

Abstract
The severe acute respiratory syndrome caused by the new coronavirus (SARS-CoV-2), termed COVID-19, has been highlighted as the most important infectious disease of our time, without a vaccine and treatment available until this moment, with a big impact on health systems worldwide, and with high mortality rates associated with respiratory viral disease. The medical and scientific communities have also been confronted by an urgent need to better understand the mechanism of host-virus interaction aimed at developing therapies and vaccines. Since this viral disease can trigger a strong innate immune response, causing severe damage to the pulmonary tract, immunotherapies have also been explored as a means to verify the immunomodulatory effect and improve clinical outcomes, whilst the comprehensive COVID-19 immunology still remains under investigation. In this review, both cellular and molecular immunopathology as well as hemostatic disorders induced by SARS-CoV-2 are summarized. The immunotherapeutic approaches based on the most recent clinical and nonclinical studies, emphasizing their effects for the treatment of COVID-19, are also addressed. The information presented elucidates helpful insights aiming at filling the knowledge gaps around promising immunotherapies that attempt to control the dysfunction of host factors during the course of this infectious viral disease.
1. Introduction, Epidemiology, and Challenges to Near-Future Breakthroughs
In December 2019 in Wuhan, China, the first cases of a serious respiratory disease detected so far without etiology were reported, named by the World Health Organization (WHO) as “new corona virus 2019” and later defined as Coronavirus for Severe Acute Respiratory Syndrome 2 (SARS-CoV-2) causing coronavirus disease, termed as COVID-19 [1, 2].
The identification of the first patients reported with COVID-19 also identified a common history associated with the local market for seafood in Huanan, where wild animals are traded. The genomic characterization of the new virus identified a close similarity to two bat coronaviruses (SARS-CoV and pangolin coronavirus) linking the origin of SARS-CoV-2 with the wholesale market and indicating its zoonotic origin [1, 3].
SARS-CoV-2, responsible for the current pandemic declared on March, 11th by the WHO, has been overwhelming on a global scale, presenting high transmission rates with a large number of deaths, bringing severe effects to health services and economies worldwide. Until July, 20th, WHO reported 14,348,858 confirmed cases of COVID-19 and 603,691 deaths worldwide, reaching all six continents [4].
The SARS-CoV-2 virus belongs to lineage B of the genera β-coronavirus, belonging to the Coronaviridae subfamily. Other viruses of this Coronaviridae family such as HCoV-OC43, HCoV-229E, HCoV-HKU1, and HCoV-NL63, responsible for causing around 15% of common colds [5], are already known, in addition to the Severe Respiratory Syndrome Virus (SARS) and Middle East Respiratory Syndrome-related Coronavirus (MERS) responsible for causing the epidemics in 2002-2004 and 2012, respectively. Of these, SARS, now called SARS-CoV-1, belongs to the same lineage B and, therefore, presents greater epidemiological similarity to SARS-CoV-2. For this reason, it is being widely used in current studies, in an attempt to gain a better knowledge and understanding of this disease and also in the search for treatment and vaccine options for COVID-19 [6–8].
COVID-19 is a disease that may remain asymptomatic, or may present light symptoms, evolving to a more severe disease and can eventually lead to death. The role that asymptomatic individuals play in the spread of SARS-CoV-2 is not well described. Many studies present asymptomatic cases as probable sources of infection, playing an important role in the spread of these viruses [9]. Research estimates that these cases can represent 60% of all infections [10]. However, further studies should be carried out to assess both the amount and infectivity of viral load in asymptomatic individuals to better understand the course of the disease and the necessary methods to fight it [11].
The great challenge of this disease is the lack of knowledge about the SARS-CoV-2 virus, its adaptation, and its effects on the human body, especially as there is no effective drug, vaccine, or treatment available against COVID-19 until this date (July 2020). Therefore, intervention measures to contain the pandemic are limited to social behavioral interventions, such as the use of masks, social distancing, self-isolation, quarantine, and even blocking entire territories and communities, to contain or, at least, mitigate the burden of the ongoing pandemic [12].
The search for a vaccine or an effective treatment is evading laboratories around the world, and several articles have been published daily to share the knowledge obtained. This public health issue is affecting several sectors, mainly the social and economic sectors, causing governmental management to opt for a shorter period of isolation measures, and proceeding to a gradual reopening of the economy, returning to a “new normal.” This restart of activity is believed to cause more outbreak waves, due to increased contact rates [7].
So far, during the onset of COVID-19 symptoms, it has been known that a dysfunction of the immune response is part of the virus-host interaction process and can lead to disease severity and poor outcomes [13–16]. Protective immunity after the infection is not guaranteed. Despite the cross-reactivity with other human common cold coronaviruses in those with previous contact, protection against SARS-CoV-2 infection cannot be confirmed [17]. Furthermore, there is little information regarding immunity [1, 6], raising questions that still have no answers, such as which pathway in the immune response can be the key to attempt to control the immune dysfunction? How can immunotherapy be effective in all subjects affected?
After seven months from the first confirmed case of COVID-19, the knowledge about immune events during SARS-CoV-2 infection remains incomplete, and solutions to the main questions are urgently needed. In this review, we aim to gather information on the main immune responses described in the literature, as well as to offer insights into the promising immunotherapies that can be helpful for the scientific community thus assisting in the delivery of solutions for the urgently needed near-future breakthroughs.
2. Cellular Immune Response during COVID-19: What Is Known So Far
2.1. Humoral Response by B Cells
There is a great discussion regarding the adaptive immune responses to SARS-CoV-2, whether these are protective or pathogenic, or whether both scenarios are possible. The management and knowledge of immune response composition, kinetics, and magnitude can determine the real protection against this pathogen [17]. In this context, the search for correlates of protection biomarkers and reliable immunological parameters has become the key to eliminating this disease [18].
Knowledge of the nature and extent of B cell memory response becomes of great importance, given that the protection from reinfection has directed medical and social initiatives, including the search for vaccine strategies and the return to regular activities. Studies from countries that were in the epicenter of the pandemic showed that, despite a large number of identified cases, the percentage of people who developed antibodies to SARS-CoV-2 is less than expected, like 19.9% in New York City, 17.5% in London, and 11.3% in Madrid [19].
The B cell responses in COVID-19 occur concomitantly with T cell responses, around 1 week after the onset of symptoms. In general, the humoral response to SARS-CoV-2 is more evident between 10 and 25 days [18]. Patients infected with SARS-CoV-2 present the highest levels of IgM antibodies between days 11 and 25 and IgG antibodies between 14 and 25 days after infection [20–22].
CD4+ helper T cells are also known to play an important role in inducing antibody production by B cells during viral infections. During infection, follicular helper T cells (Tfh) are important and crucial for initiating the antibody response at the germinal center for affinity maturation and humoral memory response [23, 24]. In COVID-19, a drop on T cell numbers have been described in the acute phase and even in mild cases, without antibody production or diminishing of IgG antibody detection in the recovery phase [25, 26]. The reduction of virus-specific antibody production could be explained by any failure on the cell differentiation caused by SARS-CoV-2, mainly for CD4+ T cells on the germinative center, where autopsy studies showed lymphocyte depletion in the spleen and lymph nodes [17, 27, 28].
The remaining question is whether the generated antibodies confer long-lasting immunity. In fact, it seems that a subset of patients may not develop long-lasting antibodies to SARS-CoV-2 [29]. Due to the pandemic being a recent event, it is not possible to obtain this response yet; however, it can be predicted based on what is learned from the study of other human coronaviruses [30]. In general, studies have shown that the antibody response among coronaviruses is rarely detected in the acute phase of the disease (until the 7th day). The median time of detection was similar across different antibodies for SARS-CoV-1 (12 days) and SARS-CoV-2 (11 days), but longer for MERS-CoV (16 days) [8].
The comparisons are greater with SARS-CoV-1, which belongs to the same strain of beta-coronavirus [30]. Since 2002, studies have been carried out in order to better understand the activity of this virus. A three-year follow-up study shows that IgM seroconversion was related to a peak on the 15th day, and the performance of IgG and neutralizing antibodies against SARS-CoV-1 peaked at month 4 for both, decreasing over time and becoming almost undetectable in up to 25% of individuals after 36 months [31]. Analysis of IgG memory cells specific for SARS-CoV-1 protein S indicates a progressive cell decrease of ~90% from 2 to 8 months after infection [32]. A follow-up study of 34 healthcare professionals infected with SARS-CoV-1 also shows that virus-specific IgG decreased after several years; however, after 12 years of infection, the authors observed specific detectable IgG [33]. These and other studies of human coronaviruses demonstrate a tendency of antibody response against these viruses to decrease over time. Some studies and mathematical models point to a short duration of immunity after infection by SARS-CoV-2. It was observed that IgG and neutralizing antibody levels in convalescent patients decrease within 2 to 3 months after infection. In another study carried out with 8 convalescent patients, 50% showed a decrease of neutralizing antibody levels in approximately 6 to 7 weeks after disease onset [26]. Further studies will be needed overtime to determine the degree of protection to SARS-CoV-2.
Although many discussions about cross-immunity are taking place, they are still at odds. Previous studies point to a cross-reaction between viruses of the coronavirus family, but reactivity between endemic HCoVs, SARS-CoV-1, and MERS viruses was minimal. Evaluation of plasma from patients with COVID-19 showed a cross-reaction with the S and N SARS-CoV-1 proteins and the S protein of MERS-CoV, but no cross-reaction was observed for the receptor-binding domain (RBD) of these viruses. In addition, it was observed that the plasma of patients with COVID-19 did not neutralize SARS-CoV-1 or MERS-CoV, suggesting that antibody-mediated neutralization is virus-specific and is probably driven by the binding of epitopes within the RBD [34]. Another study of 149 convalescent COVID-19 patients by Robbiani et al., demonstrated that despite the low activity of neutralizing antibodies in most plasmas of these individuals, recurrent anti-RBD SARS-CoV-2 antibodies with potential neutralizing activity were detected, suggesting RBD as the efficiency target for neutralizing the viral disease [35]. However, other studies showed that RBD structures of SARS-CoV-1 and SARS-CoV-2 present some differences, and therefore, only a few antibodies against SARS-CoV-1 were able to neutralize SARS-CoV-2 [36].
The correlation between the level of antibodies produced and the severity of the disease even for other coronaviruses is still unclear [8]. Preliminary studies with SARS-CoV-2 indicate that the viral load and disease severity may be related to the magnitude of IgG antibodies [33, 37]. While IgM appears both in severe and nonsevere cases at the same time, IgG appears 2 to 3 days earlier in severe cases [37, 38]. Moreover, S protein mutations may be responsible for viral resistance to antibody recognition and consequent aggressiveness [29]. Caution is needed since the total measurable antibody is not precisely the same as protective. SARS-CoV-2 RBD of the S protein (RBD-S) is highly immunogenic. Antibodies that bind to this domain, blocking the interaction with Angiotensin-Converting Enzyme 2 (ACE2) (the host's input receptor) are characterized as potentially neutralizing. Virus-neutralizing antibodies are presumed to be an important mechanism of action in COVID-19 since high titers of these antibodies have been related to severe cases of the disease. In addition, some patients present low or very low titers of COVID-19 neutralizing antibodies up to 2 weeks after hospital discharge [38, 39].
Studies report successful plasma convalescent therapy as a source of therapeutic polyclonal antibodies, indicating monoclonal antibodies as an excellent therapeutic possibility [40]. However, a few researchers are concerned about the possibility of antibody-dependent enhancement (ADE), where the preexistence of anti-SARS-CoV-2 antibodies could exacerbate the immune response to SARS-CoV-2. One of the findings that support this hypothesis is that patients who developed responses against protein S earlier are related to the worse prognosis of the disease [41]. Another possibility is that the virus-specific nonneutralizing IgG could facilitate the entry of viral particles into cells that express Fc receptors (FcR), leading to the inflammatory activation of these cells, such as macrophages and monocytes [30]. This event was also observed during the SARS-CoV-1 epidemic, where deceased patients presented much higher neutralizing antibody titers when compared to patients who recovered [41]. So far, there is no evidence that antibodies developed for SARS-CoV-2 present such characteristics.
Studies with animal models showed that serum from rats vaccinated with SARS-CoV-2 RBD [42], as well as monkeys immunized with an inactivated SARS-CoV-2 vaccine candidate [43], were successful in obtaining antibodies and showed no ADE evidence. However, this hypothesis must be validated by further scientific research and also by longer time intervals in order to assess an effective and safer concentration of antibodies. With regard to vaccine aspects, the role of follicular helper T cells in the production of antibodies should also be considered to guarantee comprehensive protection. Juno et al. in 2020 found that antigen-specific antibodies, memory B cells, and follicular helper T cells are elicited after SARS-CoV-2 infection and correlated to potent neutralizing responses, suggesting that T cells together with B cells should be investigated to evaluate the potency of antigen-based vaccines [44].
The immunological scenario regarding COVID-19 indicates a secondary role of humoral response in terms of effectiveness, suggesting the importance of innate and cellular response.
2.2. Cellular Immune Response Dysregulation: Innate and Adaptive Immunity
Some studies described that the acute phase of COVID-19 is divided into two aspects: benign and severe, and it is correlating with immune dysfunction alterations [13–16]. The literature showed that macrophages and neutrophils may be responsible for the high activation of inflammatory pathways, which exacerbates the production of proinflammatory cytokines and chemokines, leading to the “cytokine storm” phenomenon [13, 45, 46].
A highly activated innate immune response may be the cause of disease progression, as well as poor outcome, including death [2, 47, 48]. Wang et al. in 2020 showed that natural killer (NK) cells were decreased in peripheral blood from COVID-19 patients, which may be related to these cells' function in the lung compartment, contributing to greater inflammatory responses [6, 45]. T lymphocytes are also involved in the immune imbalance during COVID-19, with a massive decrease of CD4 and CD8 total numbers in the initial phase [37], and with CD4 T helper cells found in the pulmonary tract [45], showing a possible effect of SARS-CoV-2 on lymphocyte lysis and an important function of CD4 T helper cells in the lung damage in the acute phase [2, 45] (Figure 1).
Figure 1.
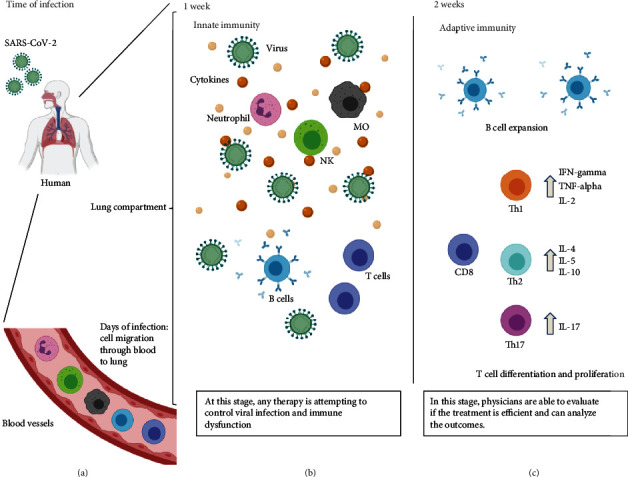
Immune response during COVID-19. (a) After SARS-CoV-2 contact with the respiratory tract of a human, susceptible cells are infected by entry through ACE2 or CD147 surface receptors. Once in the lung compartment, the first line of defense made of dendritic cells and other cells, such as macrophages, are able to recruit other immune cells. Then, cell migration through blood vessels are directed to the lung compartment, where cell activation occurs, initiating the inflammation process on the first week of signals and symptoms (b), inducing natural killer cell (NK), neutrophil, and macrophage (MO) activation in the lungs, causing the phenomenon called “cytokine storm.” In the mean time, B cells are producing antibodies, and immature T cells are also activated attempting to control the viral infection, although apoptotic events can also occur, reducing the number of T cells in the lungs and in the peripheral blood. After that, in two or more weeks (c), adaptive immune response is raised to specific IgG and neutralizing antibodies to contribute to the recovery and viral elimination. At this time, CD8 T cells are activated as cytotoxic phenotype, with the same aim, driving towards virus elimination. Meanwhile, the differentiation of CD4 T cells can trigger for more help for viral elimination by Th1 cells, but it can also lead to more lung damage by inflammatory phenotypes, such as Th2 and Th17 cells. Physicians and clinicians should evaluate each case to introduce the immunotherapies attempting to control the immune dysfunction. Created with BioRender.com.
Wang et al. in 2020, showed that SARS-CoV-2 infects T cells through the spike S protein, and the entry is facilitated not just by the ACE2 receptor, but also by the CD147 expression at the cell surface [49, 50], which may contribute to immune cells carrying the virus to other organs.
Since the beginning of the pandemic, the event termed as “cytokine storm” has been the focus of several studies and cytokine and chemokine production has been used as a predictor of severity or death, or as means to evaluate therapeutic targets [13, 46, 51]. Cytokines/chemokines IL-1β, IL-6, IL-7, IL-8, IL-9, IL-10, CXCL10, CCL2, CCL3, CCL4, TNF-α, IL-12, and IL-17, have been described in high levels on plasma/serum samples from patients with severe COVID-19 and even higher in patients prior to death [40]. Natural killer cells, macrophages, and immature T cells can produce these components, being chemoattractant to neutrophils in the lung compartments, inducing a reduction of blood vessels, and subsequently, in the blood circulation, leading to oxygen impairment and facilitating thrombosis activation [45, 47, 52] (Figures 1–3).
Figure 2.
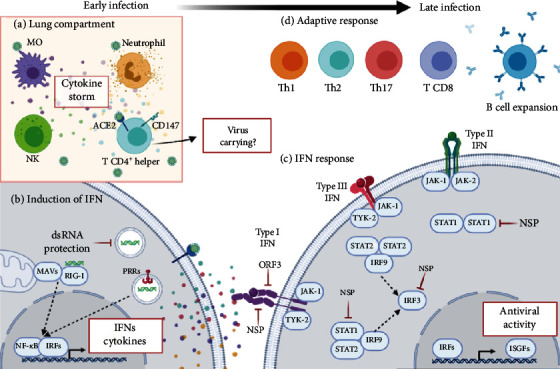
Summary of immunopathogenesis in early and late SARS-CoV-2 infections. (a) Early in the lung compartment, activated macrophages, neutrophils, NK cells, and T CD4+ helper cells release an exacerbated production of an immune mediator called “cytokine storm” that contributes to lung damage. Entry of T CD4+ helper cells is facilitated not just for ACE2 and CD147 receptors, which may contribute to carrying virus to other organs. (b) Still in the early phase, virus-cell interaction leads to an innate cellular response. During viral replication, dsRNA is recognized by RIG-I that interacts with MAVs leading to the activation of NF-κB transcription factors and IRF which induce the expression of IFNs and proinflammatory cytokines. However, SARS-CoV-2 can evade PRRP recognition shielding dsRNA by membrane-bound compartments that form during viral replication. (c) Following IFN release, the first activated pathway is type I IFN. IFN-α/β interaction with IFNAR receptor activates TYK-2 and JAK-1. This interaction could be blocked by SARS-CoV-2 by viral ORF3 and NSP. Virus can interact directly with STAT1 inhibiting interaction with STAT2 and IRF9. Next, type III IFN (IFN-λ) interaction with IFNLR/IL-10R leads to activation of TYK-2 and JAK-1, followed by STAT2-IRF9 interaction. Both type I and III IFN pathways activate IRF3, which is a target of viral NSP proteins. The last IFN pathway activated after SARS-CoV-2 infections is type II IFN (IFN-γ), related to adaptive responses. IFN-γ interaction with IFNGR receptor activates JAK-1 and JAK-2, followed by STAT1, target of viral evasion by NSP. All IFN pathways lead to expression of antiviral ISGFs. (d) Finally, the cellular profiles change to adaptive immune response with B cell expansion and specific antibody production. At this time, CD8 T cells are activated as a cytotoxic phenotype, and CD4 T cells differentiate in Th1 cells that help viral elimination, as well as inflammatory phenotypes Th2 and Th17. Created with BioRender.com.
Figure 3.
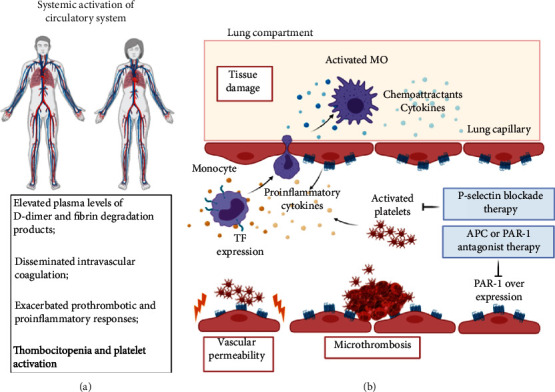
Hemostatic disorder in SARS-CoV-2 infection. (a) In COVID-19 cases, there occurs a systemic activation of the circulatory system as a reflection of what occurs in the lungs. (b) In the lung compartment, monocytes expressing TF perform an intense diapedesis under stimuli of proinflammatory and chemoattractant cytokines, infiltrating activated macrophages in the lung that lead to tissue damage. The high production of proinflammatory cytokines, as well as vascular permeability and microthrombosis events are prompt to overexpress PAR-1 receptors and abnormal platelet activation. Overexpression of PAR-1 can be blocked with APC or PAR-1 antagonist therapy, while abnormal platelet activation can be avoided by P-selectin blocker therapy. Created with BioRender.com.
The innate immune response is the organism's first line of defense, and an effective response depends on the balance between activation and regulation of the patient's immune system. Among these, type I and III interferons (IFNs) are considered the most important for antiviral defense, which mediate the onset of the innate immune response. Upon activation of pattern recognition receptors (PRRs), downstream signaling cascades trigger the secretion of cytokines [53]. Early on, coronavirus (CoV) prevents recognition by PRRs, and dsRNA is first shielded by membrane-bound compartments formed during viral replication of SARS-CoV-1 [54].
Interferon cytokines affect several other processes, including those regulating cell growth, differentiation, and apoptosis, as well as the modulation of the immune response. The main receptor is retinoic acid-inducible gene I (RIG-I) that detects viral RNA and interacts with mitochondrial antiviral signaling proteins (MAVs), leading to the activation of nuclear factor kappa B (NF-κB) transcription factors and interferon regulatory factor family (IRF) which induces the expression of IFNs and proinflammatory cytokines. Studies using BALB/c mice showed that early IFN-I response reduced SARS-CoV titer and regulated inflammation and mild clinical disease [55]. Furthermore, SARS-CoV-2 is sensitive to IFN-I pretreatment [56] as demonstrated by in vitro assays. After binding to their receptors, IFN-I and III initiate the activation of Janus kinase 1 (JAK-1) and tyrosine kinase 2 (TYK-2). Phosphorylation recruits the signal translation and transcription activators (STAT1 and 2), forming a heterodimer of interferon regulatory factor (IRF). This complex forms the gene factor stimulated by interferon (ISGFs) that translocate to the nucleus, binding to the IFN response element (ISRE), inducing the production of hundreds of ISGs [53].
Type I and III IFNs are activated after a viral infection, and several reports suggest redundant functions. However, the transcription of these genes is regulated over time, since type I IFNs are expressed and resolved quickly, while type III IFNs present late, sustained, and tissue-specific induction. On the other hand, type II interferon (IFN-γ) is synthesized almost exclusively by the activated natural killer (NK) cells and activated T cells, in response to virus-infected cells. It also plays a role in mediating protection against viral infections, especially long-term control of viral infection. Studies showed that high levels of IFN-γ were associated with greater viral load and lung damage [57]. In contrast, suppressed IFN-γ production by CD4+ T cells might be correlated with the disease severity of COVID-19 [17] (Figure 2). However, SARS-CoV-2 present several evasion mechanisms of the innate immune response, and the genome viral load encodes a protein open reading frame (ORF3b) that inhibits the induction of type I interferon [58]. Analysis of the virus sequences isolated from two patients with severe COVID-19 disease showed a variant that further increased the ability of ORF3b to suppress interferon induction [59].
The other CoV, ORFs and nonstructural proteins (NSPs) interact directly or indirectly with PRR signaling cascades and IFN signaling pathway proteins, such as TBK1, IRF3, IFNAR1, and STAT1 [60]. Besides, CoV antagonizes RNaseL and STING, consequently impairing the development of an effective innate immune response [30]. The key point in SARS-CoV-2 infection could be the effect of viral evasion of the host defenses, related to the innate immune response. Thus, suppressing the production of INF cytokines at the right time could elicit an effective response against SARS-CoV-2 (Figure 2).
After 15-20 days of signals and symptoms, people who recovered from COVID-19 are presenting a reestablishment of their lymphocyte count, with an increase of CD4 T and CD8 T memory cells specific for SARS-CoV-2 protein [61, 62]. Nevertheless, an imbalance of total lymphocytes (CD3+) is not frequent for all patients, and this observation could be related to aging [15, 28]. Even with the current limitation of serological assays related to antibody detection, some studies have shown that memory T cells are emerging from peripheral blood to contribute to virus elimination and complete recovery [61–63] (Figure 1).
Despite those findings, there are challenges concerning the full protection from possible reinfection. The protection after infection and the presence of antibodies that do not guarantee the “immunity passport” is still poorly understood, as discussed [13, 62, 63]. Considering the above, more studies are necessary, and at the same time, new information is appearing daily with regard to immunity against the new coronavirus.
2.3. Hemostatic Disorders in COVID-19: A Consequence of Immune Events?
Patients with severe COVID-19 commonly exhibit signs of systemic activation of the coagulation system that relies on increased rates of thrombotic events and elevated plasma levels of D-dimer, a known marker of thrombogenesis [64–67]. In clinical observations, nonsurvivors present significantly higher levels of D-dimer and fibrin degradation products as well as longer prothrombin and activated partial thromboplastin compared to survivors on admission, thus meeting the criteria for disseminated intravascular coagulation [57, 67]. The mechanisms of COVID-19-driven coagulopathy are not fully understood but are possibly associated with the excessive immune/proinflammatory response [13, 68], which ultimately leads to the activation of several prothrombotic pathways [69, 70]. Therefore, there is strong evidence that the exacerbated procoagulant responses account for organ failure and the increased mortality in critically ill COVID-19 patients [71]. In this context, treatment with anticoagulants may improve the progress of severe COVID-19 [72, 73].
The crosstalk between inflammation and coagulation has long been described [74]. One of the major sources of these cytokines is activated monocytes/macrophages and neutrophils (Figure 3). During the infection, high plasma levels of CCL2 and CCL3, potent chemoattractants for monocytes, are noted [75]. Moreover, CCL2 and CCL7 are reported in bronchoalveolar lavage fluid from severe patients [76]. Therefore, these cells infiltrate massively into the lungs, as demonstrated in fatal cases of COVID-19 [36, 47].
Adding to that, dysregulated T cells secrete GM-CSF, which, together with IL-6, turn infiltrated monocytes into activated CD14+CD16+ macrophages [77]. These cells, still inside capillaries, probably express Tissue Factor (TF), as demonstrated largely in a number of viral infections, such as HIV and dengue, as well as microbial and other types of sepsis [78–80].
In the absence of tissue damage, TF expressed by activated monocytes initiates TF-dependent coagulation with the conversion of prothrombin to thrombin (Coagulation Extrinsic Pathway), in an uncontrolled manner, leading to disseminated intravascular coagulation. An increase in Protease-Activated Receptor 1 (PAR-1) expression in endothelial cells after viral infections is one of the consequences observed [81]. Although the main function of thrombin is to promote the formation of clots through the activation of platelets and the conversion of fibrinogen to fibrin, this protease exerts several cellular effects and may increase the inflammation process through the activation of PAR receptors, mainly PAR-1.
The activation of PAR-1 in endothelial cells promotes proinflammatory responses, resulting in the rupture of the endothelial barrier and an increase of cell permeability. [82]. These defects in procoagulant-anticoagulant mechanisms during inflammation may be responsible for predisposition of the development of microthrombosis, disseminated intravascular coagulation, and multiple organ failure in patients with severe COVID-19 pneumonia and in nonsurvivors [83] (Figure 3).
On the other hand, PAR-1 might be activated by the anticoagulant serine protease, activated protein C (APC), which binds to its coreceptor, endothelial protein C receptor (EPCR), and exerts cytoprotective and anticoagulant responses. Recombinant APC has been used as a drug to treat severe sepsis, a pathological condition characterized by exacerbated clotting and inflammation responses. However, the mechanism by which APC-activated PAR-1 promotes cytoprotective responses is still poorly understood [82]. In this context, PAR-1 antagonists and other coagulation protease inhibitors, in addition to modulators of the protein C pathway, may play an important role in the treatment of critically ill patients with COVID-19 [84].
Still, in the context of hemostatic disorders, platelets are also an important hub of classical crossover between the inflammatory and coagulation process [85–87]. Lungs are responsible for approximately 50% of platelet production and a site of mature and immature megakaryocytes [88], enhancing the implication of these anucleated cells in COVID-19 outcomes. From a clinical point of view, thrombocytopenia has been demonstrated in approximately 20% of in-hospital COVID-19 cases, with a strong association with thrombocytopenia and COVID-19 mortality. This event is most likely due to platelet consumption leading to the formation of pulmonary thrombi, which highlights the importance of monitoring platelet count during hospitalization [89, 90]. Besides, low count leads to platelet deposition in damaged pulmonary blood vessels in SARS patients [91, 92].
It has been described that activated platelets play a critical role in hemostasis and in coagulation, as well as in angiogenesis, inflammation, and even immune response, as demonstrated in other viral infections, like dengue and influenza. This activation occurs via P-selectin/prostaglandin-1 (PGL1), given viral engulfment and virus-platelet interaction [93]. In this way, a platelet RNA sequencing study of 25 COVID-19 patients demonstrated that platelets do not express the ACE2 receptor, although mRNA from the SARS-CoV-2 N1 gene was detected in platelets from 2/25 COVID-19 patients, suggesting that platelets may take-up SARS-CoV-2 mRNA independent of ACE2. Furthermore, resting platelets from COVID-19 patients increased P-selectin expression, both basally and upon activation [94].
With regard to immunological responses, platelets present multiple immune receptors, such as immunoglobulin or complement receptors and Toll-like receptors (TLRs) [85, 94], and the capacity of NRLP-3 inflammasome activation and production of cytokines IL-1β and TGF-β [95–97]. These functions are achieved through direct interaction with endothelial cells, as well as monocytes, lymphocytes (platelet-leukocyte aggregates (PLA)) and neutrophils (platelet-neutrophil aggregates (PNA)) in a process regulated by P-selectin [93, 98, 99]. PLA formation, specifically with monocytes, induces fibrin clot formation via interaction of PGL1 with tissue factors on the platelet surface [98], reduced IL-17 and IFN production when platelets adhere to CD4+ T-cells [99], and cross-presenting of antigens to CD8+ T cells given platelet expression of the major histocompatibility complex class I (MHC-I) [100]. The formation of PNA leads to C3 release from platelets and formation of Neutrophil Extracellular Traps (NETs), a mechanism that tightly regulates host immune responses and complement system responses, but can also promote thrombosis and damage lung capillaries [101, 102].
In COVID-19 patients, it has been demonstrated that circulating platelet-neutrophil, platelet-monocyte, and platelet-T-cell aggregates were all significantly elevated, enhancing the hypothesis that P-selectin blockade may be warranted in treating COVID-19 patients [94, 103, 104] (Figure 3).
3. Immunotherapeutic Insights: Important and Promising Breakthroughs
In fact, there are still no therapeutic agents for the effective treatment of COVID-19. Immunotherapy is described as a helpful tool to attempt to control immune dysfunction. However, to this moment, therapeutic intervention has been limited to severe cases, in order to minimize the risk of death, and also in combined therapies. Beyond therapies addressed to severe cases, it should be relevant to consider the differences on gender, age, pregnant condition, diabetes, hypertension, autoimmune diseases, heart diseases, cancer, and obesity during nonclinical and clinical studies, as well as the management for mild cases to avoid the worst outcome, since some comorbities are under risk of fatal COVID-19 [48, 105–107]. Actually, the surveillance of the immune events and their behavior should also be included in the guidelines for immunotherapies to treat COVID-19, once the immune response is crucial to the clinical evolution of the patient (Figure 1).
It is worth noting that some potential immunotherapies are very specific when targeting immune events and pathways, and they are designed to avoid adverse events, to be used exclusively to minimize the inflammation process induced by the viral disease, such as COVID-19. Therefore, immunotherapy approaches are developed to be useful in any situation, regardless of comorbity conditions. In this manuscript, we summarize some commercially available immunotherapeutic components with their mode of action and immunological impact in attempting to control the immune dysregulation during COVID-19, already used in clinical trials (human) (Tables 1(a) and 1(b)) and nonclinical studies (in vitro and in vivo animal models) for treatment purposes (Table 2).
Table 2.
Nonclinical studies focused on immunotherapy with potential benefits to COVID-19 treatment.
| Product | Potential benefits for COVID-19 | Reference |
|---|---|---|
| Daclatasvir | Antiviral compounds with immunomodulatory effect: reduce viral load and proinflammatory cytokines using in vitro cell cultivation with SARS-CoV-2 | Sacramento et al. [160] |
| Bovine lactoferrin | Antiviral compounds with immunomodulatory effect: reduce viral load and possible reduction of proinflammatory cytokines and induction of T cell activation | Carvalho et al. [167] |
| Kepi—inhibitory subunit of the protein phosphatase 1 complex (PP1) | Inhibition/blockade of immune response pathways: reduce signaling of inflammatory cytokines, as TNF-α in mice models infected with SARS-CoV | McDermott et al. [176] |
| Hybrid IFN-α B/D | Inhibition/blockade of immune response pathways: it was effective to inhibit SARS-CoV replication in Vero cells by in vitro assays and could reduce SARS-CoV replication in the lungs of infected mice | Barnard et al. [177] |
| CCR2 antagonist/anti-CCR2 | Inhibition/blockade of immune response pathways: may promote monocyte egress from the bone marrow and monocyte recruitment in tissues | Merad and Martin [140] |
| IRAK4 inhibitor/PF-06650833; CA-4948 | Inhibition/blockade of immune response pathways: may reduce inflammation caused by macrophages, mediates TLR and IL-1β signaling | Merad and Martin [140] |
| PAR-1 antagonist (SCH530348), PAR-1 antagonist receptor (SCH79797) | Anticoagulants: may reduce levels of proinflammatory cytokines, neutrophilic lung inflammation, and other harmful pathological effects that are associated with the disseminated intravascular coagulation in COVID-19 | Khoufache et al./Bacil et al. [179, 183] |
| Inhibitor of Bruton's tyrosine kinase (BTKi) | Immunotherapeutic approaches with similarities to other immunopathological diseases: may reduce B cell responses and macrophage polarization, as observed in patients with B cell malignancies | Chong et al. [180] |
Our search data was based on available findings up to July 20th, 2020. Furthermore, the number of registers with status “completed” or “terminated” (https://clinicaltrials.gov) was considered in order to determine clinical trials with results disclosed. Ongoing clinical trials were set up with a number of registers under “recruiting” status (https://clinicaltrials.gov). In addition, the clinical trials with preliminary results in humans in the preprint platforms and/or already published and/or recommended as treatment by published manuscripts were also considered. Nonclinical studies were classified as bioinformatic tools, cell cultivation assays, or animal models, with findings in the preprint platforms and/or already published with SARS-CoV-2, respiratory viral infections, or other diseases with similar immunopathological disorders, with recommendations to COVID-19 treatment by authors, or registration as a clinical trial with status varying from “recruiting,” “completed,” or “terminated.” The search for literature information was performed in digital medical science libraries, such as Google Scholar, SciELO, PubMed, bioRxiv, and medRxiv. From the bibliography found, only manuscripts with full text available were considered. The keywords used were severe acute respiratory syndrome, SARS-CoV-2, COVID-19, immunotherapy, immunomodulators, immunology, and therapy.
Considering the need for solutions and the speed of results, there are clinical trials approved, but without disclosure of preliminary results. According to these findings, we have been able to also show immunomodulatory products currently in test with the mode of action, purposes, and possible benefits to humans used in the treatment of COVID-19.
3.1. Clinical Trials: The State of the Art about the Tested and Ongoing Immunotherapies
3.1.1. Antibody Plasma
Neutralizing antibodies from convalescent plasma have been used as an alternative to suppress viremia of SARS-CoV-2 in hospitalized patients, shortening the hospital stay, thus reducing mortality rates [108]. There is a risk that other infections will be installed through blood transfusion if the pathogen is not screened according to health practice guidelines and recommendations [108, 109]. Another fact regarding convalescent plasma is related to the possibility of applying it to a large number of patients, but it has been useful for the treatment of a few COVID-19 severe or critical patients [41, 110]. It is noteworthy that convalescent plasma could help the immune system to raise virus-specific T cells and improve viral elimination [30].
Passive immunity through intravenous gammaglobulin (IVIG) has been used widely for inflammation-related diseases, supplying the production of antibodies, suppressing inflammatory cytokines, and modulating T cell responses. This treatment has been used for COVID-19, not only using purified human plasma but also using purified plasma from animals. However, adverse events can occur, such as renal failure, and therefore, this treatment should be addressed with caution [41, 110] (Table 1(a)).
3.1.2. Antiviral Compounds with Immunomodulatory Effect
Chloroquine (CQ) and hydroxychloroquine (HCQ) have been highlighted as important components to treat COVID-19. Both drugs have been used for autoimmune disorders, such as rheumatologic diseases with immunomodulatory effects, where they can enhance regulatory T cell activity and attenuate the “cytokine storm” [111]. In vitro studies with SARS-CoV-2 showed that these drugs inhibited endosomal TLR7 and TLR9 directly inducing endosomal maturation, thus driving the viral particles to lysosomes. However, CQ and HCQ already used on clinical trials show some controversial results with respect to the benefits for patients, demonstrating no efficiency on clinical results for COVID-19 treatment [30, 111] (Table 1(a)).
Ivermectin, an FDA-approved broad-spectrum antiparasitic drug has been reported for its antiviral activity against SARS-CoV-2 when used through cell cultivation [112]. This drug plays an important role in paralyzing parasites and in viral replication mechanisms, described already for several viruses [113]. Ivermectin was associated with lower mortality during the treatment of COVID-19 [114]. Additionally, an immunomodulatory effect of this drug in animal models and humans has been shown, including an increase of antibody production and leukocyte count [113, 115, 116]. Therefore, the immunomodulatory effect of ivermectin in COVID-19 should be better explored, though for now there are no changes in the evidence of immune response during COVID-19 described yet.
Oseltamivir is a very well-known drug to treat influenza virus infection, demonstrating effectiveness in reducing the viral load as well as inflammation, morbidity, and innate immune responses, and affecting the magnitude of the effector CD8+ T cell responses. Results on the immunomodulatory effects of Oseltamivir were observed in animal models and humans [117, 118]. Oseltamivir was also able to reduce the number of days of hospitalization for patients with COVID-19 [119] (Table 1(a)).
Atazanavir, an antiviral compound with antiretroviral protease inhibitor activity used in HIV patients, was able to reduce SARS-CoV-2 replication in tumor cell lines as well as in decreasing proinflammatory IL-6 and TNF-α levels in the supernatant of peripheral blood mononuclear cells in cell culture with the virus [120]. However, the immunomodulatory effect levels of IL-6 and TNF-α could be explained by the reduction of the viral load using Atazanavir in vitro. Now, there is an ongoing clinical trial testing this drug on COVID-19 patients (Table 1(b)).
3.1.3. Cytokine-Directed Therapy
Interferons type I (alpha)/(beta) were widely used to treat several viral infections worldwide, due to their nonspecific antiviral ability, participating in the first line of defense [53]. Normally, interferon type I is used in combined therapies for viral infections [53, 121, 122]. Recently, Hadjadj et al. observed that interferon type I was impaired in the early acute phase of COVID-19 patients [122]. Robust IFN-I responses have also been observed in the respiratory tract and peripheral blood of patients with severe COVID-19 with high levels of interferon-stimulated genes (ISGs) along with high proinflammatory responses [123, 124]. Hung et al. in 2020 showed preliminary results from a phase 2 clinical trial with COVID-19 patients using 0.25 mg of commercial interferon beta-1b combined with other antiviral drugs (lopinavir/ritonavir/ribavirin). This combination was able to reduce the duration of viral shedding and days of hospitalization and mitigation of symptoms [125]. In a multicenter clinical trial, with 446 COVID-19 patients in Hubei, China, Wang et al. in 2020 demonstrated that early use of IFN-α decreased mortality, but its late use increased mortality and delayed recovery. In this way, the use of IFN therapy in COVID-19 needs precision medicine approaches, considering the favorable antiviral activity of IFN together with other direct antiviral therapies in early/mild infection and enhancement of harmful cytokine storm events in severe/late infections [126].
3.1.4. Inhibition/Blockade of Immune Response Pathways
Some studies have been directed at better exploring the role of corticosteroids as a means to impair the immune cell response during SARS-CoV-2 infection [51]. However, some caution is needed since corticosteroids can induce the “shutdown” of the immune cell response and increase the viral infection process, raising the viral load and lysing lung cells [127, 128]. Corticosteroids can induce cell death and impair cytokine production by macrophages, neutrophils, and T cells. This component can promote the production of annexin 1, leading to the inhibition of the phospholipase A2 expression, responsible for pain and inflammatory events. Furthermore, corticosteroids can increase the expression of inhibitory proteins, leading to NF-κB inactivation, reducing the transcription factors involved in Th2 type cytokine expression [129, 130].
Despite possible issues that could complicate the viral infection, a corticosteroid compound, Dexamethasone, has been described as a more successful therapy used until now for COVID-19 treatment in patients in intensive care units (ICU) with a severe grade of the disease, reducing the number of deaths [131–133]. Corticosteroids such as Methylprednisolone were also found to help in the treatment for COVID-19 [134] and other respiratory infections, including influenza [135], reducing the mortality rates for hospitalized patients [136].
As described here, hyperinflammation caused by SARS-CoV-2 infection leads to the “cytokine storm” [30, 111, 137]. In an attempt to control the exacerbation of lung inflammation, cytokine inhibitors and blockers of inflammatory pathways are in focus and under analysis to be used as therapy, becoming promising for the treatment of COVID-19. Some ongoing clinical trials are testing the efficacy of commercial anti-IL-6R (Tocilizumab/Actemra; Sarilumab/Kevzara) and anti-IL-6 antibody (Siltuximab/Sylvant) on COVID-19 patients used in conjunction with other therapeutic combinations (antiviral drugs and oxygen therapy). One group has already described preliminary data in a small population (n = 20), showing that patients recovered lymphocyte counts (52.6%), in which 95% of them presented a resolution of lung opacities on chest computed tomography as well as being discharged from the hospital, using Tocilizumab [138].
Perspectives of clinical trials were presented in a recent review, enumerating the potential therapeutics capable of reducing inflammation, and pointing out the blockade of monocyte costimulators, such as CCR5 (Leronlimab, Emapalumab), which may regulate monocyte and T cell migration to the infected tissue. Additionally, trials evaluating additional antibodies to inhibit IL-1β (Anakinra), IFN-γ, and myeloid-derived inflammatory cytokines (GM-CSF/anti-GM-CSF) have been described [139, 140].
Using a bioinformatics tool strategy to search for promising compounds targeting immune responses, Richardson et al. in 2020 described that Baricitinib/Ruxolitinib, used for JAK/STAT inhibition and promoting impairment of signaling transduction on cell immune response, may be promising for COVID-19 treatment [141]. Ongoing clinical trials are using JAK inhibitors to evaluate their efficiency in trying to control the cytokine storm, limiting the cytokine expression by immune cells [140, 142].
The complement system plays an important role in the initial cascade of the inflammation process in viral infections, as well as in the production of cytokines/chemokines [143, 144]. It is not different in SARS-CoV-2 infection, in which platelets and complement components may be responsible for the thrombosis observed in some COVID-19 patients [101]. A commercial anti-C5 monoclonal antibody (Eculizumab), presented preliminary results with four patients in the intensive care unit (ICU) with COVID-19, in which all patients recovered after the treatment, diminishing the C-reactive protein and shortening the period of hospitalization [145].
A case report study, treating a child with Crohn's disease and affected by COVID-19, used anti-TNF (Infliximab) with positive results, demonstrated by an improved cytokine profile, with normalization of TNF-α, and a decrease of IL-6 and IL-8 concentrations in blood. These preliminary findings support a role for the blockade of TNF-α in the treatment of the COVID-19 inflammatory cascade, with possibilities extending to the use of other TNF treatments ((Infliximab, Adalimumab, Golimumab)/Fab′-PEG (Certolizumab) Fusion TNFR2-IgG1-Fc (Etanercept)) [146] (Table 1).
Table 1.
Clinical trials with results disclosure and ongoing clinical trials focused on immunotherapy to attempt immune regulation during COVID-19.
(a) Clinical trials with result disclosures
| Product | Benefits for COVID-19 | ClinicalTrials.gov reference number | Reference |
|---|---|---|---|
| Dexamethasone/methyl prednisolone | Corticosteroids: reduce hyperinflammation and mortality rates | NCT04445506; NCT04323592; NCT04244591; NCT04273321; NCT04374071 | Gangopadhyay et al./Chroboczek et al./Ledford/Wang et al. [131–134] |
| Convalescent plasma | Antibodies from plasma: reduce viral load, days of hospitalization, and the number of deaths. Increase of effector T cell responses helping in viral elimination | NCT04407208; NCT04346446; NCT04441424; NCT04442958 | Vabret et al./Shang et al. [30, 108] |
| Chloroquine and hydroxychloroquine | Antiviral compounds with immunomodulatory effect: without proven effectiveness or potential benefit in COVID-19 | NCT04307693; NCT04261517; NCT04434144; NCT04345861; NCT04376814; NCT04343768; NCT04423991; NCT04389320; NCT04441424; NCT04308668; NCT04323592; NCT04306497; NCT04362332; NCT04342650; NCT04323527; NCT04358068; NCT04321278; NCT04333654; NCT04475588; NCT04453501; NCT04308668; NCT04304053 | Vabret et al./Zhang et al. [30, 111] |
| Ivermectin | Antiviral compounds with immunomodulatory effect: reduce mortality rates. Possible effect on raising antibody production and leukocytes | NCT04343092; NCT04434144 | Rajter et al. [114] |
| Oseltamivir | Antiviral compounds with immunomodulatory effect: reduce the hospitalization days for COVID-19 patients | NCT04349241 | Coenen et al. [119] |
| Interferon β-1b | Cytokine-directed therapy: combined with other antivirals, reduces the duration of viral shedding, days of hospitalization and mitigation of symptoms | NCT04343768; NCT04276688 | Shalhoub [125] |
| Anti-IL-6 and anti-IL-6R (Tocilizumab/Siltuximab) | Inhibition/blockade of immune response pathways: combined with other antivirals, presented resolution on lung opacities on chest computed tomography and hospital discharge using tocilizumab | NCT04331795; NCT04346355; NCT04322188 | Xu et al. [138] |
| Heparin | Anticoagulants: a preliminary study with D-dimer elevation levels, high platelet count in severe COVID-19 | NCT04412304 | Zhang et al. [111] |
| Azithromycin | Antibiotics: used as an adjuvant in therapies with antiviral drugs and immunomodulatory components to avoid secondary infections in hospitalized patients, with good results in mortality reduction and intubation shortening time | NCT04345861; NCT04434144; NCT04343092; NCT04441424; NCT04358068; NCT04321278; NCT04453501 | Saghazadeh and Rezaei/Falsenstein et al. [142, 153] |
| Multipotent mesenchymal stem cells (MSC) | Immune cell-based therapy: results showed a reduction of pulmonary edema and decreased circulating proinflammatory cytokines and mortality rates | ChiCTR2000029990 | Zhang et al./ Leng et al. [111, 154] |
| Vitamin D, zinc, Traditional Chinese Medicine | Biomolecule effect on immune response: contributing to the reduction of hyper inflammation | NCT04407572; NCT04435119; NCT04306497 | Zhang et al./Zhang et al./Meltzer et al. [111, 157, 158] |
(b) Ongoing clinical trials
| Product | Benefits for COVID-19 | ClinicalTrials.gov reference number | Reference |
|---|---|---|---|
| Intravenous gamma globulin (IVIG) | Antibodies from plasma: may suppress inflammatory cytokines and modulate T cell responses | NCT04411667; NCT04403269; NCT04400058 | Zhang et al. [111] |
| Anti-CCR5 (Leronlimab) | Inhibition/blockade of immune response pathways: may promote monocyte and T cell recruitment in tissues in COVID-19 | NCT04347239; NCT04343651 | Merad and Martin [140] |
| Recombinant IL-1 receptor antagonist (Anakinra) | Inhibition/blockade of immune response pathways: may reduce proinflammatory cytokines in COVID-19, as seen in patients with rheumatologic disorders | NCT04339712; NCT04324021; NCT04330638; NCT04443881; NCT04362943; NCT04412291; NCT04364009; NCT04357366; NCT04408326; NCT02735707; NCT04374539; NCT04278404; NCT04462757 | Muskardin [139] |
| Anti-IFN-γ (Emapalumab) | Inhibition/blockade of immune response pathways: may reduce pro-inflammatory cytokines in COVID-19 | NCT04324021 | Merad and Martin [140] |
| Anti-GM-CSF (Lenzilumab)/GM-CSF (Sargramostim) | Inhibition/blockade of immune response pathways: may reduce proinflammatory cytokines, and promotes macrophage differentiation and survival driving to tissue repair in lungs in COVID-19 | NCT04351152; NCT04326920; NCT04400929 | Merad and Martin [140] |
| JAK/STAT inhibition (Baricitinib/Ruxolitinib) | Inhibition/blockade of immune response pathways: may induce impairment of the signaling transduction on cell immune response using bioinformatic tools | NCT04390464; NCT04321993NCT04334044; NCT04351503; NCT04377620; NCT04362137; NCT04338958; NCT04348695; NCT04403243; NCT04359290; NCT04477993 | Richardson et al. [141] |
| Anti-C5 (Eculizumab) | Inhibition/blockade of immune response pathways: diminishing C reactive protein and shortening period of hospitalization | NCT04346797; NCT04351503 | Diurno et al. [145] |
| Anti-TNF (Infliximab) | Inhibition/blockade of immune response pathways: cytokine profile improved with normalization of TNF-α, a decrease in IL-6, and IL-8 concentrations | NCT04425538; NCT04344249 | Dolinger et al. [146] |
| Anti-CD147 (humanized Meplazumab) | Inhibition/blockade of immune response pathways: inhibition of SARS-CoV-2 entry and replication using in vitro cell cultures, may reduce T cell activation and T cell infectivity. Reduce viral replication in humans with COVID-19 | NCT04275245 | Wang et al./Ulrich and Pillat [147, 149] |
| rhACE2 | Inhibition/blockade of immune response pathways: inhibition of SARS-CoV-2 entry and replication as well as may control immune cell response | NCT04335136 | Zhang et al./Tu et al. [111, 150] |
| Natural killer cells (NK) | Immune cell-based therapy: may help in elimination of infected cells | NCT04365101; NCT04280224NCT04324996; NCT04375176 | Zhang et al./Tu et al. [111, 150] |
| Biomolecules: vitamin C, melatonin, nitric oxide (iNO) | Immunomodulatory effects of biomolecules: may contribute to reducing hyperinflammation and prevention of hypoxic respiratory failure | NCT04323514; NCT04357782; NCT04370288; NCT04328961; NCT04354428; NCT04264533; NCT04409522; NCT04353128; NCT04446104; NCT04326725; NCT04360980; NCT04368897; NCT04388683; NCT04338828; NCT04305457; NCT04337918; NCT04306393; NCT04312243; NCT04445246; NCT04334512; NCT03680274; NCT04335084; NCT04468139; NCT04421508; NCT04397692 | Zhang et al./Tu et al./Zhang et al./Parikh et al. [111, 150, 157, 159] |
| Atazanavir | Antiviral compounds with immunomodulatory effect: reduce viral load and proinflammatory cytokines using in vitro cell cultivation with SARS-CoV-2 | NCT04452565 | Merad and Martin [140] |
| Humanized monoclonal antibody to P-selectin (Crizanlizumab) | Anticoagulants: anti-P-selectin may help to maintain the hemostatic homeostasis, being an essential cofactor for the extrinsic pathway of blood coagulation | NCT04435184 | Neri et al. [103] |
| CAR-T cell therapy | Immunotherapeutic approaches with similarities to other immunopathological diseases: may help the elimination of infected cells | NCT04474067 | Bachanova et al./Ljungman et al. [155, 156] |
Wang et al. in 2020 performed in vitro assays showing that CD147 detected on the surface of lymphocytes and Vero cell lines can be an alternative entry for SARS-CoV-2 in host cells [147]. Koch et al. in 1999 showed that CD147 is a broadly expressed cell surface glycoprotein, whose expression is upregulated upon T cell activation [148]. Grifoni et al. in 2020 showed T cell activation by the SARS-CoV-2 protein in patients that recovered from COVID-19 and in healthy subjects with a history of human common cold coronaviruses [17]. Probably, T cell activation can facilitate the entry of SARS-CoV-2 into T cells through CD147 expression, becoming easily disseminated to other tissues through blood circulation. According to those findings, Wang et al. in 2020 also showed that an anti-CD147 monoclonal antibody could inhibit SARS-CoV-2 replication using in vitro cell cultures, being useful for near-future therapies [147]. Currently, a clinical trial is ongoing using a humanized anti-CD147 (Meplazumab) with a potential beneficial effect on COVID-19 treatment [149].
Some review studies have mentioned the use of recombinant human Angiotensin-converting enzyme 2 (rhACE2) to block the entry of SARS-CoV-2 in susceptible cells, contributing to the improvement and control of the immune response during the viral disease. The evaluation of this immunotherapy showed good results using in vitro cell culture and in clinical trials [111, 150].
3.1.5. Anticoagulants and Their Effects on Immune Events
Anticoagulant compounds play an important role in preventing the consumption of multiple clotting factors in disseminated intravascular coagulation (DIC) [80], as observed in COVID-19 [67, 151, 152]. Anticoagulant heparin was used in a few patients with severe COVID-19, with important findings on the D-dimer elevation levels and high platelet count, but without presenting any difference with regard to poor outcomes. A large sample size is needed to confirm the effects of heparin in the management of COVID-19 patients [72, 111].
Other compounds in use for hemostatic disorders were observed in COVID-19 patients. Crizanlizumab, a humanized monoclonal antibody to P-selectin recently approved for use in patients with sickle cell anemia, has been pointed as a therapeutic approach. P-selectin is related to the platelet function to maintain hemostatic homeostasis, being an essential cofactor for the extrinsic pathway of blood coagulation. Neri et al. in 2020 described that P-selectin plays a role in leukocyte recruitment into the lungs during SARS by other studies using animal models and cell culture thus suggesting that the monoclonal antibody is a possible therapy to treat COVID-19 [103]. There is an ongoing clinical trial testing this monoclonal antibody on COVID-19 patients (Table 1(b)).
3.1.6. Antibiotics with Immunomodulatory Effects
Antibiotics are normally used as an adjuvant in therapy with antiviral drugs and immunomodulatory components to avoid secondary infections, such as bacterial and fungal infections in hospitalized patients. Despite their antimicrobial function, antibiotics like Azithromycin present immunomodulatory properties, which can reduce inflammatory macrophage polarization and inhibit NF-κB signaling pathways, minimizing the hyperinflammation damage. Since the beginning of COVID-19 treatment, antibiotics have been used with good results in mortality reduction and shortening of intubation time [142, 153].
3.1.7. Immune Cell-Based Therapy
Multipotent mesenchymal stem cells (MSC) derived from placenta, bone marrow, umbilical cord, or adipose tissue have been highlighted in the tissue engineering field to treat several diseases with immunomodulatory properties. Nonclinical studies have shown the efficacy of MSC in the treatment of respiratory diseases, reducing pulmonary edema, decreasing circulating proinflammatory cytokines, and improving mortality rates. On COVID-19 patients, MSC therapy was demonstrated to be effective using allogeneic MSC transplantation without adverse effects in seven patients [111, 154].
As mentioned here, NK cells play an important role in the innate immunity during COVID-19, and their counts dropped in infected patients. Given these findings, NK cell-based immunotherapy is one option to treat COVID-19. It has been under phase I of clinical trials, and companies are aiming at repurposing their anticancer NK-based products, in which they have developed placenta hematopoietic stem cell-derived NK cells, to COVID-19 treatment. Therefore, this appears to be a promising immunotherapy for the near future [111, 150].
Regarding the context of immune cell-based therapy, some authors described the possible use of CAR-T cell therapy to treat COVID-19 patients, to compensate for the loss of T lymphocytes, to help in the elimination of infected cells. Nevertheless, there are some concerns about patient selection, and guidelines should be evaluated in each case to use this therapy. It has demonstrated effectiveness in cancer therapy, and the issues involved with the treatment of COVID-19 are related to other adverse events. Once the CAR-T cell therapy is initiated, the process cannot be aborted [155, 156]. However, no evidence of the success of these products in the management of COVID-19 treatment has been disclosed yet (Table 1).
3.1.8. Biomolecule Effect on Immune Response
In acute lung injury, like SARS, it has been discussed that excessive neutrophil accumulation in the lung tissue that induces severe damage through the release of necrotic cell contents contributes to hyperinflammation and hypoxia. Biomolecules, including compounds from Traditional Chinese Medicine, vitamin C, vitamin D, zinc, melatonin, and nitric oxide are reported to relieve those processes, being promising immunomodulator candidates in immunotherapy against COVID-19 [111, 157].
A study reported that vitamin D deficiency that is not sufficiently supplied is associated with COVID-19 risk [158]. Parikh et al. in 2020 showed that spontaneously breathing patients with COVID-19, who underwent therapy with inhaled nitric oxide (iNO), showed good results and no need for mechanical ventilation [159]. Their findings suggested that iNO therapy may play a role in preventing the progression of hypoxic respiratory failure. Other biomolecules related to good results in respiratory infections are also considered being promising in helping with COVID-19 recovery in combined therapies. Clinical trials are underway aimed at evaluating their effects.
3.2. Nonclinical Studies: State of the Art from Laboratory Findings and Other Diseases Related to Host Immune Disorders
3.2.1. Antiviral Compounds with Immunomodulatory Effects
A hepatitis C drug, Daclatasvir, is able to block hepatitis C nonstructural viral protein activities, essential for viral replication in the cell hosts. Sacramento et al. in 2020 demonstrated an inhibitory activity of Daclatasvir for SARS-CoV-2 using an in vitro cell culture system, showing similarities in antiviral activity for nsp12 protein, with a reduction of proinflammatory cytokines (IL-6 and TNF-α) in early events [160]. In addition, there is no current clinical trial for this drug carried out with COVID-19 patients (Table 2).
3.2.2. Antiviral and Immunomodulatory Effects of Biomolecules: Bovine Lactoferrin
Lactoferrin (Lf) is part of the transferrin family, being an iron-binding protein, which is present in several external secretions such as milk, semen, saliva, mucous secretions, pancreatic fluids, and gastrointestinal secretions [161, 162]. A recently published observational study using bovine lactoferrin in liposomal form showed a decrease in typical symptoms of COVID-19 in 75 patients. In this study, liposomal bovine lactoferrin (32 mg) was administered in the form of syrup together with vitamin C (12 mg) and zinc (10 mg) [163]. Regarding the antiviral activity of lactoferrin, a pioneer study using SARS pseudovirus demonstrated that lactoferrin was able to inhibit the entry of the virus by binding to heparin sulfate proteoglycans (HSPGs), preventing binding of the virus [164]. This same mechanism has demonstrated an inhibition of approximately 80% for Mayaro infection [165]. In that study, the authors demonstrate that the binding of lactoferrin to the Vero cells was highly dependent on HSPG sulfation. This mechanism strongly suggests that inhibition of the Mayaro virus infection occurs by blocking these molecules on the cell surface. Similar results were shown for Zika and Chikungunya virus [166]. More recently, lactoferrin has been shown to be able to inhibit about 85% of SARS-CoV-2 infection in Vero cells [167].
Another recent study, using the influenza virus, showed the action of bovine lactoferrin on the viral particle. The results showed an interaction between lactoferrin and viral hemagglutinin at low pH, which led to the stabilization of this protein, thus preventing the conformational changes required for the virus-cell interaction and consequently the beginning of the viral cycle [168]. In the immunological context of lactoferrin action, it is known that it has an anti-inflammatory action, produced mainly by macrophages and neutrophils, acting in several cascades of cell signaling [169, 170]. Although the bioavailability of iron is fundamental for several metabolic processes, free forms of this ion can be potentially harmful by inducing the formation of reactive oxygen species (ROS). Due to the ability of lactoferrin to bind several ions, including iron, it can act as a chelator for this ion, decreasing its availability and consequently decreasing the inflammatory process by controlling iron homeostasis [169, 171]. In addition, lactoferrin has the ability to control the translation and release of inflammatory molecules, such as IL-1α, IL-1β, IL-6, IL-8, TNF-α, IFN-α, IFN-β, and other chemokines, with the function of activating the adaptive immune response [169, 172, 173]. Although more studies are needed to reveal the action of lactoferrin in more detail, the therapeutic potential of this protein against COVID-19 is very promising (Table 2).
3.2.3. Inhibition/Blockade of Immune Response Pathways
Based on autoimmune diseases and other infections experienced [174–176], several studies focused on inhibitors or blockers of cellular immune response pathways had been directed to therapeutic targets for COVID-19 treatment, aiming at diminishing the hyperinflammation process led by viral infection. Some authors had mentioned the use of TNF-α inhibitors for COVID-19 treatment [175, 176]. McDermott et al. in 2016 used Kepi, an inhibitory subunit of the protein phosphatase 1 complex (PP1), which was managed in mice models infected with SARS-CoV to reduce signaling of inflammatory cytokines such as TNF-α [176].
Twelve commercial compounds, including anti-inflammatory drugs, immunomodulators, IFNs, and selected antiviral agents were used by Barnard et al. in 2006 to study their effects on SARS-CoV infection in in vitro cell cultures, as well as in Balb/c mice models. The results showed that only hybrid IFN-α B/D was effective in inhibiting SARS-CoV replication in Vero cells, by in vitro assays, and could reduce SARS-CoV replication in the lungs of infected mice. Nevertheless, the authors raised the concern that infected mice lost significant amounts of weight when treated with IFN. Considering that IFN may contribute to the tissue degradative effects of TNF-α already induced by SARS-CoV infection, the problem is to polarize the mouse immune system to a Th2 inflammatory response, with undesired consequences to patient recovery from SARS-CoV infection when treated with an IFN. Thus, more information regarding IFN-α B/D treatment for SARS is necessary. Consequently, its use is not yet recommended for COVID-19 treatment [177].
Monocytes (CD14+) require the chemokine receptor CCR2 to exit the bone marrow driven to inflamed tissues, where they accumulate as macrophages. Since this receptor can be blocked, COVID-19 can benefit from reducing the accumulation of macrophages in inflamed lung tissue. Adding to that, the oxidative stress caused by the virus in macrophages could be reduced using an IRAK4 inhibitor, which could influence the molecular mechanisms of TLR7 activation as well as IL-1β signaling [140] (Table 2).
3.2.4. Anticoagulants and Their Effects on Immune Events
A study using a clinically approved PAR-1 antagonist—SCH530348—demonstrated that it was able to reduce levels of proinflammatory cytokines, neutrophilic lung inflammation, and alveolar leakage during bacterial pneumonia in a murine model [178]. In another study, it was observed that the activation of PAR-1 contributed to the pathogenesis and worsened survival in mice challenged with the influenza virus (IAV), while treatment using an antagonist of this receptor—SCH79797—was able to protect the mice from IAV infection [179]. These reports, thus, reinforce the theory that inhibition of PAR receptor activation may be an interesting approach for the treatment of microthrombosis and other harmful pathological effects associated with the disseminated intravascular coagulation observed in COVID-19. Nevertheless, the PAR-1 antagonist has any disclosure with COVID-19 yet (Table 2).
3.2.5. Immunotherapeutic Approaches with Similarities to Other Immunopathological Diseases
Chong et al. in 2020 suggest the use of Bruton's tyrosine kinase inhibitor (BTKi), a major kinase in the B-cell receptor signaling pathway, mediating B-cell expansion and apoptosis and used in the management of cancer related to B-cell malignancies, for COVID-19 treatment. The authors assumed that BTKi may present pros and cons for patients with cancer affected by COVID-19 because the product plays an active role in macrophage polarization, downstream of classic M1 and M2 polarizing stimuli, and mitigates the inflammatory response in COVID-19. On the other hand, it offers a risk to secondary infections and impairment of humoral immunity. Given these findings, the authors considered the use of BTKi for the treatment of COVID-19 in patients with B-cell malignancies [180, 181].
Based on previous studies with influenza and lipopolysaccharide antigens, Thomas et al. in 2020 published a hypothesis which revealed that a low molecular weight fraction of commercial human serum albumin (LMWF5A), a novel biological development for osteoarthritis, was able to induce in vitro immunomodulatory effects, reducing the cytokine storm and restoring homeostasis to the immune response, attenuating the hyperinflammation, suggesting its positive effects to treat COVID-19 [182] (Table 2).
4. Conclusion
Severe acute respiratory syndrome 2 coronavirus (SARS-CoV-2) has been the new challenge for social, economic, and public health systems worldwide. Since March 2020, the World Health Organization (WHO) and other medical institutes have created, invested, and motivated programs and clinical trials to move forward as fast as possible, to provide any results regarding therapeutic possibilities for COVID-19 (https://www.who.int/dg/speeches/detail/who-director-general-s-opening-remarks-at-the-media-briefing-on-COVID-19---11-march-2020). Despite the race for advances in science and new breakthroughs, there is no efficient current therapy for COVID-19. The immunotherapies described here were also administered in combined treatments carried out in human clinical trials. Vaccine development is underway, but tests are needed to guarantee the safety and efficacy and avoid new waves of contamination in the future.
Here, we were able to summarize important findings in clinical trials and nonclinical studies considering the immunological approaches that could lead to the improvements of COVID-19 therapies, aiming at regulating the cellular immune dysfunction caused by this viral infection with high mortality rates (Figure 4). It is noteworthy to point out that knowledge about the disease is growing daily, with published and unpublished manuscripts (preprint platforms) showing an effort to elucidate mechanisms and understand the pathogenesis as well as propose novel therapeutic targets and strategies. It is also important to emphasize that management of COVID-19 treatment, either immunotherapeutics or not, should be performed within ethical rules and caution, following WHO guidelines and medical practice guidelines, even for in vitro and animal in vivo purposes. Finally, those scientific findings presented here will be helpful in offering substantial information for physicians, clinicians, and researchers to speed up the process towards the achievement of potential breakthroughs in the near future.
Figure 4.
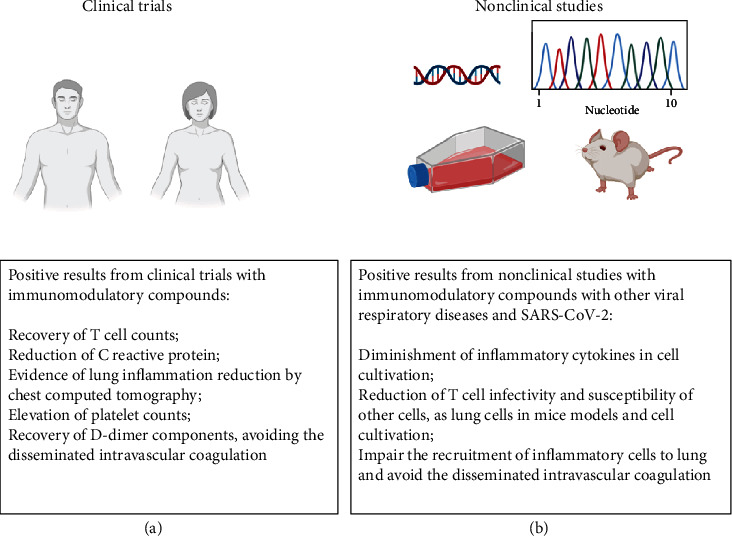
Good results of immunotherapies on clinical trials and nonclinical studies in the immune response against COVID-19. (a) Recent and preliminary positive findings with clinical trials using immunotherapy for COVID-19 treatment are summarized focusing on immune event alterations and recovery, as well as in nonclinical studies (b) for immunotherapeutic interventions using bioinformatics tools, cell cultivation, and animal models; they are also based on past information with other related viral respiratory diseases and autoimmune disorders. Created with BioRender.com.
Data Availability
No data were used to support this study. The figures and table data used to support the findings of this study are included within the article.
Conflicts of Interest
The authors declared no conflict of interests.
References
- 1.Kumar M., Taki K., Gahlot R., Sharma A., Dhangar K. A chronicle of SARS-CoV-2: part-I—epidemiology, diagnosis, prognosis, transmission and treatment. Science of The Total Environment. 2020;734, article 139278 doi: 10.1016/j.scitotenv.2020.139278. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Wang Z., Yang B., Li Q., Wen L., Zhang R. Clinical features of 69 cases with coronavirus disease 2019 in Wuhan, China. Clinical Infectious Diseases. 2020;71(15):769–777. doi: 10.1093/cid/ciaa272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Abduljalil J. M., Abduljalil B. M. Epidemiology, genome, and clinical features of the pandemic SARS-CoV-2: a recent view. New Microbes and New Infections. 2020;35, article 100672 doi: 10.1016/j.nmni.2020.100672. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.WHO-World Health Organization. Coronavirus disease (COVID-19), Situation Report - 182, 20 July 2020. https://www.who.int/docs/default-source/coronaviruse/situation-reports/20200720-covid-19-sitrep-182.pdf?sfvrsn=60aabc5c_2.
- 5.Greenberg S. Update on human rhinovirus and coronavirus infections. Seminars in Respiratory and Critical Care Medicine. 2016;37(4):555–571. doi: 10.1055/s-0036-1584797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Wang D., Hu B., Hu C., et al. Clinical characteristics of 138 hospitalized patients with 2019 novel coronavirus-infected pneumonia in Wuhan, China. JAMA. 2020;323(11):1061–1069. doi: 10.1001/jama.2020.1585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Yang Y., Peng F., Wang R., et al. The deadly coronaviruses: the 2003 SARS pandemic and the 2020 novel coronavirus epidemic in China. Journal of Autoimmunity. 2020;109, article 102434 doi: 10.1016/j.jaut.2020.102434. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Huang A. T., Garcia-Carreras B., Hitchings M. D. T., et al. A systematic review of antibody mediated immunity to coronaviruses: antibody kinetics, correlates of protection, and association of antibody responses with severity of disease. Infectious Diseases. 2020 doi: 10.1101/2020.04.14.20065771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Kong W., Wang Y., Hu J., Chughtai A., Pu H. Comparison of clinical and epidemiological characteristics of asymptomatic and symptomatic SARS-CoV-2 infection: a multi-center study in Sichuan Province, China. Travel Medicine and Infectious Disease. 2020;37, article 101754 doi: 10.1016/j.tmaid.2020.101754. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Qiu J. Covert coronavirus infections could be seeding new outbreaks. Nature. 2020;20 doi: 10.1038/d41586-020-00822-x. [DOI] [PubMed] [Google Scholar]
- 11.WHO EMRO. Transmission of COVID-19 by asymptomatic cases. 2020. http://www.emro.who.int/health-topics/corona-virus/transmission-of-covid-19-by-asymptomatic-cases.html.
- 12.Li Q., Tang B., Bragazzi N. L., Xiao Y., Wu J. Modeling the impact of mass influenza vaccination and public health interventions on COVID-19 epidemics with limited detection capability. Mathematical Biosciences. 2020;325, article 108378 doi: 10.1016/j.mbs.2020.108378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Prompetchara E., Ketloy C., Palaga T. Immune responses in COVID-19 and potential vaccines: lessons learned from SARS and MERS epidemic. Asian Pacific Journal of Allergy and Immunology. 2020;38(1):1–9. doi: 10.12932/ap-200220-0772. [DOI] [PubMed] [Google Scholar]
- 14.He R., Lu Z., Zhang L., et al. The clinical course and its correlated immune status in COVID-19 pneumonia. Journal of Clinical Virology. 2020;127, article 104361 doi: 10.1016/j.jcv.2020.104361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Gong L. L., Zhao B. B., Fan W. F., et al. Correlations of IFN-γ-inducible protein-10 with the risk of chronic hepatitis B and the efficacy of interferon therapy in Asians. International Journal of Clinical and Experimental Pathology. 2015;8(7):8367–8375. [PMC free article] [PubMed] [Google Scholar]
- 16.Henry B. M., de Oliveira M. H. S., Benoit S., Plebani M., Lippi G. Hematologic, biochemical and immune biomarker abnormalities associated with severe illness and mortality in coronavirus disease 2019 (COVID-19): a meta-analysis. Clinical Chemistry and Laboratory Medicine. 2020;58(7):1021–1028. doi: 10.1515/cclm-2020-0369. [DOI] [PubMed] [Google Scholar]
- 17.Grifoni A., Weiskopf D., Ramirez S. I., et al. Targets of T cell responses to SARS-CoV-2 coronavirus in humans with COVID-19 disease and unexposed individuals. Cell. 2020;181(7):1489–1501.e15. doi: 10.1016/j.cell.2020.05.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Altmann D. M., Douek D. C., Boyton R. J. What policy makers need to know about COVID-19 protective immunity. The Lancet. 2020;395(10236):1527–1529. doi: 10.1016/S0140-6736(20)30985-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Popovich N., Sanger-Katz M. The World Is Still Far From Herd Immunity for Coronavirus. The New York Times; 2020. https://www.nytimes.com/interactive/2020/05/28/upshot/coronavirus-herd-immunity.html. [Google Scholar]
- 20.Liu X., Wang J., Xu X., Liao G., Chen Y., Hu C.-H. Patterns of IgG and IgM antibody response in COVID-19 patients. Emerging Microbes & Infections. 2020;9(1):1269–1274. doi: 10.1080/22221751.2020.1773324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Ma H., Zeng W., He H., et al. Serum IgA, IgM, and IgG responses in COVID-19. Cellular & Molecular Immunology. 2020;17(7):773–775. doi: 10.1038/s41423-020-0474-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Long Q.-X., Liu B.-Z., Deng H.-J., et al. Antibody responses to SARS-CoV-2 in patients with COVID-19. Nature Medicine. 2020;26(6):845–848. doi: 10.1038/s41591-020-0897-1. [DOI] [PubMed] [Google Scholar]
- 23.Crotty S. T follicular helper cell biology: a decade of discovery and diseases. Immunity. 2019;50(5):1132–1148. doi: 10.1016/j.immuni.2019.04.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Vinuesa C. G., Linterman M. A., Yu D., MacLennan I. C. Follicular helper T cells. Annual Review of Immunology. 2016;34(1):335–368. doi: 10.1146/annurev-immunol-041015-055605. [DOI] [PubMed] [Google Scholar]
- 25.Guan W.-j., Ni Z.-y., Hu Y., et al. Clinical characteristics of coronavirus disease 2019 in China. The New England Journal of Medicine. 2020;382(18):1708–1720. doi: 10.1056/NEJMoa2002032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Long Q.-X., Tang X.-J., Shi Q.-L., et al. Clinical and immunological assessment of asymptomatic SARS-CoV-2 infections. Nature Medicine. 2020;26(8):1200–1204. doi: 10.1038/s41591-020-0965-6. [DOI] [PubMed] [Google Scholar]
- 27.Buja L. M., Wolf D. A., Zhao B., et al. The emerging spectrum of cardiopulmonary pathology of the coronavirus disease 2019 (COVID-19): report of 3 autopsies from Houston, Texas, and review of autopsy findings from other United States cities. Cardiovascular Pathology. 2020;48, article 107233 doi: 10.1016/j.carpath.2020.107233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Xu X., Chang X. N., Pan H. X., et al. Pathological changes of the spleen in ten patients with coronavirus disease 2019(COVID-19) by postmortem needle autopsy. Zhonghua Bing Li Xue Za Zhi. 2020;49(6):576–582. doi: 10.3760/cma.j.cn112151-20200401-00278. [DOI] [PubMed] [Google Scholar]
- 29.Tay M. Z., Poh C. M., Rénia L., MacAry P. A., Ng L. F. P. The trinity of COVID-19: immunity, inflammation and intervention. Nature Reviews Immunology. 2020;20(6):363–374. doi: 10.1038/s41577-020-0311-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Vabret N., Britton G. J., Gruber C., et al. Immunology of COVID-19: current state of the science. Immunity. 2020;52(6):910–941. doi: 10.1016/j.immuni.2020.05.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Cao W.-C., Liu W., Zhang P.-H., Zhang F., Richardus J. H. Disappearance of antibodies to SARS-associated coronavirus after recovery. The New England Journal of Medicine. 2007;357(11):1162–1163. doi: 10.1056/NEJMc070348. [DOI] [PubMed] [Google Scholar]
- 32.Traggiai E., Becker S., Subbarao K., et al. An efficient method to make human monoclonal antibodies from memory B cells: potent neutralization of SARS coronavirus. Nature Medicine. 2004;10(8):871–875. doi: 10.1038/nm1080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Guo L., Ren L., Yang S., et al. Profiling early humoral response to diagnose novel coronavirus disease (COVID-19) Clinical Infectious Diseases. 2020;71(15):778–785. doi: 10.1093/cid/ciaa310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Ju B., Zhang Q., Ge X., et al. Potent human neutralizing antibodies elicited by SARS-CoV-2 infection. Immunology. 2020 doi: 10.1101/2020.03.21.990770. [DOI] [PubMed] [Google Scholar]
- 35.Robbiani D. F., Gaebler C., Muecksch F., et al. Convergent antibody responses to SARS-CoV-2 in convalescent individuals. Nature. 2020;584(7821):437–442. doi: 10.1038/s41586-020-2456-9. July 2020, http://www.nature.com/articles/s41586-020-2456-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Tian X., Li C., Huang A., et al. Potent binding of 2019 novel coronavirus spike protein by a SARS coronavirus-specific human monoclonal antibody. Emerging Microbes & Infections. 2020;9(1):382–385. doi: 10.1080/22221751.2020.1729069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Zhao J., Yuan Q., Wang H., et al. Antibody responses to SARS-CoV-2 in patients with novel coronavirus disease 2019. Clinical Infectious Diseases. 2020;71:2027–2034. doi: 10.1093/cid/ciaa344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Tan W., Lu Y., Zhang J., et al. Viral kinetics and antibody responses in patients with COVID-19. Infectious Diseases (except HIV/AIDS) 2020 doi: 10.1101/2020.03.24.20042382. [DOI] [Google Scholar]
- 39.Wu F., Wang A., Liu M., et al. Neutralizing antibody responses to SARS-CoV-2 in a COVID-19 recovered patient cohort and their implications. Infectious Diseases (except HIV/AIDS) 2020 doi: 10.1101/2020.03.30.20047365. [DOI] [Google Scholar]
- 40.Li X. Y., du B., Wang Y. S., et al. The keypoints in treatment of the critical coronavirus disease 2019 patient(2) Zhonghua Jie He He Hu Xi Za Zhi. 2020;43(4):277–281. doi: 10.3760/cma.j.cn112147-20200224-00159. [DOI] [PubMed] [Google Scholar]
- 41.Zhang L., Zhang F., Yu W., et al. Antibody responses against SARS coronavirus are correlated with disease outcome of infected individuals. Journal of Medical Virology. 2006;78(1):1–8. doi: 10.1002/jmv.20499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Quinlan B. D., Mou H., Zhang L., et al. The SARS-CoV-2 receptor-binding domain elicits a potent neutralizing response without antibody-dependent enhancement. Microbiology. 2020 doi: 10.1101/2020.04.10.036418. [DOI] [Google Scholar]
- 43.Gao Q., Bao L., Mao H., et al. Rapid development of an inactivated vaccine for SARS-CoV-2. Microbiology. 2020 doi: 10.1101/2020.04.17.046375. [DOI] [Google Scholar]
- 44.Juno J. A., Tan H.-X., Lee W. S., et al. Humoral and circulating follicular helper T cell responses in recovered patients with COVID-19. Nature Medicine. 2020;26(9):1428–1434. doi: 10.1038/s41591-020-0995-0. [DOI] [PubMed] [Google Scholar]
- 45.Zeng Z., Xu L., Xie X.‐. Y., et al. Pulmonary pathology of early-phase COVID-19 pneumonia in a patient with a benign lung lesion. Histopathology. 2020;77(5):823–831. doi: 10.1111/his.14138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Coperchini F., Chiovato L., Croce L., Magri F., Rotondi M. The cytokine storm in COVID-19: an overview of the involvement of the chemokine/chemokine-receptor system. Cytokine & Growth Factor Reviews. 2020;53:25–32. doi: 10.1016/j.cytogfr.2020.05.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Xu Z., Shi L., Wang Y., et al. Pathological findings of COVID-19 associated with acute respiratory distress syndrome. The Lancet Respiratory Medicine. 2020;8(4):420–422. doi: 10.1016/S2213-2600(20)30076-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.du Y., Tu L., Zhu P., et al. Clinical features of 85 fatal cases of COVID-19 from Wuhan. A retrospective observational study. American Journal of Respiratory and Critical Care Medicine. 2020;201(11):1372–1379. doi: 10.1164/rccm.202003-0543OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Walls A. C., Park Y.-J., Tortorici M. A., Wall A., McGuire A. T., Veesler D. Structure, function, and antigenicity of the SARS-CoV-2 spike glycoprotein. Cell. 2020;181(2):281–292.e6. doi: 10.1016/j.cell.2020.02.058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Wang X., Xu W., Hu G., et al. Retraction Note to: SARS-CoV-2 infects T lymphocytes through its spike protein-mediated membrane fusion. Cellular & Molecular Immunology. 2020;17(8) doi: 10.1038/s41423-020-0498-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Catanzaro M., Fagiani F., Racchi M., Corsini E., Govoni S., Lanni C. Immune response in COVID-19: addressing a pharmacological challenge by targeting pathways triggered by SARS-CoV-2. Signal Transduction and Targeted Therapy. 2020;5(1) doi: 10.1038/s41392-020-0191-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Wang F., Nie J., Wang H., et al. Characteristics of peripheral lymphocyte subset alteration in COVID-19 pneumonia. The Journal of Infectious Diseases. 2020;221(11):1762–1769. doi: 10.1093/infdis/jiaa150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Samuel C. E. Antiviral actions of interferons. Clinical Microbiology Reviews. 2001;14(4):778–809. doi: 10.1128/CMR.14.4.778-809.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Knoops K., Kikkert M., Worm S. H. E., et al. SARS-coronavirus replication is supported by a reticulovesicular network of modified endoplasmic reticulum. PLoS Biology. 2008;6(9, article e226) doi: 10.1371/journal.pbio.0060226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Channappanavar R., Fehr A. R., Vijay R., et al. Dysregulated type I interferon and inflammatory monocyte-macrophage responses cause lethal pneumonia in SARS-CoV-infected mice. Cell Host & Microbe. 2016;19(2):181–193. doi: 10.1016/j.chom.2016.01.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Mantlo E., Bukreyeva N., Maruyama J., Paessler S., Huang C. Antiviral activities of type I interferons to SARS-CoV-2 infection. Antiviral Research. 2020;179, article 104811 doi: 10.1016/j.antiviral.2020.104811. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Liu Y., Yang Y., Zhang C., et al. Clinical and biochemical indexes from 2019-nCoV infected patients linked to viral loads and lung injury. Science China Life Sciences. 2020;63(3, article 1643):364–374. doi: 10.1007/s11427-020-1643-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Freundt E. C., Yu L., Park E., Lenardo M. J., Xu X.-N. Molecular determinants for subcellular localization of the severe acute respiratory syndrome coronavirus open reading frame 3b protein. Journal of Virology. 2009;83(13):6631–6640. doi: 10.1128/JVI.00367-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Konno Y., Kimura I., Uriu K., et al. SARS-CoV-2 ORF3b is a potent interferon antagonist whose activity is further increased by a naturally occurring elongation variant. 2020;2020:11. doi: 10.1101/2020.05.11.088179.088179 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Sun L., Xing Y., Chen X., et al. Coronavirus papain-like proteases negatively regulate antiviral innate immune response through disruption of STING-mediated signaling. PLoS One. 2012;7(2, article e30802) doi: 10.1371/journal.pone.0030802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Braun J., Loyal L., Frentsch M., et al. Presence of SARS-CoV-2 Reactive T Cells in COVID-19 Patients and Healthy Donors. medRxiv; 2020. [DOI] [Google Scholar]
- 62.Thevarajan I., Nguyen T. H. O., Koutsakos M., et al. Breadth of concomitant immune responses prior to patient recovery: a case report of non-severe COVID-19. Nature Medicine. 2020;26(4):453–455. doi: 10.1038/s41591-020-0819-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Melgaço J. G., Azamor T., Ano Bom A. P. D. Protective immunity after COVID-19 has been questioned: what can we do without SARS-CoV-2-IgG detection? Cellular Immunology. 2020;353, article 104114 doi: 10.1016/j.cellimm.2020.104114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.CRICS TRIGGERSEP Group (Clinical Research in Intensive Care and Sepsis Trial Group for Global Evaluation and Research in Sepsis), Helms J., Tacquard C., et al. High risk of thrombosis in patients with severe SARS-CoV-2 infection: a multicenter prospective cohort study. Intensive Care Medicine. 2020;46(6, article 6062):1089–1098. doi: 10.1007/s00134-020-06062-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Klok F. A., Kruip M. J. H. A., van der Meer N. J. M., et al. Incidence of thrombotic complications in critically ill ICU patients with COVID-19. Thrombosis Research. 2020;191:145–147. doi: 10.1016/j.thromres.2020.04.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Thachil J., Cushman M., Srivastava A. A proposal for staging COVID-19 coagulopathy. Research and Practice in Thrombosis and Haemostasis. 2020;4(5):731–736. doi: 10.1002/rth2.12372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Tang N., Li D., Wang X., Sun Z. Abnormal coagulation parameters are associated with poor prognosis in patients with novel coronavirus pneumonia. Journal of Thrombosis and Haemostasis. 2020;18(4):844–847. doi: 10.1111/jth.14768. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Zhou F., Yu T., du R., et al. Clinical course and risk factors for mortality of adult inpatients with COVID-19 in Wuhan, China: a retrospective cohort study. The Lancet. 2020;395(10229):1054–1062. doi: 10.1016/s0140-6736(20)30566-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Levi M., Thachil J., Iba T., Levy J. H. Coagulation abnormalities and thrombosis in patients with COVID-19. The Lancet Haematology. 2020;7(6):e438–e440. doi: 10.1016/S2352-3026(20)30145-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Iba T., Levy J. H., Connors J. M., Warkentin T. E., Thachil J., Levi M. The unique characteristics of COVID-19 coagulopathy. Critical Care. 2020;24(1) doi: 10.1186/s13054-020-03077-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Terpos E., Ntanasis-Stathopoulos I., Elalamy I., et al. Hematological findings and complications of COVID-19. American Journal of Hematology. 2020;95(7):834–847. doi: 10.1002/ajh.25829. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Tang N., Bai H., Chen X., Gong J., Li D., Sun Z. Anticoagulant treatment is associated with decreased mortality in severe coronavirus disease 2019 patients with coagulopathy. Journal of Thrombosis and Haemostasis. 2020;18(5):1094–1099. doi: 10.1111/jth.14817. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Paranjpe I., Fuster V., Lala A., et al. Association of treatment dose anticoagulation with in-hospital survival among hospitalized patients with COVID-19. Journal of the American College of Cardiology. 2020;76(1):122–124. doi: 10.1016/j.jacc.2020.05.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Levi M., van der Poll T. Inflammation and coagulation. Critical Care Medicine. 2010;38(2 Supplement):S26–S34. doi: 10.1097/CCM.0b013e3181c98d21. [DOI] [PubMed] [Google Scholar]
- 75.Weiss P., Murdoch D. R. Clinical course and mortality risk of severe COVID-19. The Lancet. 2020;395(10229):1014–1015. doi: 10.1016/S0140-6736(20)30633-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Zhou Z., Ren L., Zhang L., et al. Overly exuberant innate immune response to SARS-CoV-2 infection. SSRN Journal. 2020 https://www.ssrn.com/abstract=3551623. [Google Scholar]
- 77.Zhou Y., Fu B., Zheng X., et al. Pathogenic T-cells and inflammatory monocytes incite inflammatory storms in severe COVID-19 patients. National Science Review. 2020;7(6):998–1002. doi: 10.1093/nsr/nwaa041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Schechter M. E., Andrade B. B., He T., et al. Inflammatory monocytes expressing tissue factor drive SIV and HIV coagulopathy. Science Translational Medicine. 2017;9(405) doi: 10.1126/scitranslmed.aam5441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Neves-Souza P. C., Azeredo E. L., Zagne S. M., et al. Inducible nitric oxide synthase (iNOS) expression in monocytes during acute dengue fever in patients and during in vitro infection. BMC Infectious Diseases. 2005;5(1) doi: 10.1186/1471-2334-5-64. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Simmons J., Pittet J.-F. The coagulopathy of acute sepsis. Current Opinion in Anaesthesiology. 2015;28(2):227–236. doi: 10.1097/ACO.0000000000000163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Antoniak S., Owens A. P., Baunacke M., et al. PAR-1 contributes to the innate immune response during viral infection. Journal of Clinical Investigation. 2013;123(3):1310–1322. doi: 10.1172/JCI66125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Soh U. J. K., Trejo J. Activated protein C promotes protease-activated receptor-1 cytoprotective signaling through β-arrestin and dishevelled-2 scaffolds. Proceedings of the National Academy of Sciences. 2011;108(50):E1372–E1380. doi: 10.1073/pnas.1112482108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Jose R. J., Manuel A. COVID-19 cytokine storm: the interplay between inflammation and coagulation. The Lancet Respiratory Medicine. 2020;8(6):e46–e47. doi: 10.1016/S2213-2600(20)30216-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Mazzeffi M., Chow J. H., Amoroso A., Tanaka K. Revisiting the protein C pathway: an opportunity for adjunctive intervention in COVID-19? Anesthesia & Analgesia. 2020;131(3):690–693. doi: 10.1213/ANE.0000000000005059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Semple J. W., Italiano J. E., Freedman J. Platelets and the immune continuum. Nature Reviews Immunology. 2011;11(4):264–274. doi: 10.1038/nri2956. [DOI] [PubMed] [Google Scholar]
- 86.Rondina M. T., Zimmerman G. A. Platelets. Elsevier; 2019. The role of platelets in inflammation; pp. 505–522. [DOI] [Google Scholar]
- 87.Azeredo E. L., Monteiro R. Q., de-Oliveira Pinto L. M. Thrombocytopenia in dengue: interrelationship between virus and the imbalance between coagulation and fibrinolysis and inflammatory mediators. Mediators of Inflammation. 2015;2015:16. doi: 10.1155/2015/313842.313842 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Lefrançais E., Ortiz-Muñoz G., Caudrillier A., et al. The lung is a site of platelet biogenesis and a reservoir for haematopoietic progenitors. Nature. 2017;544(7648):105–109. doi: 10.1038/nature21706. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Yang X., Yang Q., Wang Y., et al. Thrombocytopenia and its association with mortality in patients with COVID-19. Journal of Thrombosis and Haemostasis. 2020;18(6):1469–1472. doi: 10.1111/jth.14848. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Liu Y., Sun W., Guo Y., et al. Association between platelet parameters and mortality in coronavirus disease 2019: retrospective cohort study. Platelets. 2020;31(4):490–496. doi: 10.1080/09537104.2020.1754383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Bone R. C., Francis P. B., Pierce A. K. Intravascular coagulation associated with the adult respiratory distress syndrome. The American Journal of Medicine. 1976;61(5):585–589. doi: 10.1016/0002-9343(76)90135-2. [DOI] [PubMed] [Google Scholar]
- 92.Peiris J. S. M., Yuen K. Y., Osterhaus A. D. M. E., Stöhr K. The severe acute respiratory syndrome. New England Journal of Medicine. 2003;349(25):2431–2441. doi: 10.1056/NEJMra032498. [DOI] [PubMed] [Google Scholar]
- 93.Weyrich A. S., Lindemann S., Zimmerman G. A. The evolving role of platelets in inflammation. Journal of Thrombosis and Haemostasis. 2003;1(9):1897–1905. doi: 10.1046/j.1538-7836.2003.00304.x. [DOI] [PubMed] [Google Scholar]
- 94.Manne B. K., Denorme F., Middleton E. A., et al. Platelet gene expression and function in patients with COVID-19. Blood. 2020;136(11):1317–1329. doi: 10.1182/blood.2020007214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Hottz E. D., Lopes J. F., Freitas C., et al. Platelets mediate increased endothelium permeability in dengue through NLRP3-inflammasome activation. Blood. 2013;122(20):3405–3414. doi: 10.1182/blood-2013-05-504449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Barros T. A., Batista D. O., Torrentes de Carvalho A., et al. Different aspects of platelet evaluation in dengue: measurement of circulating mediators, ability to interact with the virus, the degree of activation and quantification of intraplatelet protein content. Virus Research. 2019;260:163–172. doi: 10.1016/j.virusres.2018.09.013. [DOI] [PubMed] [Google Scholar]
- 97.Barros T. A., de-Oliveira-Pinto L. M. Thrombocytopenia. IntechOpen; 2018. A view of platelets in dengue. [DOI] [Google Scholar]
- 98.Falati S., Liu Q., Gross P., et al. Accumulation of tissue factor into developing thrombi in vivo is dependent upon microparticle P-selectin glycoprotein ligand 1 and platelet P-selectin. Journal of Experimental Medicine. 2003;197(11):1585–1598. doi: 10.1084/jem.20021868. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Zamora C., Cantó E., Nieto J. C., et al. Functional consequences of platelet binding to T lymphocytes in inflammation. Journal of Leukocyte Biology. 2013;94(3):521–529. doi: 10.1189/jlb.0213074. [DOI] [PubMed] [Google Scholar]
- 100.Chapman L. M., Aggrey A. A., Field D. J., et al. Platelets present antigen in the context of MHC class I. The Journal of Immunology. 2012;189(2):916–923. doi: 10.4049/jimmunol.1200580. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Fuchs T. A., Brill A., Duerschmied D., et al. Extracellular DNA traps promote thrombosis. Proceedings of the National Academy of Sciences. 2010;107(36):15880–15885. doi: 10.1073/pnas.1005743107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Looney M. R. Platelets induce neutrophil extracellular traps in transfusion-related acute lung injury. Blood. 2014;124(21) doi: 10.1182/blood.V124.21.SCI-18.SCI-18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Neri T., Nieri D., Celi A. P-selectin blockade in COVID-19-related ARDS. American Journal of Physiology-Lung Cellular and Molecular Physiology. 2020;318(6):L1237–L1238. doi: 10.1152/ajplung.00202.2020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Thachil J. What do monitoring platelet counts in COVID-19 teach us? Journal of Thrombosis and Haemostasis. 2020;18(8):2071–2072. doi: 10.1111/jth.14879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.du Y., Tu L., Zhu P., et al. Clinical features of 85 fatal cases of COVID-19 from Wuhan. A retrospective observational study. American Journal of Respiratory and Critical Care Medicine. 2020;201(11):1372–1379. doi: 10.1164/rccm.202003-0543OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.di Mascio D., Khalil A., Saccone G., et al. Outcome of coronavirus spectrum infections (SARS, MERS, COVID-19) during pregnancy: a systematic review and meta-analysis. American Journal of Obstetrics & Gynecology MFM. 2020;2(2, article 100107) doi: 10.1016/j.ajogmf.2020.100107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Liu M., Gao Y., Zhang Y., Shi S., Chen Y., Tian J. The association between severe or dead COVID-19 and autoimmune diseases: a systematic review and meta-analysis. The Journal of Infection. 2020;81(3):e93–e95. doi: 10.1016/j.jinf.2020.05.065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Shang Y., Pan C., Yang X., et al. Management of critically ill patients with COVID-19 in ICU: statement from front-line intensive care experts in Wuhan, China. Annals of Intensive Care. 2020;10(1) doi: 10.1186/s13613-020-00689-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Corman V. M., Rabenau H. F., Adams O., et al. SARS-CoV-2 asymptomatic and symptomatic patients and risk for transfusion transmission. Transfusion. 2020;60(6):1119–1122. doi: 10.1111/trf.15841. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Prete M., Favoino E., Catacchio G., Racanelli V., Perosa F. SARS-CoV-2 inflammatory syndrome. Clinical features and rationale for immunological treatment. International Journal of Molecular Sciences. 2020;21(9) doi: 10.3390/ijms21093377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Zhang J., Xie B., Hashimoto K. Current status of potential therapeutic candidates for the COVID-19 crisis. Brain, Behavior, and Immunity. 2020;87:59–73. doi: 10.1016/j.bbi.2020.04.046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Caly L., Druce J. D., Catton M. G., Jans D. A., Wagstaff K. M. The FDA-approved drug ivermectin inhibits the replication of SARS-CoV-2 in vitro. Antiviral Research. 2020;178, article 104787 doi: 10.1016/j.antiviral.2020.104787. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Sajid M. S., Iqbal Z., Muhammad G., Iqbal M. U. Immunomodulatory effect of various anti-parasitics: a review. Parasitology. 2006;132(3):301–313. doi: 10.1017/S0031182005009108. [DOI] [PubMed] [Google Scholar]
- 114.Rajter J. C., Sherman M., Fatteh N., Vogel F., Sacks J., Rajter J.-J. ICON (Ivermectin in COvid Nineteen) study: use of ivermectin is associated with lower mortality in hospitalized patients with COVID19. 2020. https://www.medrxiv.org/content/10.1101/2020.06.06.20124461v2.abstract. [DOI] [PMC free article] [PubMed]
- 115.Blakley B. R., Rousseaux C. G. Effect of ivermectin on the immune response in mice. American Journal of Veterinary Research. 1991;52(4):593–595. [PubMed] [Google Scholar]
- 116.Sajid M. S., Iqbal Z., Muhammad G., et al. Effect of ivermectin on the cellular and humoral immune responses of rabbits. Life Sciences. 2007;80(21):1966–1970. doi: 10.1016/j.lfs.2007.02.025. [DOI] [PubMed] [Google Scholar]
- 117.Chockalingam A. K., Hamed S., Goodwin D. G., et al. The effect of oseltamivir on the disease progression of lethal influenza a virus infection: plasma cytokine and miRNA responses in a mouse model. Disease Markers. 2016;2016:12. doi: 10.1155/2016/9296457.9296457 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Bird N. L., Olson M. R., Hurt A. C., et al. Oseltamivir prophylaxis reduces inflammation and facilitates establishment of cross-strain protective T cell memory to influenza viruses. PLOS ONE. 2015;10(6, article e0129768) doi: 10.1371/journal.pone.0129768. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Coenen S., van der Velden A. W., Cianci D., et al. Oseltamivir for coronavirus illness: post-hoc exploratory analysis of an open-label, pragmatic, randomised controlled trial in European primary care from 2016 to 2018. British Journal of General Practice. 2020;70(696):e444–e449. doi: 10.3399/bjgp20X711941. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Fintelman-Rodrigues N., Sacramento C. Q., Lima C. R., et al. Atazanavir Inhibits SARS-CoV-2 Replication and Pro-inflammatory Cytokine Production. bioRxiv; 2020. [DOI] [Google Scholar]
- 121.Blanco-Melo D., Nilsson-Payant B. E., Liu W.-C., et al. Imbalanced host response to SARS-CoV-2 drives development of COVID-19. Cell. 2020;181(5):1036–1045.e9. doi: 10.1016/j.cell.2020.04.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Hadjadj J., Yatim N., Barnabei L., et al. Impaired type I interferon activity and exacerbated inflammatory responses in severe Covid-19 patient. Infectious Diseases (except HIV/AIDS) 2020 doi: 10.1101/2020.04.19.20068015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Yale IMPACT Team, Lucas C., Wong P., et al. Longitudinal analyses reveal immunological misfiring in severe COVID-19. Nature. 2020;584(7821, article 2588):463–469. doi: 10.1038/s41586-020-2588-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Zhou Z., Ren L., Zhang L., et al. Heightened innate immune responses in the respiratory tract of COVID-19 patients. Cell Host & Microbe. 2020;27(6):883–890.e2. doi: 10.1016/j.chom.2020.04.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Shalhoub S. Interferon beta-1b for COVID-19. The Lancet. 2020;395(10238):1670–1671. doi: 10.1016/S0140-6736(20)31101-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Wang N., Zhan Y., Zhu L., et al. Retrospective multicenter cohort study shows early interferon therapy is associated with favorable clinical responses in COVID-19 patients. Cell Host & Microbe. 2020;28(3):455–464.e2. doi: 10.1016/j.chom.2020.07.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Traves S., Proud D. Viral-associated exacerbations of asthma and COPD. Current Opinion in Pharmacology. 2007;7(3):252–258. doi: 10.1016/j.coph.2006.11.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Singanayagam A., Glanville N., Girkin J. L., et al. Corticosteroid suppression of antiviral immunity increases bacterial loads and mucus production in COPD exacerbations. Nature Communications. 2018;9(1) doi: 10.1038/s41467-018-04574-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Stern A., Skalsky K., Avni T., et al. Corticosteroids for pneumonia. Cochrane Database of Systematic Reviews. 2017;12(12) doi: 10.1002/14651858.CD007720.pub3.CD007720 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Heming N., Sivanandamoorthy S., Meng P., Bounab R., Annane D. Immune effects of corticosteroids in sepsis. Frontiers in Immunology. 2018;9 doi: 10.3389/fimmu.2018.01736. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Ledford H. Coronavirus breakthrough: dexamethasone is first drug shown to save lives. Nature. 2020;582(7813):469–469. doi: 10.1038/d41586-020-01824-5. [DOI] [PubMed] [Google Scholar]
- 132.Chroboczek T., Lacoste M., Wackenheim C., et al. Beneficial Effect of Corticosteroids in Severe COVID-19 Pneumonia: A Propensity Score Matching Analysis. medRxiv; 2020. [DOI] [Google Scholar]
- 133.Gangopadhyay K. K., Mukherjee J. J., Sinha B., Ghosal S. The Role of Corticosteroids in the Management of Critically Ill Patients with Coronavirus Disease 2019 (COVID-19): A Meta-Analysis. medRxiv; 2020. [DOI] [Google Scholar]
- 134.Wang Y., Jiang W., He Q., et al. Early, Low-Dose and Short-Term Application of Corticosteroid Treatment in Patients with Severe COVID-19 Pneumonia: Single-Center Experience from Wuhan, China. medRxiv; 2020. [DOI] [Google Scholar]
- 135.Lansbury L., Rodrigo C., Leonardi-Bee J., Nguyen-van-Tam J., Lim W. S., Cochrane Acute Respiratory Infections Group Corticosteroids as adjunctive therapy in the treatment of influenza. Cochrane Database of Systematic Reviews. 2019;2(2) doi: 10.1002/14651858.CD010406.pub3.CD010406 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Li H., Yang S., Gu L., et al. Effect of low-to-moderate-dose corticosteroids on mortality of hospitalized adolescents and adults with influenza A(H1N1)pdm09 viral pneumonia. Influenza and Other Respiratory Viruses. 2017;11(4):345–354. doi: 10.1111/irv.12456. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Shih H.-I., Wu C.-J., Tu Y.-F., Chi C.-Y. Fighting COVID-19: a quick review of diagnoses, therapies, and vaccines. Biomedical Journal. 2020;43(4):341–354. doi: 10.1016/j.bj.2020.05.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Xu X., Han M., Li T., et al. Effective treatment of severe COVID-19 patients with tocilizumab. Proceedings of the National Academy of Sciences. 2020;117(20):10970–10975. doi: 10.1073/pnas.2005615117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Wampler Muskardin T. L. Intravenous Anakinra for macrophage activation syndrome may hold lessons for treatment of cytokine storm in the setting of coronavirus disease 2019. ACR Open Rheumatology. 2020;2(5):283–285. doi: 10.1002/acr2.11140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Merad M., Martin J. C. Pathological inflammation in patients with COVID-19: a key role for monocytes and macrophages. Nature Reviews Immunology. 2020;20(6):355–362. doi: 10.1038/s41577-020-0331-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Richardson P., Griffin I., Tucker C., et al. Baricitinib as potential treatment for 2019-nCoV acute respiratory disease. The Lancet. 2020;395(10223):e30–e31. doi: 10.1016/S0140-6736(20)30304-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Felsenstein S., Herbert J. A., McNamara P. S., Hedrich C. M. COVID-19: immunology and treatment options. Clinical Immunology. 2020;215, article 108448 doi: 10.1016/j.clim.2020.108448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Melgaço J. G., Veloso C. E., Pacheco-Moreira L. F., Vitral C. L., Pinto M. A. Complement system as a target for therapies to control liver regeneration/damage in acute liver failure induced by viral hepatitis. Journal of Immunology Research. 2018;2018:9. doi: 10.1155/2018/3917032.3917032 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Gralinski L. E., Sheahan T. P., Morrison T. E., et al. Complement activation contributes to severe acute respiratory syndrome coronavirus pathogenesis. mBio. 2018;9(5, article e01753) doi: 10.1128/mBio.01753-18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Diurno F., Numis F. G., Porta G., et al. Eculizumab treatment in patients with COVID-19: preliminary results from real life ASL Napoli 2 Nord experience. European Review for Medical and Pharmacological Sciences. 2020;24(7):4040–4047. doi: 10.26355/eurrev_202004_20875. [DOI] [PubMed] [Google Scholar]
- 146.Dolinger M. T., Person H., Smith R., et al. Pediatric Crohn disease and multisystem inflammatory syndrome in children (MIS-C) and COVID-19 treated with infliximab. Journal of Pediatric Gastroenterology and Nutrition. 2020;71(2):153–155. doi: 10.1097/MPG.0000000000002809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Wang K., Chen W., Zhou Y.-S., et al. SARS-CoV-2 Invades Host Cells via a Novel Route: CD147-Spike Protein. bioRxiv; 2020. [DOI] [Google Scholar]
- 148.Koch C., Staffler G., Hüttinger R., et al. T cell activation-associated epitopes of CD147 in regulation of the T cell response, and their definition by antibody affinity and antigen density. International Immunology. 1999;11(5):777–786. doi: 10.1093/intimm/11.5.777. [DOI] [PubMed] [Google Scholar]
- 149.Ulrich H., Pillat M. M. CD147 as a target for COVID-19 treatment: suggested effects of azithromycin and stem cell engagement. Stem Cell Reviews and Reports. 2020;16(3):434–440. doi: 10.1007/s12015-020-09976-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Tu Y.-F., Chien C.-S., Yarmishyn A. A., et al. A review of SARS-CoV-2 and the ongoing clinical trials. International Journal of Molecular Sciences. 2020;21(7) doi: 10.3390/ijms21072657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Lippi G., Plebani M., Henry B. M. Thrombocytopenia is associated with severe coronavirus disease 2019 (COVID-19) infections: a meta-analysis. Clinica Chimica Acta. 2020;506:145–148. doi: 10.1016/j.cca.2020.03.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Dolhnikoff M., Duarte-Neto A. N., Almeida Monteiro R. A., et al. Pathological evidence of pulmonary thrombotic phenomena in severe COVID-19. Journal of Thrombosis and Haemostasis. 2020;18(6):1517–1519. doi: 10.1111/jth.14844. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Saghazadeh A., Rezaei N. Towards treatment planning of COVID-19: rationale and hypothesis for the use of multiple immunosuppressive agents: anti-antibodies, immunoglobulins, and corticosteroids. International Immunopharmacology. 2020;84, article 106560 doi: 10.1016/j.intimp.2020.106560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Leng Z., Zhu R., Hou W., et al. Transplantation of ACE2− mesenchymal stem cells improves the outcome of patients with COVID-19 pneumonia. Aging and disease. 2020;11(2):216–228. doi: 10.14336/AD.2020.0228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Bachanova V., Bishop M. R., Dahi P., et al. Chimeric antigen receptor T cell therapy during the COVID-19 pandemic. Biology of Blood and Marrow Transplantation. 2020;26(7):1239–1246. doi: 10.1016/j.bbmt.2020.04.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Per Ljungman, for the European Society for Blood and Marrow Transplantation, Mikulska M., de la Camara R., et al. The challenge of COVID-19 and hematopoietic cell transplantation; EBMT recommendations for management of hematopoietic cell transplant recipients, their donors, and patients undergoing CAR T-cell therapy. Bone Marrow Transplantation. 2020;55:2071–2076. doi: 10.1038/s41409-020-0919-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Zhang L., Liu Y. Potential interventions for novel coronavirus in China: a systematic review. Journal of Medical Virology. 2020;92(5):479–490. doi: 10.1002/jmv.25707. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Meltzer D. O., Best T. J., Zhang H., Vokes T., Arora V., Solway J. Association of Vitamin D Deficiency and Treatment with COVID-19 Incidence. medRxiv; 2020. [DOI] [Google Scholar]
- 159.Parikh R., Wilson C., Weinberg J., Gavin D., Murphy J., Reardon C. C. Inhaled nitric oxide treatment in spontaneously breathing COVID-19 patients. Therapeutic Advances in Respiratory Disease. 2020;14 doi: 10.1177/1753466620933510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Sacramento C. Q., Fintelman-Rodrigues N., Temerozo J. R., et al. The In Vitro Antiviral Activity of the Anti-hepatitis C Virus (HCV) Drugs Daclatasvir and Sofosbuvir against SARS-CoV-2. bioRxiv; 2020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Baker E., Baker H. A structural framework for understanding the multifunctional character of lactoferrin. Biochimie. 2009;91(1):3–10. doi: 10.1016/j.biochi.2008.05.006. [DOI] [PubMed] [Google Scholar]
- 162.Baker E. N., Baker H. M. Lactoferrin. Cellular and Molecular Life Sciences. 2005;62(22):2531–2539. doi: 10.1007/s00018-005-5368-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Serrano G., Kochergina I., Albors A., et al. Liposomal lactoferrin as potential preventative and cure for COVID-19. International Journal of Research in Health Sciences. 2020;8(1):8–15. doi: 10.5530/ijrhs.8.1.3. [DOI] [Google Scholar]
- 164.Lang J., Yang N., Deng J., et al. Inhibition of SARS pseudovirus cell entry by lactoferrin binding to heparan sulfate proteoglycans. PLoS One. 2011;6(8, article e23710) doi: 10.1371/journal.pone.0023710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Carvalho C. A. M., Sousa I. P., Silva J. L., Oliveira A. C., Gonçalves R. B., Gomes A. M. O. Inhibition of Mayaro virus infection by bovine lactoferrin. Virology. 2014;452-453:297–302. doi: 10.1016/j.virol.2014.01.022. [DOI] [PubMed] [Google Scholar]
- 166.Carvalho C. A. M., Casseb S. M. M., Gonçalves R. B., Silva E. V. P., Gomes A. M. O., Vasconcelos P. F. C. Bovine lactoferrin activity against Chikungunya and Zika viruses. Journal of General Virology. 2017;98(7):1749–1754. doi: 10.1099/jgv.0.000849. [DOI] [PubMed] [Google Scholar]
- 167.de Carvalho C. A. M., da Rocha Matos A., Caetano B. C., et al. In Vitro Inhibition of SARS-CoV-2 Infection by Bovine Lactoferrin. bioRxiv; 2020. [DOI] [Google Scholar]
- 168.Superti F., Agamennone M., Pietrantoni A., Ammendolia M. G. Bovine lactoferrin prevents influenza A virus infection by interfering with the fusogenic function of viral hemagglutinin. Viruses. 2019;11(1) doi: 10.3390/v11010051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Kruzel M. L., Zimecki M., Actor J. K. Lactoferrin in a context of inflammation-induced pathology. Frontiers in Immunology. 2017;8, article 1438 doi: 10.3389/fimmu.2017.01438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Kruzel M. L., Actor J. K., Zimecki M., et al. Novel recombinant human lactoferrin: differential activation of oxidative stress related gene expression. Journal of Biotechnology. 2013;168(4):666–675. doi: 10.1016/j.jbiotec.2013.09.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Baker H. M., Baker E. N. Lactoferrin and iron: structural and dynamic aspects of binding and release. Biometals. 2004;17(3):209–216. doi: 10.1023/B:BIOM.0000027694.40260.70. [DOI] [PubMed] [Google Scholar]
- 172.Curran C. S., Bertics P. J. Lactoferrin regulates an axis involving CD11b and CD49d integrins and the chemokines MIP-1α and MCP-1 in GM-CSF-treated human primary eosinophils. Journal of Interferon & Cytokine Research. 2012;32(10):450–461. doi: 10.1089/jir.2011.0111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.Vallabhapurapu S., Karin M. Regulation and function of NF-κB transcription factors in the immune system. Annual Review of Immunology. 2009;27(1):693–733. doi: 10.1146/annurev.immunol.021908.132641. [DOI] [PubMed] [Google Scholar]
- 174.Boechat J. L., Chora I., Delgado L. Imunologia da doença por coronavírus-19 (COVID-19): uma perspetiva para o clínico, nos primeiros 4 meses da emergência do SARS-CoV-2. Medicina Interna. 2020;27 doi: 10.24950/rspmi/COVID19/FMUP/S/2020. [DOI] [Google Scholar]
- 175.Feldmann M., Maini R. N., Woody J. N., et al. Trials of anti-tumour necrosis factor therapy for COVID-19 are urgently needed. The Lancet. 2020;395(10234):1407–1409. doi: 10.1016/S0140-6736(20)30858-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.McDermott J. E., Mitchell H. D., Gralinski L. E., et al. The effect of inhibition of PP1 and TNFα signaling on pathogenesis of SARS coronavirus. BMC Systems Biology. 2016;10(1) doi: 10.1186/s12918-016-0336-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Barnard D. L., Day C. W., Bailey K., et al. Evaluation of immunomodulators, interferons and known in vitro SARS-CoV inhibitors for inhibition of SARS-Cov replication in BALB/c mice. Antiviral Chemistry and Chemotherapy. 2006;17(5):275–284. doi: 10.1177/095632020601700505. [DOI] [PubMed] [Google Scholar]
- 178.José R. J., Williams A. E., Mercer P. F., Sulikowski M. G., Brown J. S., Chambers R. C. Regulation of neutrophilic inflammation by proteinase-activated receptor 1 during bacterial pulmonary infection. The Journal of Immunology. 2015;194(12):6024–6034. doi: 10.4049/jimmunol.1500124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179.Khoufache K., Berri F., Nacken W., et al. PAR1 contributes to influenza A virus pathogenicity in mice. Journal of Clinical Investigation. 2013;123(1):206–214. doi: 10.1172/JCI61667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Chong E. A., Roeker L. E., Shadman M., Davids M. S., Schuster S. J., Mato A. R. BTK inhibitors in cancer patients with COVID-19: “The winner will be the one who controls that chaos” (Napoleon Bonaparte) Clinical Cancer Research. 2020;26(14):3514–3516. doi: 10.1158/1078-0432.CCR-20-1427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181.Lucas F., Woyach J. Inhibiting Bruton’s tyrosine kinase in CLL and other B-cell malignancies. Targeted Oncology. 2019;14(2):125–138. doi: 10.1007/s11523-019-00635-7. [DOI] [PubMed] [Google Scholar]
- 182.Thomas G., Frederick E., Hausburg M., et al. The novel immunomodulatory biologic LMWF5A for pharmacological attenuation of the “cytokine storm” in COVID-19 patients: a hypothesis. Patient Safety in Surgery. 2020;14(1) doi: 10.1186/s13037-020-00248-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 183.Bacil E. D. A., Mazzardo Júnior O., Rech C. R., Legnani R. F. S., Campos W. Physical activity and biological maturation: a systematic review. Revista Paulista de Pediatria. 2015;33(1):114–121. doi: 10.1016/j.rpped.2014.11.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
No data were used to support this study. The figures and table data used to support the findings of this study are included within the article.