

Abstract
The corticostriatal circuitry and its glutamate-GABA interactions play an essential role in regulating neuronal excitability during reward-seeking behavior. However, the contribution of GABAergic interneurons in the corticostriatal circuitry remains unclear. To investigate the role of GABAergic interneurons, we focused on parvalbumin-expressing fast-spiking interneurons (Pv-FSI) in the corticostriatal circuitry using the designer receptors exclusively activated by designer drugs (DREADD) approach in a Pv-Cre mouse model. We hypothesize that Pv-FSI activation elicits changes in cortical glutamate levels and reward-seeking behaviors. To determine molecular and behavioral effects of Pv-FSI, we performed microdialysis and operant conditioning tasks for sucrose and alcohol rewards. In addition, we also examined how alcohol reward itself affects Pv-FSI functioning. Interestingly, our microdialysis results demonstrate that alcohol exposure inhibits Pv-FSI functioning in the medial prefrontal cortex (mPFC) and this consequently can regulate glutamate levels downstream in the nucleus accumbens (NAc). For sucrose reward-seeking behaviors, Pv-FSI activation in the mPFC increases sucrose self-administration whereas it does not promote alcohol seeking. For alcohol rewards, however, Pv-FSI activation in the mPFC results in increased compulsive head entry in operant chambers during devaluation procedures. Overall, our results suggest that not only do Pv-FSI contribute to changes in the cortical microcircuit and reward-seeking behaviors but also that alcohol affects Pv-FSI neurotransmission. Therefore, Pv-FSI has prompted interest in their role in maintaining a balance in neuronal excitation/inhibition and in regulating reward-seeking processes such as compulsivity, all of which are important factors for excessive alcohol seeking.
Keywords: Parvalbumin expressing fast-spiking interneurons (Pv-FSI), microdialysis, mPFC, glutamate, reward-seeking
Introduction
The corticostriatal circuitry, consisting of the medial pre-frontal cortex (mPFC) and the nucleus accumbens (NAc), plays an essential role in coordinating and regulating neuronal excitability during reward-seeking behaviors. Moreover, the mPFC is important for the expression and extinction of cue-mediated behavior and is regulated by cortical neurons, while the NAc is an essential hub integrating cognitive, contextual, and affective information and is mediated by medium-sized spiny neurons (Nam et al. 2011; Barker et al. 2015). Glutamate dysregulation in this circuit plays a role in the reward-seeking behaviors as well as the compulsive behaviors that characterize addiction and its development.
Much research has been done on these brain regions and how glutamate dysregulation is involved in reward-seeking behaviors. However, the contribution of interneurons in glutamate circuitries have not been fully established. Parvalbumin (Pv) interneurons are fast-spiking interneurons (FSI) and they have been implicated in studies on cognition and drug addiction as well (Hu et al. 2014). Although Pv-FSI are comprised of only 2% of the neuronal population in the mPFC and NAc, these smaller neuronal populations modulate the glutamatergic activity of pyramidal neurons in the mPFC (Kim et al. 2016; Sparta et al. 2014; Gonzalez-Burgos & Lewis 2012; Kim et al. 2015) while Pv-FSI activity in the NAc affects GABAergic medium spiny neurons (MSNs) (Lee et al. 2017; Jinno & Kosaka 2004; Wang et al. 2018). As a single Pv-FSI inhibits an estimated number of 100 MSNs (Burguiere et al. 2015), Pv-FSI understandably can have astounding effects on the communication of this circuitry. In fact, activation of Pv-FSI in the mPFC and NAc both result in behavioral changes to reward (Wang et al. 2018).
Therefore, it is required to study how Pv-FSI activation modulates corticostriatal circuitry and how it alters reward-seeking behaviors for natural rewards like sucrose and drug rewards like alcohol. It has been reported that mPFC-specific Pv-FSI activation upon demonstration of cues associated with reward significantly accelerated the extinction of reward-seeking behavior (Sparta et al. 2014), while NAc-specific Pv-FSI activation also influences cocaine and methamphetamine reward properties (Todtenkopf et al. 2004; Wang et al. 2018). Even though alcohol is known to inhibit GABA release from interneurons and induce NMDA glutamate receptor hypo-function in Pv-FSI (Homayoun & Moghaddam 2007), the impact of alcohol on Pv-FSI regulation, however, has not been studied with regards to alcohol-seeking behaviors.
The chemogenetics approach using Designer Receptor Exclusively Activated by Designer Drugs (DREADD) (Roth 2016) in Cre-recombinase mouse models enabled us to test the role of Pv-FSI in the mPFC and NAc on the reward system. In this study, we bilaterally transduced Pv-FSI with the AAV5 virus that carries the hM3Dq designer receptor to induce firing of the interneurons. Pv-FSI were activated in the mPFC or NAc using Cre-dependent DREADD expression in Pv-Cre mice (Madisen et al. 2010). To elucidate the role of Pv-FSI activation during alcohol reward-seeking behaviors, we examined how Pv-FSI activation and alcohol treatment induces neurotransmitter changes using in vivo microdialysis in the target brain regions, which may also explain the molecular mechanism of Pv-FSI and its role in addiction development. Then, we utilized operant conditioning behavioral paradigms to compare reward-seeking responses and compulsivity for both sucrose and alcohol rewards. Our molecular and behavioral work collectively elucidates a novel Pv-FSI microcircuitry as a fine-tuning mechanism in the corticostriatal circuitry controlling reward-seeking behaviors.
Materials and Methods
This study was not pre-registered.
Animals
Pv-Cre mice were purchased from The Jackson Laboratory (Bar Harbor, ME; B6;129P2-Pvalbtm1(cre)Arbr (IMSR Cat# JAX:008069, RRID:IMSR_JAX:008069). Male mice aged 9 weeks at the beginning of the experiments were used. Mice were individually-housed in standard Plexiglas cages under a 12 h light/dark cycle (lights on at 6:00 AM) at a constant temperature (24 ± 0.5°C) and humidity (60 ± 2%) with food and water available ad libitum. The animal care and handling procedures were both approved by the LSUHSC-Shreveport Institutional Animal Care and Use Committees (P-17-038) in accordance with NIH guidelines. A total of 43 mice were used in this study.
Stereotaxic surgery and DREADD expression
Cre-dependent DREADD constructs (AAV-hSyn-DIO-mCherry (RRID:Addgene_50459); AAV-hSyn-DIO-hM3D(Gq)-mCherry (RRID:Addgene_44361); and AAV-hSyn-HA-hM3D(Gq)-IRES-mCitrine (RRID:Addgene_50463)) were purchased from Addgene. For DREADD expression in the target brain regions, stereotaxic injections of AAV viruses were conducted as described previously (Sullivan et al. 2019). 9-weeks old Pv-Cre mice were anesthetized with 2–3% isoflurane gas and placed in a digital stereotaxic alignment system (Model 1900; David Kopf Instruments). Two holes for bilateral injection were drilled into the skull above the target brain region. The injector (33 Ga; Plastics One) was connected to a 10 μl Hamilton syringe (Cat# 65460-05) and the viral infusion was controlled. To infuse the virus, 1 μl of DREADD constructs were bilaterally injected into either the prelimbic region of the medial prefrontal cortex (mPFC) (AP: 1.94 mm; ML: ±0.7 mm; DV: −1.0 mm) or the nucleus accumbens (NAc) (AP: 1.34 mm; ML: ±1.3 mm; DV: −3.5 mm) or at a rate of 0.1 μl/min. Injectors remained in place for an additional 5 min after each infusion. After surgery, mice were monitored daily for signs of pain or discomfort and provided a subcutaneous injection of carprofen (5–10 mg/kg) if needed. To obtain full viral expression, mice were under recovery for 3 weeks. DREADD activation experiments were performed when mice were 12-weeks old by injecting either saline or clozapine-N-oxide dihydrochloride (CNO; 1 mg/kg, dissolved in saline, i.p; product #HB6149; Hello Bio Inc, Princeton, NJ, USA) (Page et al. 2019). The coordinate for viral expression was validated by mCherry expression. There was no significant off target expression that occurred.
Immunofluorescence for c-fos expression
To detect DREADD-induced neural activity in the mPFC, CNO (1 mg/kg) or saline was injected in DREADD-expressing mice (n=3 per group) 30 min prior to transcardial perfusion and brain collection. Mice were anesthetized with 2–3% isoflurane and fixed during transcardial perfusion with phosphate-buffered saline (PBS) and 4% paraformaldehyde in PBS. Brains were removed, post-fixed overnight, and incubated in 30% sucrose at room temperature until they sank. Using a freezing microtome, they were sectioned at 20–30 μm. Immunohistochemistry was performed on free-floating sections. Sections were incubated with primary rabbit antibody (c-fos (9F6) Rabbit; Cell Signaling Technology Cat# 2250, RRID:AB_2247211) in 1:6400 dilution overnight at 4°C followed by an incubation of secondary antibody purchased from ThermoFisher (Molecular Probes Cat# A-11008, RRID:AB_143165) goat anti-rabbit in 1:1000 dilution at room temperature for 1 h. Immunofluorescence was detected on a Nikon SIM-E & A1R confocal microscopy system to examine DREADD viral infusion and c-fos proteins. No blinding was performed. Quantification was achieved using the unbiased stereology method to count the percent of c-fos expression co-expressed in mCherry-expressing PV-FSI cells.
Electrophysiology
Electrophysiology experiments were conducted as described in (Kang et al. 2017). Mice were anesthetized by inhalation of 2–3% isoflurane and the brain was rapidly removed and placed in ice-cold-sucrose-based artificial cerebrospinal fluid (aCSF) containing the following (in mM): 210 sucrose, 2.5 KCl, 1.25 NaH2PO4, 1 MgCl2, 2 CaCl2, 25 NaHCO3, 0.3 l-ascorbate, and 10 glucose, and oxygenated with 95% O2/5% CO2. Coronal (300 μm) slices were cut with a vibrating microtome (S100, Leica Corp., Bannockburn, IL), then transferred to a holding chamber and incubated for 30 min at 34°C and kept for at least 1 h at room temperature (24–25°C) in carbonated (95%O2/5%CO2) standard artificial cerebrospinal fluid (aCSF) containing the following (in mM): 126 NaCl, 2.5 KCl, 1.25 NaH2PO4, 1 MgCl2, 2 CaCl2, 25 NaHCO3, 1 l-ascorbate and 10 glucose. After equilibration, a single slice was transferred to a submersion-type recording chamber and mechanically stabilized with an anchor (Warner Instruments, Hamden, CT). Electrical signals were recorded with an Axon 700B amplifier, a Digidata 1550B A/D converter, Clampex and analyzed with Clampfit 11.0 software (Molecular Devices Co., Union City, CA, USA). Throughout the experiments, the bath was continually perfused with warm (32°C) carbonated aCSF (2.0–2.5 ml/min). Patch pipettes had a resistance of 6–8 MΩ when filled with a solution containing (in mM): 140 K-gluconate, 5 KCl, 2 MgCl2, 10 HEPES, 2 MgATP and 0.2 Na2GTP for current clamping, and 140 Cs-methane sulfonate, 5 KCl, 2 MgCl2, 10 HEPES, 2 MgATP and 0.2 Na2GTP for voltage clamping. Access resistance was monitored throughout the experiments by a step of −10 mV and recordings were discarded if the access resistance changed by more than 20%. The parvalbumin-positive neurons were identified with a red fluorescent protein color.
Bilateral microdialysis and LC-MS/MS
Mice brains were virally infused with DREADD bilaterally in the mPFC. 2 weeks following viral infusion, mice had surgical implantation of two microdialysis guide cannulas. Mice were under anesthesia with isoflurane and placed in a digital stereotaxic alignment system (Model 1900; David Kopf Instruments). First, holes were drilled into the skull above the left mPFC (AP: 1.94 mm, ML: −0.5 mm, DV: −2.0 mm) and right NAc (AP: 1.34 mm, ML: 1.3 mm, DV: −3.5 mm). Metal screws were fixed onto the posterior part of the skull to hold the cement. Afterwards, the probe (DZ D-I-2-01, Eicom) was inserted into the mPFC and NAc locations. After surgery, we provided a subcutaneous injection of analgesic, carprofen (5–10 mg/kg). Mice were given 1 week to recover from microdialysis stereotaxic surgery and were monitored daily to ensure a healthy recovery.
For the microdialysis measurements (n=4 per group), the probe was equilibrated with artificial CSF containing the following (in mM): 155 NaCl, 3.5 KCl, 1 CaCl2, 1 MgCl2, and 2 NaH2PO4, pH 7.4, and the perfusion was delivered by a 500 μl SGE syringe filled with ddH2O controlled by a Harvard microinjection pump. All microdialysates were collected at a continuous flow rate of 2 μl/min for every 7 min. Real-time measurements of the neurotransmitter quantification were at 7-minute intervals. After a stabilization period of 2 h, baseline microdialysates were measured for 1 h. After the 1 h baseline period, treatment of either saline or alcohol (EtOH;1.5 g/kg, i.p) was injected. Then, CNO (1 mg/kg, i.p) was injected an hour later. Microdialysates were collected for an hour each after treatment and CNO injections. Each fraction of dialysate (14 μl) were deidentified and analyzed using liquid chromatography-mass spectrometry (LC-MS/MS; UPLC Aquity/XEVO TQ MS; Waters, Milford, MA) as described (Buck et al. 2009). Samples were separated by BEH HILIC column (1.7 μm, 2.1 × 100 mm; Waters, Milford, MA). Multi-reaction monitoring (MRM) were used to quantify glutamate (148→ 84; 9.3 min), GABA (104→ 69; 4.5 min), dopamine (154→ 137; 4.3 min) and novaline (118→ 72; 9.8 min). Neurotransmitter profiles were calculated by the ratio of neurotransmitters to the average of the 1 h baseline measures of the individual mouse.
Operant conditioning
Operant conditioning experiments were performed using computer-controlled mouse operant chambers and as previously described (Reker et al. 2018). Throughout the training and testing period, mice were placed on a food restriction schedule to maintain 85% of their free-feeding weight and trained to press the correct lever in order to receive 20% sucrose rewards. Mice were first trained to associate a tone with the reward delivery during 30 min magazine-training sessions in which tones (1 s duration) were presented randomly using a hand clicker throughout the sessions and paired with delivery of 100 μl of the sucrose reward solution. For 5 consecutive days a week and twice a day, mice were administered 30 min training sessions under a fixed-ratio 1 (FR1) schedule for sucrose reinforcement. The mice were selected by successive approximation to elicit a lever press for sucrose. This operant behavior was maintained by FR1 until mice received 30 reinforcers within 30 min (Nam et al. 2013), at which point mice were considered to be trained. If mice did not meet this inclusion criteria, they were excluded from the study. We initially trained a total of 33 mice, of which 5 mice were excluded based on the criteria.
These 28 trained mice were arbitrarily selected for bilateral DREADD viral infusions in either the mPFC or NAc. 1 week after recovery from viral surgery, operant conditioning behaviors for 20% sucrose rewards were maintained by fixed ratio (FR1) until mice received 30 reinforcers. During the operant sessions, behaviors such as correct lever-pressing activity demonstrated reward-seeking, whereas head entry to obtain the reward denoted compulsive behavior. To obtain full viral expression, mice were under recovery for 3 weeks. Mice were employed 30 min operant sessions during which unlimited reinforcements were provided. Operant behaviors in response to saline control or CNO administration were recorded.
Operant conditioning for alcohol reinforcements entailed the same training methods and criteria for DREADD surgery as described above (n=7–9). However, after recovery from viral surgery, mice were exposed to alcohol reinforcements using a mixture of sucrose and alcohol. For the following 2 weeks, mice were trained for ethanol reinforcement by the sucrose fade-out method. In this method, the concentration of sucrose in the infusion pump connected to the operant chamber gradually decreases or fades-out in order to establish ethanol reinforcement. This sucrose fade-out method consists of a solution mixed with different concentrations of sucrose and ethanol every 2 days such that sucrose concentrations progressively decrease while ethanol concentrations progressively increase. For instance, on the first day of this method, the reward solution consisted of 10% alcohol in 20% sucrose solution. By the end of the second week, the solution only consisted of 10% alcohol and no sucrose and is thus when sucrose has faded-out and considered to be an ethanol reinforcement. After sucrose fade-out was complete and only ethanol was present, baseline measures of operant conditioning behaviors for 10% alcohol rewards were measured.
In the devaluation paradigm of operant conditioning, aversive conditioning was induced by injecting lithium chloride (LiCl; 20 mg/kg of 0.15 M, i.p) after mice went through operant sessions for alcohol reward. This was done for 6 consecutive days, once a day. Saline or CNO injections to activate Pv-FSI were given 30 min prior to operant conditioning sessions. To pair alcohol reward with an aversive event, LiCl injections were given after each operant conditioning session and before the mouse was returned to their home cage. In the extinction phase, mice were injected with either saline or CNO and put in the operant chamber 30 min later. During the extinction paradigm, mice did not receive any rewards for correct lever presses. Sound and light cues were also no longer present. Measures of operant behaviors for alcohol reward were recorded for both devaluation and extinction sessions.
All operant procedures were performed in computer-controlled mouse operant chambers (Model ENV-307W; Med Associates) equipped with house light. Operant behavior experiments were conducted between 9 am and 3 pm. All phases of operant sessions lasted 30 min. Measures of operant behaviors such as total lever presses and head entries were recorded for the sessions. No blinding was performed for these experiments.
Statistical analysis
All data were expressed as mean ± standard error of the mean (SEM). No sample calculation was performed, but sample size was determined by previous publications (Nam et al. 2011; Nam et al. 2013). For c-fos expression data (Fig.1), statistics were performed by two-tailed Student’s unpaired t-test since it was normally distributed (GraphPad Prism version 8.0.0, La Jolla, CA). For the mouse behavioral experiment (Fig.4), statistics were performed by Wilcoxon matched-pairs signed rank test. For microdialysis (Fig.2 and 3) and devaluation experiment (Fig.5), Kolmogorov-Smirnov (K-S) tests were conducted and then analyzed by two-way ANOVA followed by Tukey post hoc test (SigmaStat 13, SYSTAT software, Point Richmond, CA). No test for outliers was conducted. Criterion for statistical significance was p<0.05.
Fig. 1.
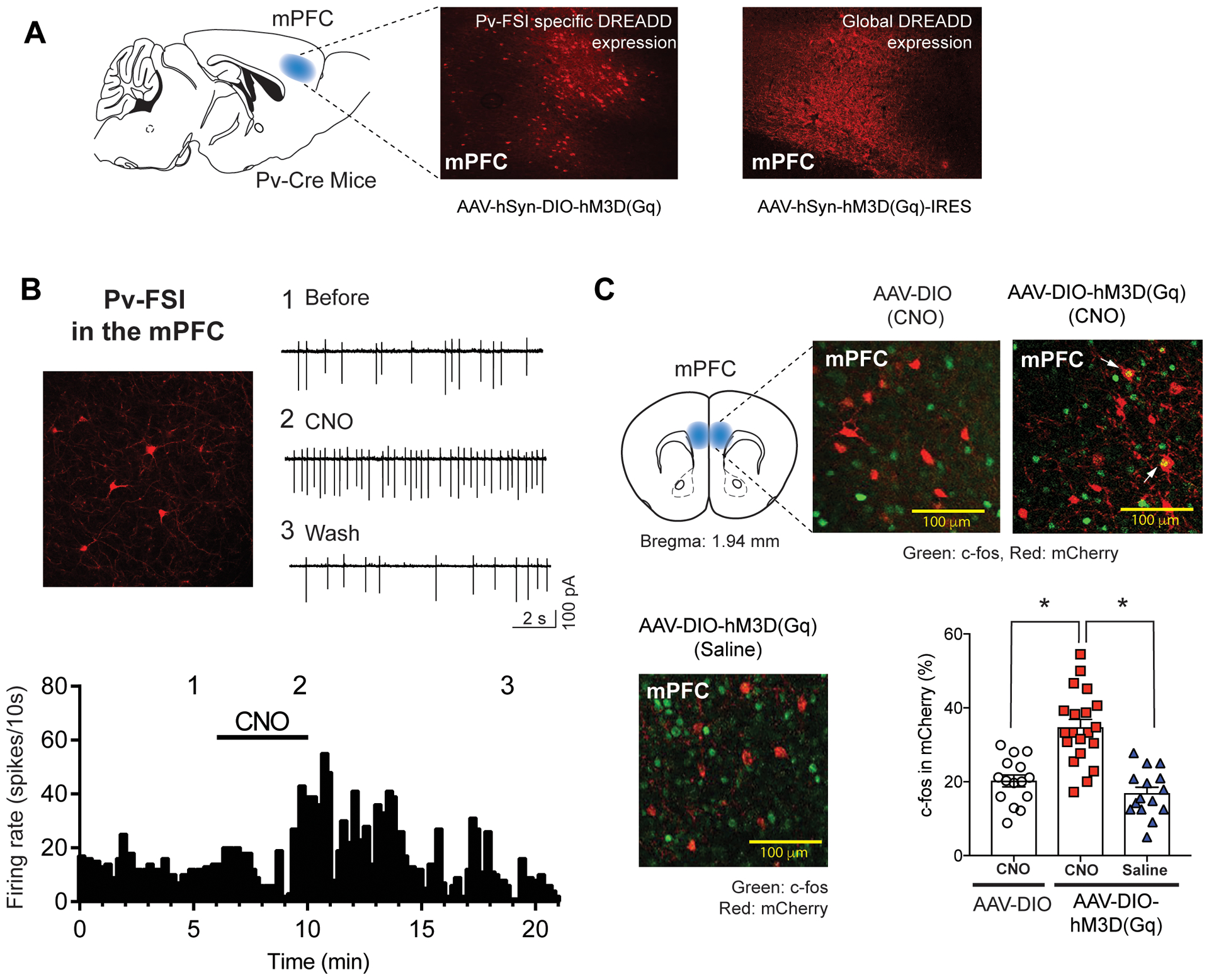
Parvalbumin-Fast Spiking Interneuron (Pv-FSI) activation in the mPFC using DREADD. The designer receptor, hM3Dq, mimics Gq and gets activated by the designer drug, Clozapine-N-Oxide (CNO), resulting in enhanced neuronal firing i.e. activation of Pv-FSI. (A) Representative image of DREADD expression in the mPFC of Pv-Cre mice. The Pv-FSI specific DREADD expression is induced by the Cre-dependent DREADD construct (AAV-hSyn-DIO-hM3D(Gq)-mCherry) in the Pv-Cre mice compared to that of global DREADD expression (AAV-hSyn-HA-hM3D(Gq)-IRES-mCitrine) in the Pv-Cre mice (n=3). (B) Electrophysiological representation of the firing rate of Pv-FSI before (session 1) and after (session 2) Pv-FSI DREADD activation following CNO bath application (session 3). Confocal image demonstrates Pv-FSI immunofluorescence (n=3) (C) Verification of DREADD activation in the mPFC of Pv-Cre mice using the stereological analysis of c-fos (green) immunoreactivity in the mCherry (red) expressing cells. CNO administration in the mice expressing DREADD (AAV-hSyn-DIO-hM3D(Gq)-mCherry) leads to a significantly high colocalized expression of double labeling yellow cells (white arrow) compared to CNO administration in Pv-Cre mice with control virus expression (AAV-hSyn-DIO-mCherry) or saline administration in Pv-Cre mice with DREADD expression (AAV-hSyn-DIO-hM3D(Gq)-mCherry). (*p< 0.05 unpaired t-test; n=9; 3 mice per group)
Fig 4.
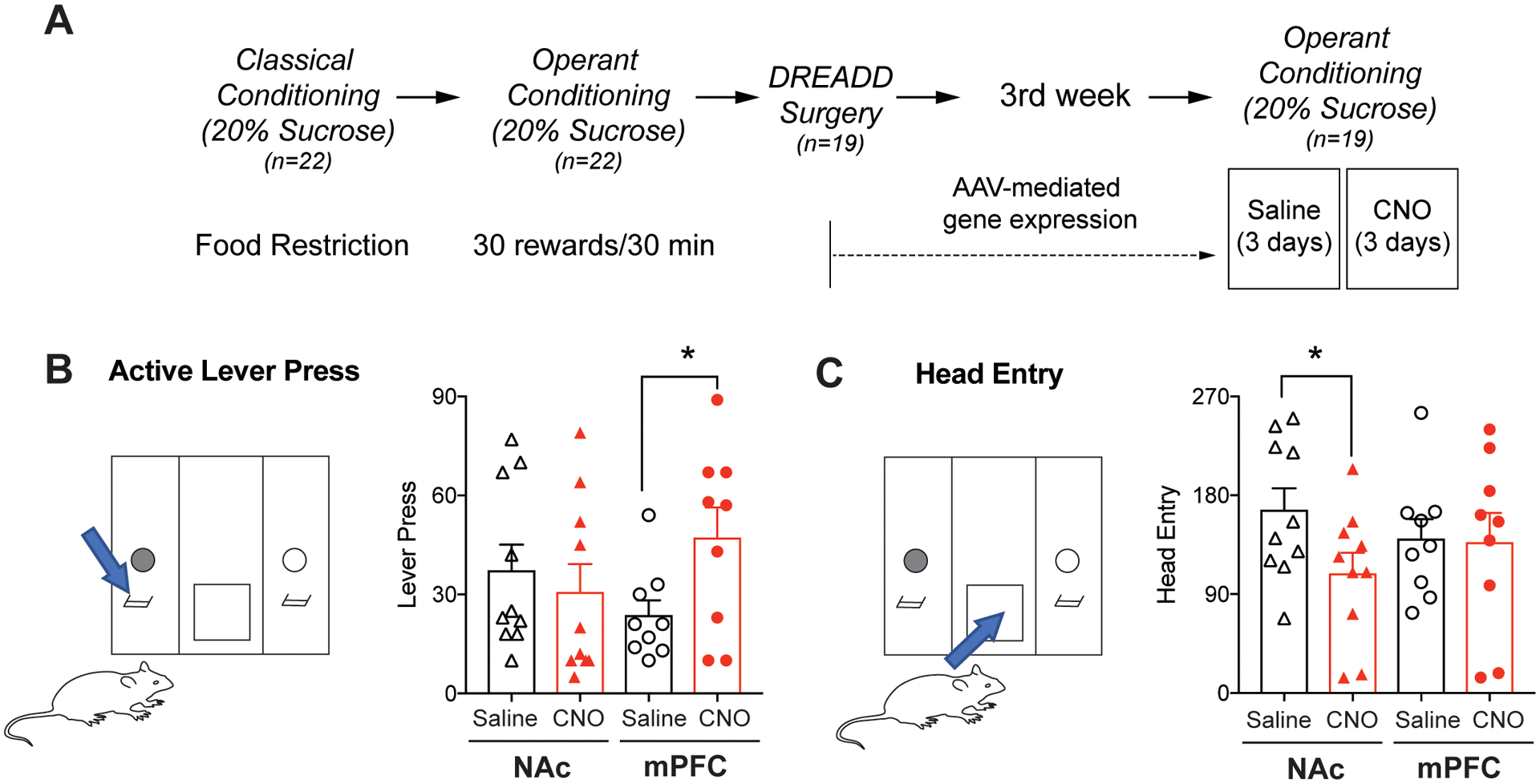
Operant behavior tasks measuring sucrose reward-seeking behavior. (A) Experimental design and timeline to test effects of Pv-FSI on reward-seeking behaviors for natural reward, sucrose. Mice were put on a food-restricted diet and simultaneously trained for operant conditioning to obtain sucrose rewards. 3 weeks after DREADD surgery, mice operant responding was measured. (B) Operant conditioning chamber in which mice must press the correct lever (blue arrow) to obtain rewards, denoting reward-seeking behavior. DREADD activation of Pv-FSI in the mPFC by CNO administration significantly increased goal-directed and reward-seeking behaviors shown by higher lever pressing activity compared to saline control (n=9–10 per group; *p< 0.05 paired t-test). (C) Laser beam in the food trough (blue arrow), where rewards are dispensed, senses and records mice head entry to obtain rewards, denoting compulsivity. DREADD activation of Pv-FSI in the NAc significantly decreased compulsive responding shown by lesser head entries compared to that of saline control. No difference in compulsive behavior seen in mice with DREADD activation of Pv-FSI in the mPFC (*p < 0.05 Wilcoxon matched-pairs signed rank test, n=10 for NAc DREADD expression, n=9 for mPFC DREADD expression). All data are presented as mean ± SEM.
Fig 2.
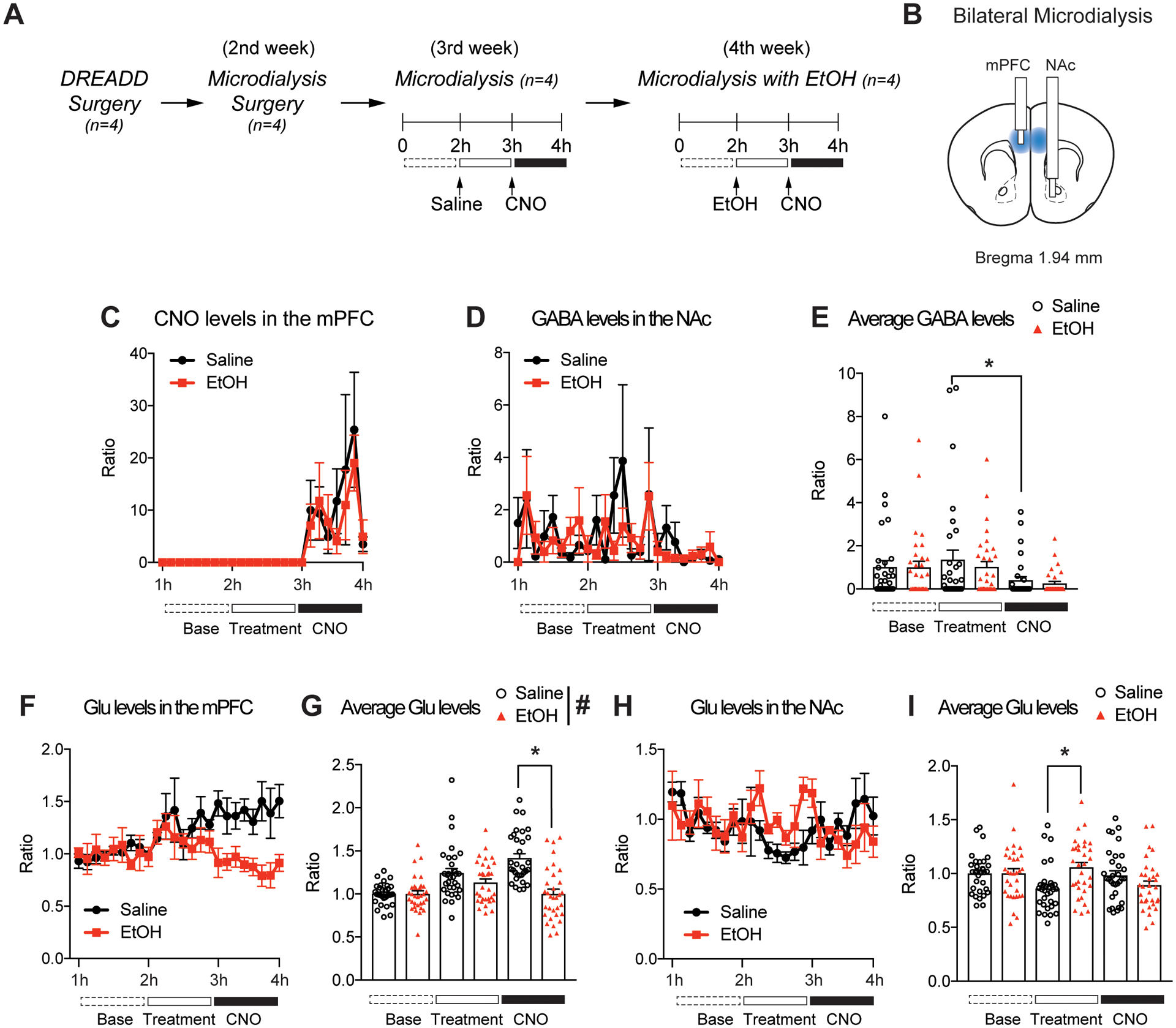
Bilateral microdialysis quantifying neurotransmitters in the mPFC and NAc. (A) Experimental design and timeline of microdialysis. Mice undergo DREADD viral surgery in the mPFC and then microdialysis surgery, followed by 4 h of microdialysis sessions testing the effect of alcohol (EtOH; 1.5g/kg, i.p) on Pv-FSI activity. (B) Blue regions indicate location of Gq-DREADD viral infusion. During bilateral microdialysis surgery, one of the microdialysis probes are surgically placed in the mPFC of the left hemisphere and the other is placed downstream in the NAc of the right hemisphere. (C) CNO is detected only after CNO (1 mg/kg, i.p) administration at 3 h. Pre-exposure to saline or alcohol do not cause differences in CNO levels between the two pre-treated groups. (D and E) Following Pv-FSI activation in the mPFC by CNO, GABA levels downstream in the NAc are diminished regardless of saline/EtOH pre-treatment (n=4 mice per treatment group; *p < 0.05 for main effect of session by two-way ANOVA; n=32 per average condition). (F and G) Pv-FSI activation in the mPFC by CNO attenuates glutamate (Glu) in the mPFC of EtOH pre-treated mice compared to saline control (n=4 mice per treatment group;*p < 0.05 by two-way ANOVA Tukey post hoc tests between saline and EtOH treatment; #p < 0.05 main effect of treatment by two-way ANOVA, n=32 per average condition). (H and I) EtOH pre-treatment significantly increase Glu in the NAc compared to saline control (n=4 mice per treatment group; *p< 0.05 by two-way ANOVA Tukey post hoc tests between saline and EtOH; n=32 per average condition). All data are presented as mean ± SEM.
Fig 3.
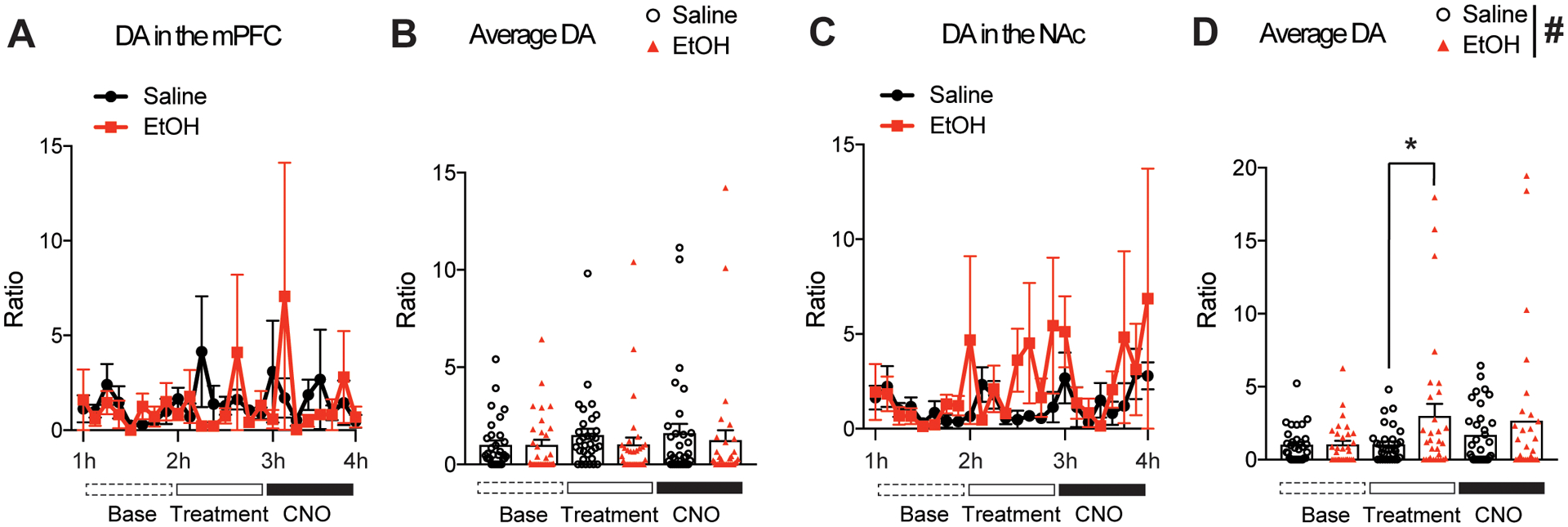
Dopamine (DA) levels in the mPFC and NAc during Pv-FSI activation after EtOH exposure (EtOH; 1.5g/kg, i.p). (A and B) DA levels in the mPFC do not change under EtOH pre-treatment nor saline, and CNO activation of Pv-FSI does not induce changes in DA levels for either pre-exposed groups. (C and D) EtOH induces a marked increase of DA in the NAc compared to that of saline control. CNO administration did not significantly alter DA levels after saline or ethanol pre-treatment (n=4 mice per treatment group; *p < 0.05 compared to that of saline pre-treated group; #p < 0.05 main effect of treatment by two-way ANOVA; n=32 per average condition). All data are presented as mean ± SEM.
Fig 5.
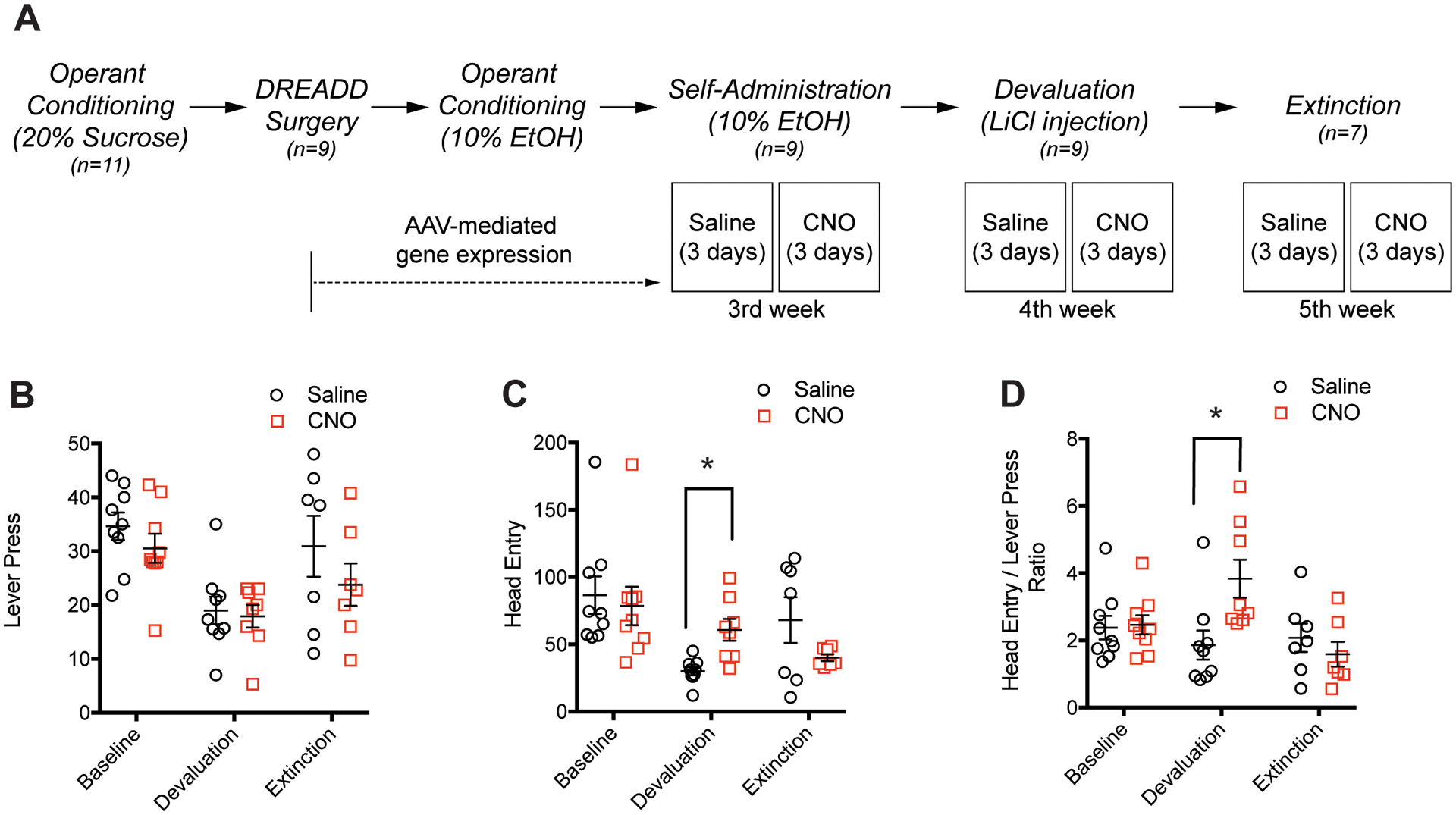
Operant behavior tasks measuring alcohol reward-seeking behavior. (A) Experimental design and timeline of operant conditioning for alcohol self-administration, devaluation, and extinction paradigms. (B) Lever pressing activity of mice when Pv-FSI is activated by CNO decreases during devaluation and increases back to near-baseline levels during extinction. (C) Pv-FSI activation by CNO results in more compulsive responding, shown by the increased head entries compared to the significant decrease in saline control from baseline levels (*p < 0.05 for main effect of operant session by two-way ANOVA). (D) Pv-FSI activation by CNO increases the ratio of head entries to lever presses in devaluation compared to saline (*p < 0.05 for main effect of operant phase by two-way ANOVA; 9 mice were used for baseline and devaluation experiments, 7 mice were used for extinction experiments). All data are presented as mean ± SEM.
Results
Pv-FSI-specific DREADD expression and functional validation of the DREADD model
To selectively activate Pv-FSI in the corticostriatal circuitry, we bilaterally injected DREADD viruses into the mPFC of Pv-Cre mice. We induced either Pv-FSI-specific DREADD expression (AAV-hSyn-DIO-hM3D(Gq)-mCherry) or global DREADD expression (AAV-hSyn-HA-hM3D(Gq)-IRES-mCitrine) in as a control (Fig. 1A). Immunohistochemistry results show robust red fluorescence of Pv-FSI surrounding the area of DREADD viral implantation in the mPFC. Results show differences in neuronal expression of Pv-FSI-specific DREADD and Global DREADD, which activates Pv-FSI and other neuronal cells in that brain region. The image of the Pv-FSI-specific DREADD compared to global expression demonstrates results confirming that Pv-FSI are a smaller neuronal population, indicated by the fewer area of fluorescence shown in the panel.
Moreover, to test Gq in modulating Pv-FSI activity in vitro, patch-clamp membrane potential recordings were performed on the mCherry-expressing neurons in mice with the Pv-FSI-specific DREADD virus. CNO increased the firing rate of Gq-expressing Pv-FSI compared to before CNO administration (Fig. 1B). Following washout, the neuronal firing decreased as expected due to less CNO to induce activation of the designer Gq receptors. These electrophysiological attributes resembled those reported previously showing an increase in Pv-FSI firing rate using optogenetic activation and multiunit electrode recording (Sparta et al. 2014). Our data also demonstrate that the DREADD technology can reliably activate Pv-FSI.
Because c-fos expression in the brain is an indication of neural activation, we wanted to examine whether CNO activates target Pv-FSI in the mPFC in vivo after DREADD viral implantation. We measured the overlap of c-fos expression in mCherry-expressing Pv-FSI to assess the effect of CNO on activating Pv-FSI. Overlaid images depict significantly more c-fos expression under CNO administration in Pv-FSI-specific DREADD (AAV-hSyn-DIO-hM3D(Gq)-mCherry) compared to when CNO was administered under the control virus (AAV-hSyn-DIO-mCherry) (t(33)= 5.025, p< 0.0001). In addition, CNO administration also induced more c-fos expression than when saline was administered in Pv-FSI-specific DREADD (t(33)= 6.151, p< 0.0001) (Fig. 1C).
Measurement of neurochemical changes in both the mPFC and NAc simultaneously using in vivo microdialysis
As Pv-FSI in the mPFC are vast inhibitory connections and have rapid firing rates, we wanted to determine the impact of DREADD-induced activation of Pv-FSI. To determine if Pv-FSI activation in the mPFC alters neurotransmission and whether interaction with alcohol also affects Pv-FSI functioning, we quantified neurochemical changes in the mPFC and NAc using in vivo bilateral microdialysis. Because Pv-FSI activation in the mPFC can subsequently activate the striatum downstream and affect the corticostriatal circuitry, we wanted to examine possible changes in the neurotransmitter levels that are housed in the circuitry. To do this, we performed bilateral microdialysis on mice 1 week after the Pv-FSI-specific DREADD virus was surgically implanted, and then collected dialysates from the mPFC and NAc (Fig. 2A). Since the Gq-DREADD viral infusion was done bilaterally in the mPFC, we wanted to ascertain whether bilateral microdialysis could be a reliable representation of the communication in the corticostriatal circuitry. We were able to measure neurotransmitter levels in this circuitry by positioning one microdialysis probe in the mPFC and the other downstream in the NAc (Fig. 2B). In order to test for reliability, we first needed to confirm whether CNO can be detected in the mPFC following CNO i.p administration. Results show that CNO lasted in the mPFC for 1 h after injection (Fig. 2C). In addition, saline or alcohol injections given to mice prior to CNO administration did not impact CNO levels in the mPFC.
As both Pv-FSI and alcohol can regulate the corticostriatal circuitry, we wanted to examine whether and how they might induce changes in local extracellular neurotransmitter release. The neurotransmitter levels were compared between groups that received saline injections and alcohol injections prior to the administration of CNO to activate Pv-FSI. This allowed us to see if alcohol pre-treatment caused differences in Pv-FSI functioning and elucidate the impact on the changes in neurotransmission. It was difficult to detect GABA in the dialysates from the mPFC region however, GABA was detectable in the NAc (Fig. 2D). Two-way ANOVA indicated a significant main effect of CNO during the microdialysis procedure (F(2,186) = 4.961, p < 0.01). Tukey post hoc test identified that average GABA levels show a significant difference between saline pre-treatment and CNO trials (Fig. 2E). As expected, GABA levels show a significant decrease after CNO administration for saline pre-treated mice. Pre-treatment of alcohol did not affect GABA levels significantly after Pv-FSI activation.
Alcohol inhibits Pv-FSI activation and decreases glutamate levels in the mPFC
As most of the cortical outputs are glutamatergic, it was important to look at how Pv-FSI activation in the corticostriatal circuitry affects glutamate neurotransmission and how it is affected in alcohol exposure. We found that not only does Pv-FSI in the mPFC affect glutamate neurotransmission, but that it can do so differentially under alcohol pre-treatment. Two-way ANOVA showed a significant main effect in trials (F(2,186) = 12.671, p < 0.001), treatment (F(2,186) = 22.35, p < 0.001), and an interaction between session and treatment (F2,186) = 11.32, p < 0.001) (Fig. 2F). Tukey post hoc analysis revealed that a significant difference in glutamate levels occurred during CNO sessions (Fig. 2G). Mice in alcohol pre-treatment trials showed a significant decline in glutamate in the mPFC following CNO activation of Pv-FSI, compared to the steady increase of glutamate levels observed in the saline pre-exposed control trials.
Glutamate levels in the NAc, however, did not show such pronounced differences as seen in the mPFC. Two-way ANOVA showed an interaction between trial and treatment of either saline control or CNO (F(2,186) = 4.114, p = 0.018) (Fig. 2H). Tukey post hoc tests identified that mice had a significant increase in glutamate during the alcohol pre-treatment trials compared to that of saline control (p = 0.014) (Fig. 2I) however, there were no changes in glutamate levels following Pv-FSI activation regardless of pre-treatment. Together, these results suggest that Pv-FSI activation in the mPFC increased glutamate release in the mPFC while there were no changes in these levels in the NAc. Overall, results indicate that Pv-FSI activation can induce changes in neurocircuitry and neurotransmission, specifically with a greater and more profound effect on GABA in the NAc and glutamate in the mPFC.
Alcohol treatment in mice induces changes in dopamine neurotransmission in the NAc
To examine whether dopamine neurotransmission is significant in the corticostriatal circuitry and the reward system relating to Pv-FSI activation, we also looked at dopamine levels in target brain regions. Measurements showed that alcohol pre-treatment prior to CNO administration did not cause significant changes in dopamine neurotransmitters in the mPFC compared to that of their saline controls, and that alcohol did not alter dopamine levels following Pv-FSI activation by CNO administration (Fig. 3A and 3B). In contrast, Two-way ANOVA showed a significant main effect in treatment for the dialysates of the NAc (F(2,186) =5.119, p = 0.025). Tukey post hoc tests identified that mice had a significant increase in dopamine during the alcohol pre-treatment trials compared to that of saline control (p=0.024) (Fig. 3C and 3D). This finding is consistent with literature that show alcohol increases dopamine release in the NAc (Boileau et al. 2003).
Pv-FSI activation in the mPFC increases sucrose reward-seeking behaviors
After confirming the functional effect of Gq DREADD activation of Pv-FSI and observing drastic molecular changes, we examined the behavioral effects of Pv-FSI activation in regards to reward seeking. We trained Pv-Cre mice to seek sucrose rewards while keeping them under a food-restricted diet. 3 weeks after DREADD viral surgery, operant responding for sucrose rewards under a fixed ratio 3 (FR3) schedule was recorded (Fig. 4A). CNO administration was given to activate Pv-FSI and saline administration served as a control to observe the normal basal function of Pv-FSI. During the operant conditioning sessions, mice learned to press the correct lever in order to obtain sucrose reward through the food trough. CNO treatment significantly increased the number of lever presses for mice with the DREADD virus and Pv-FSI activation in the mPFC (p=0.023) compared to that of saline control. There were no changes in lever-pressing activity for mice with Pv-FSI-specific DREADD activation in the NAc (Fig. 4B).
Moreover, we recorded changes in compulsive behavior for sucrose rewards. To measure this, we observed the number of mice head entry to obtain sucrose reward by using the infrared beam sensor in the food trough to sense movement (Fig. 4C). Activating only Pv-specific neurons in the NAc region decreased compulsive responding significantly (p=0.029), as demonstrated by the lower number of head entries when given CNO compared to that of saline. Taken together, these behavioral patterns of lever pressing activity and head entry demonstrate that CNO activation of Pv-FSI have regional effects on motivational behavior such that activation in the mPFC increases reward-seeking behavior and activation in the NAc decreases compulsive behavior.
Pv-FSI activation promotes compulsive head entry during alcohol devaluation
Then, we examined whether Pv-FSI regulates reward seeking for alcohol rewards. Since alcohol exposure is known to inhibit GABA release from interneurons (Homayoun & Moghaddam 2007), we examined the impact of Pv-FSI activation on the mPFC and how it may regulate alcohol-seeking behaviors. We used operant conditioning and a sucrose fading-out method to provide 10% alcohol rewards (Fig. 5A). To further investigate the role of Pv-FSI activation during alcohol reward seeking, we examined alcohol reinforcement in the devaluation period, by inducing aversion with LiCl injections, and in the extinction period (Dickinson et al. 2002; Nam et al. 2013).
We found that Pv-FSI activation in the mPFC did not induce significant changes in alcohol reward-seeking behaviors by lever-pressing activity nor head entry. Therefore, we employed an alcohol reward devaluation and extinction paradigm using operant conditioning. Both the CNO and saline treatment groups demonstrated decreased lever-pressing activity during the devaluation stage and a recovery pattern during the extinction period where lever presses go back to near baseline levels, although the lever-pressing activity did not differ between treatment groups. Two-way ANOVA tests indicate a significant main effect of lever pressing for alcohol seeking during devaluation and extinction (F(2, 48) = 10.33, p < 0.001), while there is no effect of CNO treatment in either behavioral sessions (Fig. 5B).
Head entries for alcohol seeking are significantly changed during devaluation and extinction sessions, but there is no overall effect of Pv-FSI activation by CNO treatment. Two-way ANOVA identified that head entries significantly differed for devaluation and extinction phases during Pv-FSI activation by CNO (F(2, 48) = 6.187, p = 0.004), with an interaction between phases and treatment groups (F(2, 48) = 3.29, p = 0.004). Multiple comparisons using Tukey tests showed that upon CNO administration and consequent Pv-FSI activation, mice show a significant increase in head entries during the devaluation session (Fig. 5C). These results implicate that Pv-FSI activation in the mPFC increases compulsivity during the devaluation period of alcohol seeking.
To determine the degree of compulsivity due to Pv-FSI activation, we measured the ratio between head entries and lever-pressing activity for alcohol reward. Two-way ANOVA tests indicate a significant main effect of alcohol seeking during devaluation and extinction (F(2, 48) = 3.04, p = 0.032) and an interaction between behavioral sessions and CNO treatment (F(2, 48) = 4.85, p = 0.013). Tukey tests identified that Pv-FSI activation by CNO administration significantly increases the ratio for head entries per lever press during the devaluation session. Saline-treated mice demonstrate a 1.86 ratio (head entry/lever press) while CNO-treated mice demonstrate a 3.84 ratio (head entries/lever press) during the devaluation (Fig. 5D). Overall, our findings suggest that Pv-FSI activation in the mPFC increases natural reward seeking such as for sucrose, but this mechanism is not applicable for alcohol reinforcement. However, Pv-FSI activation in the mPFC promotes compulsivity during the alcohol devaluation period.
Discussion
Many researchers have been keen on evaluating the role of the corticostriatal circuit in reward-seeking behaviors. Although it is known that the GABA-glutamate balance plays an important role in this process, exactly how the circuit is interconnected and what cellular subtypes contribute to aspects of addiction are unknown. Our current study demonstrates how the Pv-FSI microcircuit regulates corticostriatal neurotransmission and behavioral outcomes related to self-administration, devaluation, and extinction using operant conditioning. Utilizing electrophysiology and microdialysis, we demonstrated that Pv-FSI activation regulates glutamate levels in the mPFC and that alcohol may inhibit Pv-FSI functioning. Interestingly, as Pv-FSI activation in the mPFC promotes natural reward seeking for sucrose by increasing lever-pressing activity but does not induce reward seeking for alcohol, it seems that alcohol itself suppresses Pv-FSI function. Overall, although alcohol self-administration and head entry showed no significant behavioral changes in response to CNO treatment compared to those of saline, deflection between the devaluation trials demonstrates critical evidence that Pv-FSI may influence reward-seeking properties.
Although our data shows that local Pv-FSI microcircuitry is necessary for the increase in reward-seeking activity and decrease in compulsivity for sucrose reward, it is not enough to explain the effect of GABAergic interneurons on excitatory glutamatergic neurons or neighboring neurons in the mPFC. In addition, Pv-FSI may inherently be hyperactive in propagating inhibitory signals, as they are fast-spiking GABAergic cells and cause increased oscillations of gamma waves. Thus, it would be ideal to have studies incorporating Pv-FSI inhibition of neuronal firing using a Cre-dependent Gi-DREADD system (Owen et al. 2018). This approach would allow us to confirm the results of the hM3Dq receptors and examine whether Pv-FSI activation or Pv-FSI inhibition is required for reward seeking and compulsivity. Moreover, measuring neurotransmitters using in vivo microdialysis would enable us to better understand the connection between the molecular effects of Pv-FSI activation to modulate glutamate and GABA levels in the corticostriatal circuit and the behavioral effects. Theoretically, Pv-FSI activation in the mPFC would mean the inhibitory signal from the GABAergic Pv-FSI would decrease the firing of the excitatory signal from the pyramidal neurons. However, with Pv-FSI activation in the mPFC, less excitatory signals would get passed on downstream to the MSNs to stimulate them and elicit their effects. Thus, less excitatory activity in the mPFC could mean more GABAergic activation in the NAc. Our results from two-way ANOVA indicated that for glutamate, Pv-FSI activation by CNO had more of an effect in the mPFC than in the NAc. We also found that alcohol significantly changed glutamate levels in the NAc. Furthermore, alcohol pre-exposure affected Pv-FSI functioning for GABA/glutamate/dopamine neurotransmission and interestingly, differentially between brain regions.
Because the striatum receives glutamatergic inputs from the cortex and consists mostly of MSNs, which primarily have dopamine D1-and D2-type neurons (Groenewegen et al. 1996; Britt et al. 2012), it would be important to look at the effects of Pv-FSI activation on the changes in dopamine levels. Our results demonstrated that Pv-FSI activation in the mPFC increases reward seeking (Fig.4B), while Pv-FSI activation in the NAc decreases head entry without changes in reward seeking (Fig.4C). Since dopamine plays a significant role in the reward system, Pv-FSI activation in the mPFC and NAc can be important for the effects seen on reward-seeking behavior and compulsivity. Therefore, we measured operant behaviors during two additional reward seeking paradigms, devaluation and extinction. Assessing behavior in these phases were critical to demonstrate the severity of reward-seeking and compulsivity, since the devaluation session should ideally decrease reward seeking and compulsivity due to the aversive effect of LiCl after each operant session. Interestingly, Pv-FSI activation in the mPFC increases compulsive responding during the alcohol devaluation period. The extinction session should ideally further decrease reward seeking and compulsivity because of the lack of rewards and cues during this operant session (Samson et al. 2004). However, our data demonstrated that Pv-FSI activation in mice mPFC impairs their ability to undergo the normal extinction process.
Our bilateral microdialysis results can contribute to the understanding of the neural population of the corticostriatal circuitry and their neurotransmitter functioning due to the presence of alcohol. Pv-FSI are GABAergic cells and activating them would mean less GABA release downstream in the NAc. As expected, activation of Pv-FSI in the mPFC by systemic administration of CNO significantly decreased the levels of GABA in the NAc for saline-treated mice. Interestingly, the same decrease in GABA following Pv-FSI activation by CNO was seen in alcohol-treated mice as well; this indicates that alcohol may not affect GABA neurotransmission mediated by Pv-FSI activation any differently than it does for saline control. However, alcohol pre-exposure inhibits glutamate increase induced by Pv-FSI activation in the mPFC. Chronic alcohol treatment is known to shrink the dendritic tree of parvalbumin interneurons in the striatum as compared to saline controls (De Giorgio et al. 2012). In line with this physiological effect of alcohol on Pv-FSI, we also observed molecular effects of alcohol on Pv-FSI. For example, alcohol pre-exposure itself increases glutamate in the NAc but has no effect on Pv-FSI functioning pertaining to glutamatergic neurotransmission in the NAc (Fig.2I). This is possibly due to the fact that majority of the neurons in the mPFC are glutamatergic pyramidal neurons and less are in the NAc, thus, it senses and contributes to a greater change in the glutamatergic levels in the NAc. In addition, dopamine levels were not significant in the mPFC, and this could be because Pv-FSI in the cortex may not regulate dopamine release.
The combination of DREADD and simultaneous microdialysis is a novel in vivo approach to demonstrate the real-time neurochemical changes in the cortico-striatal circuitry. Even though there were several technical difficulties, we have adjusted the experimental design to overcome them successfully. For example, DREADD viral expression needs to be limited to the mPFC and NAc, but its expression is only enough to cover the microdialysis probing regions (around 1 mm3). Therefore, the injection volume for virus particle and the coordinates for stereotaxic surgery need to be adjusted and then confirmed by histological sectioning. In addition, as neurochemical changes in mice are affected by stress or handling, a vehicle control must be included to compare the effect of CNO administration. To confirm DREADD activation during the experimental period, we also simultaneously detected CNO from the brain dialysate using LC-MS/MS (Fig.2C). These technical advancements enable us to determine neurochemical changes in response to DREADD activation of Pv-FSI and provide the evidence to design a behavioral timeline such as the start of recording measurements 30 min after DREADD activation by CNO.
In the brain, alcohol is known to promote dopamine release in the NAc (Boileau et al. 2003). The transient increase of dopamine in the NAc after alcohol administration (Fig.3) could be due to the dopaminergic output from other brain regions that are also a part of the reward circuitry but not a part of the corticostriatal circuitry specifically. Mesolimbic pathways and other pathways involving dopamine and the reward system can play a role as well (Hodge et al. 1997). For example, previous studies suggest that the mesopallidal, mesoaccumbens, and nigrostriatal dopamine systems are more sensitive to the effects of alcohol than the nigropallidal dopamine system (Melendez et al. 2003). Therefore, much research advocates that the mesolimbic pathway is involved in motivation, cognition, and the reward system. However, there has not been as many advancements in this. In fact, research has shown that dopamine is not the sole contributor. For instance, knockout of DAT transporters in mice still show an increase in cocaine self-administration, indicating that there are other neurotransmitter systems at play in contributing to reward-seeking (Koob & Volkow 2010).
Moreover, sexual dimorphism was observed in the developmental effects of Pv-FSI in the mPFC (Goodwill et al. 2018) and NAc (Ravenelle et al. 2014). As disturbances in reward-seeking behaviors such as substance abuse, addiction, and even depression also present gender as a demographic difference, it is imperative to research such differences in Pv-FSI activity related to reward-seeking behaviors. This can provide further insight into how Pv-FSI can contribute to the cognitive and behavioral dysfunction and the neurochemical imbalance that occurs in reward-seeking impairments and therefore also provide individualized treatment methods based on gender. Therefore, further study is required to clarify Pv-FSI functioning not only in specific brain regions but also in its function in an age- and sex-dependent manner.
There are some caveats associated with the biotransformation of CNO to clozapine (Manvich et al. 2018), which may affect behavioral changes in mice. Clozapine is a dopaminergic drug and chronic clozapine administration is known to increase dopamine levels in the NAc shell (Shilliam & Dawson 2005), which may affect reward seeking. However, we observed that CNO treatment did not alter dopamine levels in the NAc compared to saline control. We considered there to be an interaction between alcohol and clozapine since one clinical study reported that schizophrenia patients taking clozapine treatment experienced significant reductions in severity of alcohol abuse (Drake et al. 2000). The amount of CNO and duration of treatment in our study may not transform CNO to clozapine in enough levels to induce the behavioral changes in mice. Furthermore, we validated our Cre-dependent Gq-DREADD system and compared results to the AAV-DIO-mCherry control; we assessed the neural activity of DREADD activation by measuring c-fos intensity in the Pv-FSI in response to CNO. However, future endeavors using Gi-DREADD with CNO will enable us to exclude the role of CNO during reward seeking.
Overall, the results from our findings suggest that not only does Pv-FSI contribute to changes in the corticostriatal circuitry and reward-seeking behavior, but also that alcohol has an effect on Pv-FSI functioning. This is shown through a series of operant behavior tasks and quantification of neurotransmitters from microdialysis procedures.
Acknowledgments
We would like to thank Dr. John Sullivan for his technical support. We would like to appreciate Dr. Nicholas Goeders and Dr. Nancy Leidenheimer for their comments and critiques of this work.
Funding and Disclosure
The authors declare no conflict of interest. This work was supported by COBRE (1P20GM121307-01A1) from NIGMS, NARSAD Young Investigator Award (26530) from the Brain & Behavior Research Foundation, BRF Seed Funding Program from Biomedical Research Foundation to HWN.
Abbreviations used:
- CNO
clozapine-N-oxide
- DA
dopamine
- DREADD
designer receptors exclusively activated by designer drugs
- FR1
fixed-ratio 1
- GABA
γ-aminobuturic acid
- LiCl
lithium-chloride
- mPFC
medial prefrontal cortex
- MSNs
medium spiny neurons
- NAc
nucleus accumbens
- Pv-FSI
Parvalbumin expressing fast-spiking interneurons
References
- Barker JM, Corbit LH, Robinson DL, Gremel CM, Gonzales RA and Chandler LJ (2015) Corticostriatal circuitry and habitual ethanol seeking. Alcohol 49, 817–824. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boileau I, Assaad JM, Pihl RO, Benkelfat C, Leyton M, Diksic M, Tremblay RE and Dagher A (2003) Alcohol promotes dopamine release in the human nucleus accumbens. Synapse 49, 226–231. [DOI] [PubMed] [Google Scholar]
- Britt JP, Benaliouad F, McDevitt RA, Stuber GD, Wise RA and Bonci A (2012) Synaptic and behavioral profile of multiple glutamatergic inputs to the nucleus accumbens. Neuron 76, 790–803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buck K, Voehringer P and Ferger B (2009) Rapid analysis of GABA and glutamate in microdialysis samples using high performance liquid chromatography and tandem mass spectrometry. J Neurosci Methods 182, 78–84. [DOI] [PubMed] [Google Scholar]
- Burguiere E, Monteiro P, Mallet L, Feng G and Graybiel AM (2015) Striatal circuits, habits, and implications for obsessive-compulsive disorder. Curr Opin Neurobiol 30, 59–65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Giorgio A, Comparini SE, Intra FS and Granato A (2012) Long-term alterations of striatal parvalbumin interneurons in a rat model of early exposure to alcohol. J Neurodev Disord 4, 18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dickinson A, Wood N and Smith JW (2002) Alcohol seeking by rats: action or habit? Q J Exp Psychol B 55, 331–348. [DOI] [PubMed] [Google Scholar]
- Drake RE, Xie H, McHugo GJ and Green AI (2000) The effects of clozapine on alcohol and drug use disorders among patients with schizophrenia. Schizophr Bull 26, 441–449. [DOI] [PubMed] [Google Scholar]
- Gonzalez-Burgos G and Lewis DA (2012) NMDA receptor hypofunction, parvalbumin-positive neurons, and cortical gamma oscillations in schizophrenia. Schizophr Bull 38, 950–957. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goodwill HL, Manzano-Nieves G, LaChance P et al. (2018) Early Life Stress Drives Sex-Selective Impairment in Reversal Learning by Affecting Parvalbumin Interneurons in Orbitofrontal Cortex of Mice. Cell Rep 25, 2299–2307 e2294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Groenewegen HJ, Wright CI and Beijer AV (1996) The nucleus accumbens: gateway for limbic structures to reach the motor system? Prog Brain Res 107, 485–511. [DOI] [PubMed] [Google Scholar]
- Hodge CW, Samson HH and Chappelle AM (1997) Alcohol self-administration: Further examination of the role of dopamine receptors in the nucleus accumbens. Alcoholism: Clinical and Experimental Research 21, 1083–1091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Homayoun H and Moghaddam B (2007) NMDA receptor hypofunction produces opposite effects on prefrontal cortex interneurons and pyramidal neurons. J Neurosci 27, 11496–11500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu H, Gan J and Jonas P (2014) Interneurons. Fast-spiking, parvalbumin(+) GABAergic interneurons: from cellular design to microcircuit function. Science 345, 1255263. [DOI] [PubMed] [Google Scholar]
- Jinno S and Kosaka T (2004) Parvalbumin is expressed in glutamatergic and GABAergic corticostriatal pathway in mice. J Comp Neurol 477, 188–201. [DOI] [PubMed] [Google Scholar]
- Kang S, Li J, Zuo W, Fu R, Gregor D, Krnjevic K, Bekker A and Ye JH (2017) Ethanol Withdrawal Drives Anxiety-Related Behaviors by Reducing M-type Potassium Channel Activity in the Lateral Habenula. Neuropsychopharmacology 42, 1813–1824. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim H, Ahrlund-Richter S, Wang X, Deisseroth K and Carlen M (2016) Prefrontal Parvalbumin Neurons in Control of Attention. Cell 164, 208–218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim T, Thankachan S, McKenna JT et al. (2015) Cortically projecting basal forebrain parvalbumin neurons regulate cortical gamma band oscillations. Proc Natl Acad Sci U S A 112, 3535–3540. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koob GF and Volkow ND (2010) Neurocircuitry of addiction. Neuropsychopharmacology 35, 217–238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee K, Holley SM, Shobe JL, Chong NC, Cepeda C, Levine MS and Masmanidis SC (2017) Parvalbumin Interneurons Modulate Striatal Output and Enhance Performance during Associative Learning. Neuron 93, 1451–1463 e1454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Madisen L, Zwingman TA, Sunkin SM et al. (2010) A robust and high-throughput Cre reporting and characterization system for the whole mouse brain. Nat Neurosci 13, 133–140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Manvich DF, Webster KA, Foster SL, Farrell MS, Ritchie JC, Porter JH and Weinshenker D (2018) The DREADD agonist clozapine N-oxide (CNO) is reverse-metabolized to clozapine and produces clozapine-like interoceptive stimulus effects in rats and mice. Sci Rep 8, 3840. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Melendez RI, Rodd-Henricks ZA, McBride WJ and Murphy JM (2003) Alcohol stimulates the release of dopamine in the ventral pallidum but not in the globus pallidus: a dual-probe microdialysis study. Neuropsychopharmacology 28, 939–946. [DOI] [PubMed] [Google Scholar]
- Nam HW, Hinton DJ, Kang NY, Kim T, Lee MR, Oliveros A, Adams C, Ruby CL and Choi DS (2013) Adenosine transporter ENT1 regulates the acquisition of goal-directed behavior and ethanol drinking through A2A receptor in the dorsomedial striatum. The Journal of neuroscience : the official journal of the Society for Neuroscience 33, 4329–4338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nam HW, Lee MR, Zhu Y et al. (2011) Type 1 equilibrative nucleoside transporter regulates ethanol drinking through accumbal N-methyl-D-aspartate receptor signaling. Biol Psychiatry 69, 1043–1051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Owen SF, Berke JD and Kreitzer AC (2018) Fast-Spiking Interneurons Supply Feedforward Control of Bursting, Calcium, and Plasticity for Efficient Learning. Cell 172, 683–695 e615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Page CE, Shepard R, Heslin K and Coutellier L (2019) Prefrontal parvalbumin cells are sensitive to stress and mediate anxiety-related behaviors in female mice. Sci Rep 9, 19772. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ravenelle R, Neugebauer NM, Niedzielak T and Donaldson ST (2014) Sex differences in diazepam effects and parvalbumin-positive GABA neurons in trait anxiety Long Evans rats. Behav Brain Res 270, 68–74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reker AN, Oliveros A, Sullivan JM 3rd et al. (2018) Neurogranin in the nucleus accumbens regulates NMDA receptor tolerance and motivation for ethanol seeking. Neuropharmacology 131, 58–67. [DOI] [PubMed] [Google Scholar]
- Roth BL (2016) DREADDs for Neuroscientists. Neuron 89, 683–694. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Samson HH, Cunningham CL, Czachowski CL, Chappell A, Legg B and Shannon E (2004) Devaluation of ethanol reinforcement. Alcohol 32, 203–212. [DOI] [PubMed] [Google Scholar]
- Shilliam CS and Dawson LA (2005) The effect of clozapine on extracellular dopamine levels in the shell subregion of the rat nucleus accumbens is reversed following chronic administration: comparison with a selective 5-HT(2C) receptor antagonist. Neuropsychopharmacology 30, 372–380. [DOI] [PubMed] [Google Scholar]
- Sparta DR, Hovelso N, Mason AO, Kantak PA, Ung RL, Decot HK and Stuber GD (2014) Activation of prefrontal cortical parvalbumin interneurons facilitates extinction of reward-seeking behavior. J Neurosci 34, 3699–3705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sullivan JM 3rd, Grant CA, Reker AN, Nahar L, Goeders NE and Nam HW (2019) Neurogranin regulates sensorimotor gating through cortico-striatal circuitry. Neuropharmacology 150, 91–99. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Todtenkopf MS, Stellar JR, Williams EA and Zahm DS (2004) Differential distribution of parvalbumin immunoreactive neurons in the striatum of cocaine sensitized rats. Neuroscience 127, 35–42. [DOI] [PubMed] [Google Scholar]
- Wang X, Gallegos DA, Pogorelov VM, O’Hare JK, Calakos N, Wetsel WC and West AE (2018) Parvalbumin Interneurons of the Mouse Nucleus Accumbens are Required For Amphetamine-Induced Locomotor Sensitization and Conditioned Place Preference. Neuropsychopharmacology 43, 953–963. [DOI] [PMC free article] [PubMed] [Google Scholar]