercise and pharmacotherapy in the treatment of major depressive disorder. Psychosomatic Medicine, 69(7), 587. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blumenthal J.A., Babyak M.A., Moore K.A., et al. (1999). Effects of exercise training on older patients with major depression. Archives of Internal Medicine, 159(19), 2349–56. [DOI] [PubMed] [Google Scholar]
- Borg G. (1990). Psychophysical scaling with applications in physical work and the perception of exertion. Scandinavian Journal of Work, Environment & Health, 16(Suppl 1), 55–58. [DOI] [PubMed] [Google Scholar]
- Bradley M.M., Lang P.J. (1994). Measuring emotion: the self-assessment manikin and the semantic differential. Journal of Behavior Therapy and Experimental Psychiatry, 25(1), 49–59. doi: 10.1016/0005-7916(94)90063-9 [DOI] [PubMed] [Google Scholar]
- Brush C.J., Hajcak G., Bocchine A.J., et al. (2020). A randomized trial of aerobic exercise for major depression: examining neural indicators of reward and cognitive control as predictors and treatment targets. Psychological Medicine, 1–11. doi: 10.1017/S0033291720002573 [DOI] [PubMed] [Google Scholar]
- Budde H., Machado S., Ribeiro P., Wegner M. (2015). The cortisol response to exercise in young adults. Frontiers in Behavioral Neuroscience, 9, 13. doi: 10.3389/fnbeh.2015.00013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buysse D.J. (2004). Insomnia, depression and aging. Assessing sleep and mood interactions in older adults. Geriatrics, 59(2), 47–51. quiz 52 [PubMed] [Google Scholar]
- Cappon J., Brasel J.A., Mohan S., Cooper D.M. (1994). Effect of brief exercise on circulating insulin-like growth factor I. Journal of Applied Physiology, 76(6), 2490–96. doi: 10.1152/jappl.1994.76.6.2490 [DOI] [PubMed] [Google Scholar]
- Castrén E., Kojima M. (2017). Brain-derived neurotrophic factor in mood disorders and antidepressant treatments. Neurobiology of Disease, 97(Pt B), 119–26. doi: 10.1016/j.nbd.2016.07.010 [DOI] [PubMed] [Google Scholar]
- Chen M.C., Chang C., Glover G.H., Gotlib I.H. (2014). Increased insula coactivation with salience networks in insomnia. Biological Psychology, 97, 1–8. doi: 10.1016/j.biopsycho.2013.12.016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chirles T.J., Reiter K., Weiss L.R., Alfini A.J., Nielson K.A., Smith J.C. (2017). Exercise training and functional connectivity changes in mild cognitive impairment and healthy elders. Journal of Alzheimer’s Disease, 57(3), 845–56. doi: 10.3233/JAD-161151 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Choi E.Y., Yeo B.T., Buckner R.L. (2012). The organization of the human striatum estimated by intrinsic functional connectivity. Journal of Neurophysiology, 108(8), 2242–63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chung F., Abdullah H.R., Liao P. (2016). STOP-Bang questionnaire: a practical approach to screen for obstructive sleep apnea. Chest, 149(3), 631–38. doi: 10.1378/chest.15-0903 [DOI] [PubMed] [Google Scholar]
- Cotman C.W., Berchtold N.C., Christie L.-A. (2007). Exercise builds brain health: key roles of growth factor cascades and inflammation. Trends in Neurosciences, 30(9), 464–72. doi: 10.1016/j.tins.2007.06.011 [DOI] [PubMed] [Google Scholar]
- Cox R.W., Glen D.R. (2013). Nonlinear warping in AFNI. Poster Presented at the 19th Annual Meeting of the Organization for Human Brain Mapping. [Google Scholar]
- Cox R.W. (1996). AFNI: software for analysis and visualization of functional magnetic resonance neuroimages. Computers and Biomedical Research, 29(3), 162–73. doi: 10.1006/cbmr.1996.0014 [DOI] [PubMed] [Google Scholar]
- Dhar A.K., Barton D.A. (2016). Depression and the link with cardiovascular disease. Frontiers in Psychiatry, 7, doi: 10.3389/fpsyt.2016.00033 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dickens C., McGowan L., Clark-Carter D., Creed F. (2002). Depression in rheumatoid arthritis: a systematic review of the literature with meta-analysis. Psychosomatic Medicine, 64(1), 52–60. doi: 10.1097/00006842-200201000-00008 [DOI] [PubMed] [Google Scholar]
- Dietrich A., McDaniel W.F. (2004). Endocannabinoids and exercise. British Journal of Sports Medicine, 38(5), 536. doi: 10.1136/bjsm.2004.011718 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ding Q., Vaynman S., Akhavan M., Ying Z., Gomez-Pinilla F. (2006). Insulin-like growth factor I interfaces with brain-derived neurotrophic factor-mediated synaptic plasticity to modulate aspects of exercise-induced cognitive function. Neuroscience, 140(3), 823–33. doi: 10.1016/j.neuroscience.2006.02.084 [DOI] [PubMed] [Google Scholar]
- Ekkekakis P., Hall E.E., VanLanduyt L.M., Petruzzello S.J. (2000). Walking in (affective) circles: can short walks enhance affect?. Journal of Behavioral Medicine, 23(3), 245–75. [DOI] [PubMed] [Google Scholar]
- Ekkekakis P., Petruzzello S.J. (1999). Acute aerobic exercise and affect—Current status, problems and prospects regarding dose-response. Sports Medicine, 28(5), 337–74. doi: 10.2165/00007256-199928050-00005 [DOI] [PubMed] [Google Scholar]
- Eklund A., Nichols T.E., Knutsson H. (2016). Cluster failure: why fMRI inferences for spatial extent have inflated false-positive rates. Proceedings of the National Academy of Sciences of the United States of America, 113(28), 7900–05. doi: 10.1073/pnas.1602413113 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Erickson K.I., Voss M.W., Prakash R.S., et al. (2011). Exercise training increases size of hippocampus and improves memory. Proceedings of the National Academy of Sciences of the United States of America, 108(7), 3017–22. doi: 10.1073/pnas.1015950108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fang X., Zhang Y., Zhou Y., et al. (2016). Resting-state coupling between core regions within the central-executive and salience networks contributes to working memory performance. Frontiers in Behavioral Neuroscience, 10, doi: 10.3389/fnbeh.2016.00027 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Faubel R., Lopez-Garcia E., Guallar-Castillón P., et al. (2009). Sleep duration and health-related quality of life among older adults: a population-based cohort in Spain. Sleep, 32(8), 1059–68. [PMC free article] [PubMed] [Google Scholar]
- Finan P.H., Quartana P.J., Remeniuk B., et al. (2017). Partial sleep deprivation attenuates the positive affective system: effects across multiple measurement modalities. Sleep, 40, 1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Finan P.H., Quartana P.J., Smith M.T. (2015). The effects of sleep continuity disruption on positive mood and sleep architecture in healthy adults. Sleep, 38(11), 1735–42. doi: 10.5665/sleep.5154 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fischl B. (2012). FreeSurfer. 20 Years of fMRI, 62(2), 774–81. doi: 10.1016/j.neuroimage.2012.01.021 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Flodin P., Martinsen S., Mannerkorpi K., et al. (2015). Normalization of aberrant resting state functional connectivity in fibromyalgia patients following a three month physical exercise therapy. NeuroImage: Clinical, 9, 134–39. doi: 10.1016/j.nicl.2015.08.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Folstein M.F., Robins L.N., Helzer J.E. (1983). The Mini-Mental State Examination. JAMA Psychiatry, 40(7), 812–812. doi: 10.1001/archpsyc.1983.01790060110016 [DOI] [PubMed] [Google Scholar]
- Fox A.S., Lapate R.C., Davidson R.J., Shackman A.J. (2018). The nature of emotion: a research agenda for the twenty-first century. The Nature of Emotion. Fundamental Questions, 2, 81 [Google Scholar]
- Friedrich M.J. (2017). Depression is the leading cause of disability around the world. JAMA, 317(15), 1517–1517. doi: 10.1001/jama.2017.3826 [DOI] [PubMed] [Google Scholar]
- Furman D.J., Hamilton J.P., Gotlib I.H. (2011). Frontostriatal functional connectivity in major depressive disorder. Biology of Mood & Anxiety Disorders, 1(1), 11. doi: 10.1186/2045-5380-1-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldstein A.N., Greer S.M., Saletin J.M., Harvey A.G., Nitschke J.B., Walker M.P. (2013). Tired and apprehensive: anxiety amplifies the impact of sleep loss on aversive brain anticipation. Journal of Neuroscience, 33(26), 10607–15. doi: 10.1523/JNEUROSCI.5578-12.2013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Golkar A., Lonsdorf T.B., Olsson A., et al. (2012). Distinct contributions of the dorsolateral prefrontal and orbitofrontal cortex during emotion regulation. PloS One, 7(11), e48107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goncharova N., Chigarova O., Rudenko N., Oganyan T. (2019). Glucocorticoid negative feedback in regulation of the hypothalamic-pituitary-adrenal axis in rhesus monkeys with various types of adaptive behavior: individual and age-related differences. Frontiers in Endocrinology, 10, doi: 10.3389/fendo.2019.00024 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gordon B.R., McDowell C.P., Hallgren M., Meyer J.D., Lyons M., Herring M.P. (2018). Association of efficacy of resistance exercise training with depressive symptoms: meta-analysis and meta-regression analysis of randomized clinical trials. JAMA Psychiatry, 75(6), 566–76. doi: 10.1001/jamapsychiatry.2018.0572 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jakicic J.M., Wing R.R., Butler B.A., Robertson R.J. (1995). Prescribing exercise in multiple short bouts versus one continuous bout: effects on adherence, cardiorespiratory fitness, and weight loss in overweight women. International Journal of Obesity and Related Metabolic Disorders: Journal of the International Association for the Study of Obesity, 19(12), 893–901. [PubMed] [Google Scholar]
- Kamphuis J., Meerlo P., Koolhaas J.M., Lancel M. (2012). Poor sleep as a potential causal factor in aggression and violence. Sleep Medicine, 13(4), 327–34. doi: 10.1016/j.sleep.2011.12.006 [DOI] [PubMed] [Google Scholar]
- Kennedy B.K., Berger S.L., Brunet A., et al. (2014). Geroscience: linking aging to chronic disease. Cell, 159(4), 709–13. doi: 10.1016/j.cell.2014.10.039 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khan A.R., Wang L., Beg M.F. (2008). FreeSurfer-initiated fully-automated subcortical brain segmentation in MRI using large deformation diffeomorphic metric mapping. NeuroImage, 41(3), 735–46. doi: 10.1016/j.neuroimage.2008.03.024 [DOI] [PMC free article] [PubMed] [Google Scholar]
- King A.C., Oman R.F., Brassington G.S., Bliwise D.L., Haskell W.L. (1997). Moderate-intensity exercise and self-rated quality of sleep in older adults: a randomized controlled trial. JAMA, 277(1), 32–37. doi: 10.1001/jama.1997.03540250040029 [DOI] [PubMed] [Google Scholar]
- Kirschbaum C., Hellhammer D.H. (1989). Salivary cortisol in psychobiological research: an overview. Neuropsychobiology, 22(3), 150–69. doi: 10.1159/000118611 [DOI] [PubMed] [Google Scholar]
- Kline C.E., Crowley E.P., Ewing G.B., et al. (2011). The effect of exercise training on obstructive sleep apnea and sleep quality: a randomized controlled trial. Sleep, 34(12), 1631–40. doi: 10.5665/sleep.1422 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koch S.B.J., van Zuiden M., Nawijn L., Frijling J.L., Veltman D.J., Olff M. (2016). Aberrant resting-state brain activity in posttraumatic stress disorder: a meta-analysis and systematic review. Depression and Anxiety, 33(7), 592–605. doi: 10.1002/da.22478 [DOI] [PubMed] [Google Scholar]
- Krause A.J., Ben Simon E., Mander B.A., et al. (2017). The sleep-deprived human brain. Nature Reviews. Neuroscience, 18(7), 404–18. doi: 10.1038/nrn.2017.55 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krystal A.D. (2012). Psychiatric disorders and sleep. Neurologic Clinics, 30(4), 1389–413. doi: 10.1016/j.ncl.2012.08.018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lemola S., Ledermann T., Friedman E.M. (2013). Variability of sleep duration is related to subjective sleep quality and subjective well-being: an actigraphy study. Plos One, 8(8), e71292. doi: 10.1371/journal.pone.0071292 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lennox S.S., Bedell J.R., Stone A.A. (1990). The effect of exercise on normal mood. Journal of Psychosomatic Research, 34(6), 629–36. doi: 10.1016/0022-3999(90)90106-E [DOI] [PubMed] [Google Scholar]
- Li J., Vitiello M.V., Gooneratne N.S. (2018). Sleep in normal aging. Sleep Medicine Clinics, 13(1), 1–11. doi: 10.1016/j.jsmc.2017.09.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lupien S.J., Maheu F., Tu M., Fiocco A., Schramek T.E. (2007). The effects of stress and stress hormones on human cognition: implications for the field of brain and cognition. Brain and Cognition, 65(3), 209–37. doi: 10.1016/j.bandc.2007.02.007 [DOI] [PubMed] [Google Scholar]
- Mander B.A., Winer J.R., Walker M.P. (2017). Sleep and human aging. Neuron, 94(1), 19–36. doi: 10.1016/j.neuron.2017.02.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mastorakos G., Pavlatou M. (2005). Exercise as a stress model and the interplay between the hypothalamus-pituitary-adrenal and the hypothalamus-pituitary-thyroid axes. Hormone and Metabolic Research = Hormon Und Stoffwechselforschung = Hormones Et Metabolisme, 37(9), 577–84. doi: 10.1055/s-2005-870426 [DOI] [PubMed] [Google Scholar]
- Menon V. (2015). Salience Network In A. W. Toga (Ed.), Brain Mapping: An encyclopedic reference (Vol.2, pp. 597–611). London: Academic Press: Elsevier. [Google Scholar]
- Menon V. (2011). Large-scale brain networks and psychopathology: a unifying triple network model. Trends in Cognitive Sciences, 15(10), 483–506. [DOI] [PubMed] [Google Scholar]
- Menon V., Uddin L.Q. (2010). Saliency, switching, attention and control: a network model of insula function. Brain Structure & Function, 214(5), 655–67. doi: 10.1007/s00429-010-0262-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mezuk B., Eaton W.W., Albrecht S., Golden S.H. (2008). Depression and type 2 diabetes over the lifespan: a meta-analysis. Diabetes Care, 31(12), 2383–90. doi: 10.2337/dc08-0985 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Minkel J., Moreta M., Muto J., et al. (2014). Sleep deprivation potentiates HPA axis stress reactivity in healthy adults. Health Psychology: Official Journal of the Division of Health Psychology, American Psychological Association, 33(11), 1430–34. doi: 10.1037/a0034219 [DOI] [PubMed] [Google Scholar]
- Morres I.D., Hatzigeorgiadis A., Stathi A., et al. (2019). Aerobic exercise for adult patients with major depressive disorder in mental health services: a systematic review and meta-analysis. Depression and Anxiety, 36(1), 39–53. doi: 10.1002/da.22842 [DOI] [PubMed] [Google Scholar]
- Munafò M.R., Nosek B.A., Bishop D.V., et al. (2017). A manifesto for reproducible science. Nature Human Behaviour, 1(1), 1–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Newgard C.B., Sharpless N.E. (2013). Coming of age: molecular drivers of aging and therapeutic opportunities. The Journal of Clinical Investigation, 123(3), 946–50. doi: 10.1172/JCI68833 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oldfield R.C. (1971). The assessment and analysis of handedness: the Edinburgh inventory. Neuropsychologia, 9(1), 97–113. doi: 10.1016/0028-3932(71)90067-4 [DOI] [PubMed] [Google Scholar]
- Ownby R.L., Crocco E., Acevedo A., John V., Loewenstein D. (2006). Depression and risk for Alzheimer disease. Archives of General Psychiatry, 63(5), 530–38. doi: 10.1001/archpsyc.63.5.530 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pereira A.C., Huddleston D.E., Brickman A.M., et al. (2007). An in vivo correlate of exercise-induced neurogenesis in the adult dentate gyrus. Proceedings of the National Academy of Sciences of the United States of America, 104(13), 5638–43. doi: 10.1073/pnas.0611721104 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peterson M.J., Benca R.M. (2006). Sleep in mood disorders. Psychiatric Clinics of North America, 29(4), 1009–32. doi: 10.1016/j.psc.2006.09.003 [DOI] [PubMed] [Google Scholar]
- Pires G.N., Bezerra A.G., Tufik S., Andersen M.L. (2016). Effects of acute sleep deprivation on state anxiety levels: a systematic review and meta-analysis. Sleep Medicine, 24, 109–18. doi: 10.1016/j.sleep.2016.07.019 [DOI] [PubMed] [Google Scholar]
- Poldrack R.A., Baker C.I., Durnez J., et al. (2017). Scanning the horizon: towards transparent and reproducible neuroimaging research. Nature Reviews. Neuroscience, 18(2), 115–26. doi: 10.1038/nrn.2016.167 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prehn K., Lesemann A., Krey G., et al. (2019). Using resting-state fMRI to assess the effect of aerobic exercise on functional connectivity of the DLPFC in older overweight adults. Brain and Cognition, 131, 34–44. doi: 10.1016/j.bandc.2017.08.006 [DOI] [PubMed] [Google Scholar]
- Rajab A.S., Crane D.E., Middleton L.E., Robertson A.D., Hampson M., MacIntosh B.J. (2014). A single session of exercise increases connectivity in sensorimotor-related brain networks: a resting-state fMRI study in young healthy adults. Frontiers in Human Neuroscience, 8, 625. doi: 10.3389/fnhum.2014.00625 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reed J., Ones D.S. (2006). The effect of acute aerobic exercise on positive activated affect: a meta-analysis. Psychology of Sport and Exercise, 7(5), 477–514. doi: 10.1016/j.psychsport.2005.11.003 [DOI] [Google Scholar]
- Rhyner K.T., Watts A. (2016). Exercise and depressive symptoms in older adults: a systematic meta-analytic review. Journal of Aging and Physical Activity, 24(2), 234–46. doi: 10.1123/japa.2015-0146 [DOI] [PubMed] [Google Scholar]
- Rodda J., Walker Z., Carter J. (2011). Depression in older adults. BMJ, 343, d5219. doi: 10.1136/bmj.d5219 [DOI] [PubMed] [Google Scholar]
- Santos R.V.T., Viana V.A.R., Boscolo R.A., et al. (2012). Moderate exercise training modulates cytokine profile and sleep in elderly people. Cytokine, 60(3), 731–35. doi: 10.1016/j.cyto.2012.07.028 [DOI] [PubMed] [Google Scholar]
- Sciolino N.R., Holmes P.V. (2012). Exercise offers anxiolytic potential: a role for stress and brain noradrenergic-galaninergic mechanisms. Neuroscience and Biobehavioral Reviews, 36(9), 1965–84. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sheline Y.I., Price J.L., Yan Z., Mintun M.A. (2010). Resting-state functional MRI in depression unmasks increased connectivity between networks via the dorsal nexus. Proceedings of the National Academy of Sciences of the United States of America, 107(24), 11020–25. doi: 10.1073/pnas.1000446107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Šidák Z. (1967). Rectangular confidence regions for the means of multivariate normal distributions. Journal of the American Statistical Association, 62(318), 626–33. [Google Scholar]
- Simon E.B., Oren N., Sharon H., et al. (2015). Losing neutrality: the neural basis of impaired emotional control without sleep. Journal of Neuroscience, 35(38), 13194–205. doi: 10.1523/JNEUROSCI.1314-15.2015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simon E.B., Walker M.P. (2018). Sleep loss causes social withdrawal and loneliness. Nature Communications, 9(1), 1–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spiegel D., Giese-Davis J. (2003). Depression and cancer: mechanisms and disease progression. Biological Psychiatry, 54(3), 269–82. doi: 10.1016/s0006-3223(03)00566-3 [DOI] [PubMed] [Google Scholar]
- Spielberger C.D., Gorsuch R.L., Lushene R.E., Vagg P.R., Jacobs G.A. (1970). State-trait Anxiety Inventory. Palo Alto. CA: consulting psychologists press. [Google Scholar]
- Spira A.P., Stone K., Beaudreau S.A., Ancoli-Israel S., Yaffe K. (2009). Anxiety symptoms and objectively measured sleep quality in older women. The American Journal of Geriatric Psychiatry: Official Journal of the American Association for Geriatric Psychiatry, 17(2), 136–43. doi: 10.1097/JGP.0b013e3181871345 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Steptoe A., Bolton J. (1988). The short-term influence of high and low intensity physical exercise on mood. Psychology & Health, 2(2), 91–106. doi: 10.1080/08870448808400346 [DOI] [Google Scholar]
- Steptoe A., O’Donnell K., Marmot M., Wardle J. (2008). Positive affect, psychological well-being, and good sleep. Journal of Psychosomatic Research, 64(4), 409–15. doi: 10.1016/j.jpsychores.2007.11.008 [DOI] [PubMed] [Google Scholar]
- Stranahan A.M., Lee K., Mattson M.P. (2008). Central mechanisms of HPA axis regulation by voluntary exercise. Neuromolecular Medicine, 10(2), 118–27. doi: 10.1007/s12017-008-8027-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tazawa Y., Wada M., Mitsukura Y., et al. (2019). Actigraphy for evaluation of mood disorders: a systematic review and meta-analysis. Journal of Affective Disorders, 253, 257–69. doi: 10.1016/j.jad.2019.04.087 [DOI] [PubMed] [Google Scholar]
- Thomason M.E., Hamilton J.P., Gotlib I.H. (2011). Stress-induced activation of the HPA axis predicts connectivity between subgenual cingulate and salience network during rest in adolescents. Journal of Child Psychology and Psychiatry, and Allied Disciplines, 52(10), 1026–34. doi: 10.1111/j.1469-7610.2011.02422.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tingley D., Yamamoto T., Hirose K., Keele L., Imai K. (2014). mediation: R package for causal mediation analysis. UCLA Statistics/American Statistical Association. https://dspace.mit.edu/handle/1721.1/91154
- Triantafillou S., Saeb S., Lattie E.G. Mohr D.C., Kording K.P. (2019). Relationship between sleep quality and mood: ecological momentary assessment study. JMIR Mental Health, 6, 3. doi: 10.2196/12613 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tukey J.W. (1977). Exploratory data analysis In: Addison-Wesley Exploratory Data Analysis. Frederick Mosteller; Reading, MA: 27–55. [Google Scholar]
- van Marle H.J.F., Hermans E.J., Qin S., Fernández G. (2010). Enhanced resting-state connectivity of amygdala in the immediate aftermath of acute psychological stress. NeuroImage, 53(1), 348–54. doi: 10.1016/j.neuroimage.2010.05.070 [DOI] [PubMed] [Google Scholar]
- Vincent J.L., Kahn I., Snyder A.Z., Raichle M.E., Buckner R.L. (2008). Evidence for a frontoparietal control system revealed by intrinsic functional connectivity. Journal of Neurophysiology, 100(6), 3328–42. doi: 10.1152/jn.90355.2008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Voss M.W., Prakash R.S., Erickson K.I., et al. (2010). Plasticity of brain networks in a randomized intervention trial of exercise training in older adults. Frontiers in Aging Neuroscience, 2, doi: 10.3389/fnagi.2010.00032 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wahl D., Solon-Biet S.M., Cogger V.C., et al. (2019). Aging lifestyle and dementia. Neurobiology of Disease, 130, 104481. doi: 10.1016/j.nbd.2019.104481 [DOI] [PubMed] [Google Scholar]
- Watson D., Clark L.A., Tellegen A. (1988). Development and validation of brief measures of positive and negative affect: the PANAS scales. Journal of Personality and Social Psychology, 54(6), 1063–70. doi: 10.1037//0022-3514.54.6.1063 [DOI] [PubMed] [Google Scholar]
- Watson S., Mackin P. (2006). HPA axis function in mood disorders. Mood Disorders 2, 5(5), 166–70. doi: 10.1383/psyt.2006.5.5.166 [DOI] [Google Scholar]
- Won J., Alfini A.J., Weiss L.R., et al. (2019a). Brain activation during executive control after acute exercise in older adults. International Journal of Psychophysiology, 146, 240–48. doi: 10.1016/j.ijpsycho.2019.10.002 [DOI] [PubMed] [Google Scholar]
- Won J., Alfini A.J., Weiss L.R., et al. (2019b). Semantic memory activation after acute exercise in healthy older adults. Journal of the International Neuropsychological Society: JINS, 1–12. doi: 10.1017/S1355617719000171 [DOI] [PubMed] [Google Scholar]
- Wrzus C., Wagner G.G., Riediger M. (2014). Feeling good when sleeping in? Day-to-day associations between sleep duration and affective well-being differ from youth to old age. Emotion (Washington, D.C.), 14(3), 624–28. doi: 10.1037/a0035349 [DOI] [PubMed] [Google Scholar]
- Yeo B.T., Krienen F.M., Sepulcre J., et al. (2011), The organization of the human cerebral cortex estimated by intrinsic functional connectivity. Journal of Neurophysiology, 106, 1125–1165. [DOI] [PMC free article] [PubMed] [Google Scholar]