

Abstract
Background
Microcirculatory disturbance is closely associated with multiple diseases such as ischemic and septic stroke. Luteolin (3,4,5,7-tetrahydroxyflavone) is a vascular protective flavonoid present in several dietary foods. However, how luteolin plays a role in microcirculatory disturbance is still unknown. The purpose of this study was to find out the influence of luteolin on the lipopolysaccharide (LPS)-induced microcirculatory disturbance, focusing on its effect on leukocyte adhesion and the underlying mechanism of this effect.
Methods
After injecting LPS into rats, we used an inverted intravital microscope to observe the velocity of red blood cells in venules, numbers of leukocytes adherent to and emigrated across the venular wall, hydrogen peroxide production in venular walls and mast cell degranulation. Intestinal microcirculation blood flow was measured by High-resolution Laser Doppler Perfusion Imaging. Histological changes of small intestine and mesenteric arteries were evaluated. Additionally, cell adhesion stimulated by LPS was tested on EA.hy926 and THP-1 cells. The production of pro-inflammatory cytokines, adhesion molecules and the activation of TLR4/Myd88/NF-κB signaling pathway were determined.
Results
The results showed luteolin significantly inhibited LPS-induced leukocyte adhesion, hydrogen peroxide production and mast cell degranulation, and increased intestinal microcirculation blood flow and ameliorated pathological changes in the mesenteric artery and the small intestine. Furthermore, luteolin inhibited the release of pro-inflammatory cytokines, the expression of TLR4, Myd88, ICAM-1, and VCAM-1, the phosphorylation of IκB-α and NF-κB/p65 in LPS stimulated EA.hy926.
Conclusions
Our findings revealed that it is likely that luteolin can ameliorate microcirculatory disturbance. The inhibitory effects of luteolin on the leukocyte adhesion stimulated by LPS, which participates in the development of microcirculatory disturbance, are mediated through the regulation of the TLR4/Myd88/NF-κB signaling pathway.
Supplementary Information
The online version contains supplementary material available at 10.1186/s12906-020-03196-9.
Keywords: Luteolin, LPS, Leukocyte adhesion, Microcirculatory disturbance
Background
Microcirculation is composed of arterioles, capillaries, and venules, functioning essentially as oxygen and source supply depending on the unique physiological need of the supplied organs [1]. It is composed of endothelial cells (ECs), basement membranes, pericytes, and smooth muscle cells. Microcirculatory disturbance, which can cause multiple organ damage, is a complex pathological process involving abnormal leukocyte-EC interactions, excessive production of peroxide, degranulated mast cells, and hyperpermeability of microvessel wall [2, 3]. It is also worth noting that leukocyte-EC interactions, especially leukocyte adhesion is a crucial step in microcirculatory disturbance processes.
The intact structure and optimal function of ECs ensure the normal microcirculation. The morphological characteristics of endothelial cells [4] and the expression of adhesion molecules determine that the leukocyte-EC interaction occurs more in venules [5, 6] than in arterioles [7]. The raised in leukocyte-EC interaction in inflamed vessels is identified as risk factors for thrombosis [8, 9]. Therefore, as a response to inflammatory stimulation, the dysfunction of venular EC may lead to thrombosis, stroke and other related diseases. In previous studies, venules have been found to be closely related to some diseases [10] such as ischemic stroke [11]. In the process of treating related diseases, improving the disturbance of microcirculation is an important measure to achieve a good prognosis.
Injection of Lipopolysaccharide (LPS) on animals is widely used as a model in the field of microcirculation disturbance research. LPS, located at the cell wall of gram-negative bacteria [9], can cause systemic inflammatory and septic shock accompanied with microcirculatory disturbance [12]. LPS could promote the expression of adhesion molecules [13], resulting in adhesion of leukocytes to ECs [14, 15]. Peroxides and proteases are produced when leukocytes adhere to the venular wall, which damages ECs and aggravate microvessel injury [16]. Moreover, these could also cause mast cells to degranulate and release pro-inflammatory cytokines [17]. These factors subsequently cause microcirculatory disturbance. Accordingly, suppression of adhesion of leukocytes and ECs is of great importance to improve LPS-induced microcirculatory disturbance.
Luteolin (3,4,5,7-tetrahydroxyflavone) is the main flavonoid contained in wild chrysanthemum and peanut shells [18, 19]. Moreover, it is one of the most common flavonoids in many dietetic foods, such as broccoli, olive oil, celery, parsley, green pepper, various herbs, dandelion, and Japanese honeysuckle [20]. It exhibits a variety of biological and pharmacological activities, including anti-inflammatory [21, 22], antioxidant, antihypertension [23], cardiovascular protection [24], anticancer and other activities. The improvement of vascular endothelial inflammation is widely considered to be the most significant advantage of luteolin. Vogl and coworkers reported that there was a decreasing expression of interleukin-8 (IL-8) and E-selectin protein caused by luteolin in ECs after LPS stimulation [25]. Luteolin could also protect against tumor necrosis factor-alpha (TNF-α)-induced vascular inflammation via interfering with NF-κB-mediated pathway [26].
In our preliminary experiment, we found that luteolin protects blood vessels by suppressing hypertensive vascular remodeling [23]. However, the role of luteolin in microcirculatory disturbance is still not clear and its underlying mechanism remains unexplored. Therefore, in this study, we employed a Sprague-Dawley (SD) rat inflammatory model induced by LPS to evaluate whether luteolin is capable of improving microcirculatory disturbance in rat mesenteric venules in vivo. Besides, we tried to elucidate its molecular mechanism of action with the LPS-induced EA.hy926 model in vitro. This research aimed to illuminate the mechanism of luteolin against microcirculatory disturbance and provide a theoretical basis for further development and utilization of luteolin.
Methods
Drug and chemicals preparation
Luteolin (LUT, purity>98%, L9283) and LPS were obtained from Sigma Chemical Co (St.Louis, MO, USA). We used DMSO to dissolve the compounds in vitro as the stock solution, which was stored at − 20 °C and diluted before use.
Luteolin enriched extracts (TLUT, purity>50%) were extracted from peanut shells according to the method described by Su et al. [23]. The peanut shells were purchased from New Nongdu Co., Ltd. (Hang Zhou, China).
Dihydrorhodamine 123 (DHR) was purchased from Molecular Probes Ltd. (Eugene, OR, USA). LPS (O55:B5), and toluidine blue were from Sigma Chemical Co. (St Louis, MO, USA).
Animals
Male Sprague-Dawley (SD) rats, with body weights ranging from 160 to 180 g, were provided by Experimental Animal Center of Zhejiang Province (Certificate no. SCXK 2014–0001, Hangzhou, China). They were acclimatized for 2 weeks before experiments. During this period, all of the rats were fed with standard diet and water. Animals were housed in a room with constant temperature, humidity, and light/dark cycle. All animal care and experimental procedures were approved by the Institutional Animal Care and Use Committee (IACUC) in the Zhejiang Chinese Medical University (permission number: ZSLL-2015-84).
Animal treatments and experimental design
Forty SD rats were randomly divided into four groups by SPSS statistics (version 17.0) software’s random number generator, and the experimental data analyses were conducted in a blinded manner. Group one (G1) was set as the control group. Rats injected with LPS and gavaged with distilled water, were designated as the LPS group (G2). Group three (G3) and group four (G4) were orally administered with TLUT (at the doses of 75 and 45 mg/kg, respectively) for 20 days in addition to LPS treatment.
The operation was similar to what Kai Sun performed [27]. Briefly, rats were anesthetized with an intraperitoneal injection of pentobarbital sodium (50 mg/kg) and then injected by tail vein with LPS (1 mg/kg) dissolved in saline while the rats of the control group were injected with saline alone. After that, the abdomen was opened by a midline blade (2–3 cm in length), and the ileocecal portion of the mesentery was mounted with care on a transparent plastic stage designed for the rat. The mesentery was kept warm in a constant temperature device and humidified by continuous dropping with saline at 37 °C. Microcirculatory hemodynamics in the mesentery was perceived by using a dynamic visualization of the microcirculation tracking system (Gene&I, Beijing, China). The single unbranched venules with 30 to 50 μm wide and about 200 μm long were selected for observation.
Intravital observation of mesenteric microcirculatory
The microcirculatory hemodynamics in rat mesentery was inspected and recorded continuously [28]. During the playback video images, the venular diameters at 20 min interval after 10 min of LPS injection was measured using Image-Pro Plus 6.0 software. The average diameter from three sites was calculated.
The movement of red blood cells (RBCs) was recorded (500 frames/sec) by a high-speed video camera system (Integrated Design Tools, Inc., USA). The velocity of RBC was analyzed by Image-Pro Plus 6.0 software. The average value was calculated from three different RBCs in the same time frame.
With the recorded videotaped images, we defined the rolling leukocytes as those that could be seen moving 10 s within the same segment of 200 μm long vessel. In addition, the adherent leukocytes were defined as those adhered to the same site for more than 10 s, and an average of 3 times of rolling and adherent leukocytes were counted.
To quantify the hydrogen peroxide production in venular wall, 10 μM DHR, a H2O2-sensitive fluorescent probe, was dropped to the mesenteric surface after dynamic observing 90 min. DHR fluorescence intensity on the venular wall (DB) and extra-vascular (DW) was measured using an image processor. DB-DW reflected the vascular wall superoxide production.
After DHR fluorescence examination, the mast cells in the mesentery were stained with 0.1% toluidine blue. Then we assessed the mast cell degranulation by averaging ratios of the number of degranulated mast cells to the total number of mast cells from the six captured pictures.
High-resolution laser doppler perfusion imaging
Intestinal microcirculation blood flow was evaluated by laser Doppler with a moorFLPI V3 software (Moor Instruments Ltd., UK) [29, 30]. After intravital observation of mesenteric microcirculation, the mesentery of rats was chosen to examine with an appropriate distance between the scanner and the mesentery of rats. The intensity of blood flow is presented with the different colors expanding from blue to green (low blood flow) and yellow to red (highest blood flow). Intestinal average microcirculation blood flow within each 15 s from three regions was determined by mFLPIReviewV3 image analysis software.
Histological evaluation
The rats were euthanized after completing the experiment, anesthetized with an intraperitoneal injection of pentobarbital sodium (150 mg/kg), approved methods for euthanasia by the Animal Care and Welfare Committee in the Zhejiang Chinese Medical University. The small intestine and mesenteric artery were excised and fixed in 10% formalin. After fixing, tissues were embedded in paraffin and cut into 4 μm thick sections. Which were stained with hematoxylin and eosin (Nanjing Jiangcheng Bioengineering Institute, Nanjing, China). Stained sections were visualized under the microscope (100X and 400X).
Cell culture
Human umbilical vein cell fusion cell (EA.hy926) with vascular endothelial cell characteristics (purchased from the Type Culture Collection of the Chinese Academy of Sciences, Shanghai, China) were maintained in Dulbecco’s modified Eagle’s medium (DMEM; Life Technologies Corporation, Beijing, China.), Human monocytic leukemia cell (THP-1; Type Culture Collection of the Chinese Academy of Sciences, Shanghai, China) were cultured in RPMI 1640 Medium (Gino Biological Pharmaceutical Co., Ltd. Hangzhou, China). Both mediums were supplemented with 10% fetal bovine serum (FBS) (Gemini Bio-Products, Australia.), 100 U/mL penicillin, 100 μg/mL streptomycin Cells were maintained in a 37 °C and 5% CO2 incubator.
Cytotoxicity assay
In the experiment, we used MTT to assess the viability of EA.hy926. EA.hy926cells were passaged and then inoculated into 96-well plates at a concentration of 1× 104/well 24 h after inoculation, cells were treated with different concentrations of LUT (0.1, 1, 5, 10, 20, 30 μM) in serum-free medium for another 24 h. The viability of cells was measured as described previously [23].
Cell adhesion assay
In order to evaluate the effect of luteolin on adhesive ability, we used LPS to stimulate EA.hy926 and THP-1 cells. Briefly, EA.hy926 cells were seeded at 1.5 × 104 /well in 96-well plates for 12 h. They were pre-treated with 5, 10, 20 μM LUT for 12 h, then co-incubated with 3 μg/mL LPS for 6 h. Meanwhile, THP-1 labeled with 5 μg/mL DAPI overnight was added into EA.hy926 plate (5 × 104 /well) and incubated for 30 min. Cells were carefully washed 3 times with PBS to remove non-adherent ones. Under the inverted fluorescence microscope (Olympus Corporation, Japan), nine random views were captured in each well at 200 × magnification and images were analyzed with Image-Pro Plus 5.1 software.
Enzyme-linked immunosorbent assay (ELISA)
EA.hy926 (1.5 × 104 each well) were grown in 96-well plates for 12 h and incubated with LUT at 5, 10, 20 μM and 3 μg/ml LPS for 24 h. Then cell supernatants were collected. TNF-α and IL-8 level were determined using ELISA kits (Shanghai Yuanye Biological pharmaceutical technology co., Ltd., Shanghai, China) according to the manufacturer’s protocol, respectively.
Cell protein extraction and western blotting analysis
The method resembled that of Jie Su [23]. In short, cells treated with various concentrations of LUT were collected. Cell proteins were extracted with RAPI buffer then separated by SDS-PAGE. Proteins were electro-transferred onto a polyvinylidene-difluoride membrane. After blocking with 5% BSA, the membranes were cultivated overnight at 4 °C with corresponding first antibodies against the NF-κBp65, p-NF-κBp65, Myd88, IκB-α, p-IκB-α (Cell Signaling Technology, Canada), toll-like receptor 4 (TLR4), intercellular adhesion molecule 1 (ICAM-1), vascular cell adhesion molecule 1 (VCAM-1) (Santa Cruz Biotechnology, USA). β-actin (Abcam, UK) was used as a loading control. Appropriate secondary antibody labeled with IRDye® 800CW (Li-cor, USA) were incubated with the membranes after TBST washes. Lastly, the protein bands were examined by LI-COR Odyssey CLx Infrared Imaging System.
Statistical analysis
All quantitative results were presented mean ± standard deviation. The differences between groups were analyzed by One-way analysis of variance (ANOVA) with LSD t-test or Dunnet t-test by SPSS 17.0 software. When homogeneity of variance assumptions is satisfied, the LSD t-test will be used; otherwise, when heterogeneity of variance assumptions is satisfied, the Dunnet t-test will be applied. The value of P< 0.05 was deemed to be statistically significant. Diagrams were generateded by Graph Prims.
Results
Effect of luteolin on the leukocyte adhesion to venular wall
The effect of luteolin on the adhesion between leukocytes and ECs was investigated both in vivo and in vitro. In vivo study, pictures of leukocyte adhesion to venular wall of each group at 10, 50, 90 min after LPS injection were presented in Fig.1a. Under various conditions, we did not observe adherent leukocytes in control group. In contrast, after LPS injection, numerous adherent leukocytes were observed in LPS group, while only a few adherent leukocytes in TLUT groups. Moreover, no adherent leukocyte was observed at 10 min after LPS injection in TLUT 75 mg/kg group, while still some adherent leukocytes remained at 10 min in TLUT 45 mg/kg group.
Fig. 1.

The effect of luteolin on LPS-induced leukocyte adhesion to the rat mesenteric venular wall. a Representative images illustrating the effect of luteolin on leukocyte adhesion to the venular wall induced by LPS in rat mesentery at 10, 50, 90 min after LPS injection, respectively (400X). Arrows: leukocyte adhesion to venular wall. b Time course of the number of leukocytes adhesion to the rat mesenteric venular wall. The data were expressed as means ± SEM (n = 6). Significant differences were indicated by #P< 0.05, ##P< 0.01 as compared with control group, *P< 0.05, **P< 0.01 as compared with LPS group
Furthermore, Fig.1b depicted the time course of leukocytes adhesion to the venular wall under different conditions, confirming the above qualitative evaluation. There were few adherent leukocytes in the control group, if any, throughout the whole observation period. Conversely, the number of adherent leukocytes is much higher in LPS group at each time point compared with control group (P< 0.01 or P< 0.05), and it reached the highest level of 18.6 ± 2.8 cells / 200 μm at 70 min after LPS injection. After LPS stimulation, the number of adherent leukocytes increased significantly from 10 min to 90 min. However, it declined significantly in groups treated with 75 mg/kg and 45 mg/kg TLUT (P< 0.01 or P< 0.05). Furthermore, the decrease was more dramatic in TLUT 75 mg/kg group than TLUT 45 mg/kg group. Impressively, treatment with TLUT resulted in marked inhibition of the LPS-induced leukocyte adhesion in vivo.
To prove the effect of luteolin on cell adhesion in vitro, we examined the cell adhesion in EA.hy926 and THP-1 cells. First of all, we detected the cytotoxicity of luteolin on EA.hy926 cells. As presented in Fig.2c, Compared with control group, it had a significant inhibition of the cell viability of EA.hy926 after treatment with LUT at 30 μM (P< 0.01). However, there was no marked difference between other groups and the control group in cell viability. The results turned out that cytotoxicity of luteolin was at a concentration of up to 30 μM. Therefore, LUT at 5 μM, 10 μM, and 20 μM were used in subsequent experiments. As shown in Fig.2b, EA.hy926 stimulated by 3 μg/mL LPS attracted more THP-1 cells in comparison with the control group (P< 0.01). The numbers of LPS-induced adherent cells significantly decreased when EA.hy926 cells were incubated 24 h with 5 μM, 10 μM, and 20 μM LUT (P< 0.01 or P< 0.05).
Fig. 2.

The effect of luteolin on LPS-induced cell adhesion between EA.hy926 and THP-1. a Representative images of cell adhesion between EA.hy926 and THP-1. Pictures were captured at 200x magnification. b A quantitative evaluation of adherent cells. The data were expressed as means ± SEM (n = 9). Significant differences were indicated by ##P< 0.01 as compared with the control group, *P< 0.05, **P< 0.01 as compared with the LPS group. c The effect of luteolin on cell viability in EA.hy926. The data were expressed as means ± SEM (n = 12). Significant differences were indicated by ##P< 0.01 as compared with the control group
In conclusion, luteolin could significantly inhibit leukocyte adhesion to the venular wall in vivo and in vitro induced by LPS, demonstrate a direct inhibitory effect on leukocytes adhesion to the endothelium.
Effect of luteolin on the leukocyte rolling along venular wall
As illustrated in Table 1, the number of rolling leukocytes along a 200 μm venular wall in the control group was less than five per 10s. Compared with the control group, the number of rolling leukocytes increased in the LPS group which became most significant at 10 min and 30 min after LPS injection, but there was no significant difference in statistics. Moreover, the data showed that there was no significant statistical difference between the TLUT group and the LPS group at any time point, but it was found that 10 and 30 min after the injection of LPS, the treatment with TLUT could alleviate the rolling leukocyte increase induced by LPS to some extent, but this effect was not conspicuous.
Table 1.
The effect of luteolin on leukocyte rolling along venular wall
| Groups | Time after LPS injection (min) | ||||
|---|---|---|---|---|---|
| 10 | 30 | 50 | 70 | 90 | |
| Control | 1.9±1.2 | 1.7±1.1 | 1.8±1.0 | 1.6±1.1 | 2.4±1.9 |
| LPS 1 mg/kg | 9.1±4.4 | 5.2±2.1 | 3.8±3.5 | 2.4±1.8 | 2.2±2.0 |
| TLUT 75 mg/kg | 5.4±4.2 | 4.2±2.4 | 5.0±4.1 | 3.8±2.0 | 5.9±3.3 |
| TLUT 45 mg/kg | 4.4±3.2 | 3.7±2.1 | 4.1±2.0 | 3.7±2.2 | 4.7±1.9 |
The data were expressed as means ± SEM (n = 6)
Effect of luteolin on the hydrogen peroxide production in the venular wall
As presented in Fig.3a, almost no detectable DHR fluorescence in rat mesenteric venular wall was observed in the control group. In contrast, the DHR fluorescence intensity increased impressively in the LPS group after 90 min of LPS injection (P< 0.01) and was high to 12.4 ± 1.5. However, treatment with 75 mg/kg and 45 mg/kg TLUT markedly inhibited LPS-induced hydrogen peroxide production (P< 0.01 or P< 0.05), and the DHR fluorescence intensity in TLUT 75 mg/kg group was low to 4.6 ± 2.4.
Fig. 3.

The effect of luteolin on LPS-induced hydrogen peroxide production, mast cell degranulation and intestinal microcirculation blood flow. a The effect of luteolin on LPS-induced hydrogen peroxide production. Pictures were captured at 400x magnification. Arrows: DHR fluorescence on the venular walls. b The effect of luteolin on LPS-induced mast cell degranulation. Pictures were captured at 400x magnification. Arrows: degranulated mast cell. c The effect of luteolin on LPS-stimulated intestinal microcirculation blood flow. All of the representative images of each group were presented with the respective quantification showing right correspondingly. All of the data were expressed as means ± SEM (n = 6). Significant differences were indicated by ##P< 0.01 as compared with control group, *P< 0.05, **P< 0.01 as compared with LPS group
Effect of luteolin on the mast cell degranulation
Mast cell degranulation after 90 min of LPS injection was analyzed and expressed as the ratio of the number of degranulated mast cells to the total number of mast cells, as shown in Fig.3b. In the control group, degranulated mast cells were very small. After LPS injection, the percentage of degranulated mast cells significantly increased (P< 0.01), which was high to 71.9% ± 19.8%. However, treatment with 75 mg/kg and 45 mg/kg TLUT markedly suppressed the LPS-induced degranulation of mast cells (P< 0.01). The value was 35.0% ± 8.9 and 42.2% ± 6.6%, respectively.
Effect of luteolin on the intestinal microcirculation blood flow and RBC velocity
We measured the intestinal microcirculation blood flow by the laser Doppler perfusion imaging. Figure 3c illustrates the intestinal microcirculation blood flow, which was examined after observing mast cell degranulation. The control SD rats had the adequate blood flow. On the contrary, the blood flow in the LPS group impressively declined (P< 0.01), which was low to 431.1 ± 62.2 PU. However, the blood flow was apparently increased with the treatment of 75 mg/kg and 45 mg/kg TLUT (P< 0.01). The value was 554.2 ± 27.2 PU and 511.8 ± 42.9 PU, respectively.
As is known, the blood flow velocity is a direct factor affecting blood flow. Thus we further examined the RBC velocity in venules by a high-speed video camera system. Table 2 demonstrated the period in which RBC velocity changes in mesenteric venules. The velocity of RBC in LPS group progressively declined at 10 min and 30 min after LPS injection in comparison with the control group (P< 0.01 or P< 0.05). However, This LPS-induced decrease in RBC velocity was without significant attenuated by treatment with TLUT. In addition, saline injection led to a progressive decrease in RBC velocity, and a remarkable difference was observed 50 min, 70 min and 90 min after saline injection compared with 10 min (P< 0.01 or P< 0.05).
Table 2.
The effect of luteolin on RBC velocity in mesenteric venules of rats injected with LPS (mm/s)
| Groups | Time after LPS injection (min) | ||||
|---|---|---|---|---|---|
| 10 | 30 | 50 | 70 | 90 | |
| Control | 2.96±0.60 | 2.57±0.44 | 2.02±0.47$$ | 2.15±0.58$ | 1.98±0.56$$ |
| LPS 1 mg/kg | 1.35±0.34## | 1.41±0.31# | 1.39±0.26 | 1.37±0.31 | 1.25±0.28 |
| TLUT 75 mg/kg | 1.65±0.66 | 1.70±0.96 | 1.82±0.89 | 1.72±0.88 | 1.55±0.96 |
| TLUT 45 mg/kg | 1.38±0.77 | 1.41±0.85 | 1.10±0.34 | 1.04±0.47 | 1.07±0.34 |
The data were expressed as means ± SEM (n = 6); #P< 0.05 vs. control group;##P< 0.01 vs. control group;$P< 0.05 vs. 10 min;$$P< 0.01 vs. 10 min
The data showed that luteolin could improve intestinal microcirculation blood flow suppressed by LPS, but it had no significant effect on RBC velocity, suggesting that the effect of luteolin on improving intestinal microcirculation blood flow is not related to RBC velocity.
Effect of luteolin on the histology of mesenteric artery and small intestine
The HE staining for rat mesenteric artery and small intestine were presented in Fig.4. As shown in Fig.4a, in the LPS group, membrane cells of the mesenteric artery had the irregular arrangement, and inflammatory cells infiltrated compared with the control group. Moreover, Fig.4b illustrates the situation of the disintegration of intestinal villi existed in the LPS group. However, it is comfortable that the situation was ameliorated by 75 mg/kg and 45 mg/kg TLUT. Especially, the effect in 75 mg/kg TLUT group was more significant.
Fig. 4.
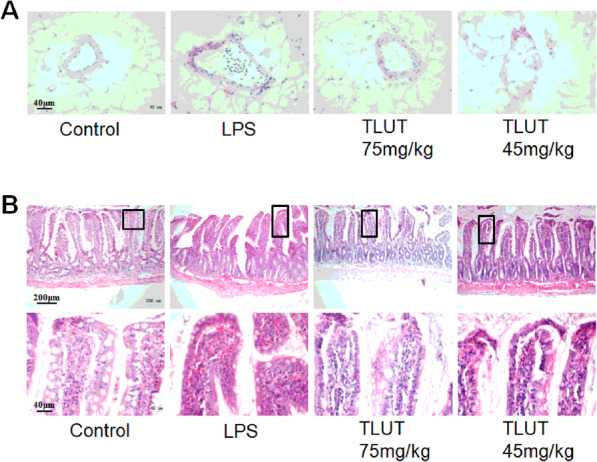
The effect of luteolin on LPS-stimulated mesenteric artery and small intestine pathological changes. a Representative images of HE staining of the mesenteric artery in each group (400X). b Typical images of HE staining of the small intestine in each group (100X). The area in the rectangle of each image was enlarged and shown below correspondingly (400X)
Effect of luteolin on the LPS/TLR4/NF-κB signaling pathway in EA.hy926
TLR4 participated in the signaling transduction induced by LPS, which led to the activation of NF-κB. NF-κB, a ubiquitous transcription factor, controls the expression of a large number of adhesion molecules and pro-inflammatory cytokines. In order to explore the anti-adhesion mechanisms of luteolin, we examined the expression of related factors of LPS/TLR4/NF-κB signaling pathway in EA.hy926 stimulated with LPS by western blot, including TLR4, Myd88, ICAM-1, VCAM-1 and activation of IκB-α and NF-κB/p65. The results of western blot were illustrated in Fig. 5. The expression of TLR4, Myd88, ICAM-1 and VCAM-1 were increased in LPS stimulated EA.hy926 cells. While treatment of LUT markedly reduced their expression, especially in the high concentration of LUT at 20 μM.
Fig. 5.

The effect of luteolin on LPS/TLR4/NF-κB pathway. a The TNF-α and IL-8 level in cultural supernatants of EA.hy926 cells. The data were expressed as means ± SEM (n = 8). Significant differences were indicated by ##P< 0.01 as compared with control group, **P< 0.01 as compared with LPS group. b The expression of TLR4, Myd88, IκB-α, NF-κB/p65, ICAM-1 and VCAM-1, and activation of IκB-α and NF-κB/p65 in EA.hy926 cells
Phosphorylation of IκB-α through proteolytic degradation leads to the nuclear translocation of NF-κB. Exposure to LPS increased phosphorylation of IκB-α and p65 in the cells. As expected, treatment with LUT decreased the LPS-induced phosphorylation of IκB-α and p65.
Moreover, to evaluate the effect of luteolin on the production of pro-inflammatory cytokines in EA.hy926 cells, we tested the level of TNF-α and IL-8 in cell supernatants using ELISA kits. As presented in Fig. 5a, the TNF-α and IL-8 secreted from EA.hy926 cells in LPS group were apparently more abundant than that in the control group (P< 0.01). As expected, 5 μM, 10 μM, and 20 μM LUT markedly diminished LPS-stimulated TNF-α and IL-8 level in cell supernatants (P< 0.01).
Taken together, LPS could increase TLR4 expression in EA.hy926 cells and promote the activation of its downstream signaling pathways. Our data suggest that luteolin decreased the expression of adhesion molecules and pro-inflammatory cytokines may through inhibiting LPS/TLR4/NF-κB signaling pathway.
Discussion
Microcirculation plays a crucial role in supplying oxygen and nutrients by blood flow to the tissues to meet their metabolic demands. Many studies have found that microcirculation disturbance is one of the leading causes of multiple diseases. Luteolin (3′, 4′, 5, 7-tetrahydroxyflavone), a kind of flavonoid found in nature, is enriching in many natural plants and widely presented in certain food items, especially in olive, honey and broccoli, providing an essential link between diet and prevention of chronic disease [18]. Previous studies have reported that luteolin may be a vascular protective agent by directly acting on ECs, and it prevents vascular dysfunction likely associated with antioxidant and anti-inflammatory mechanisms [31], and has the vasorelaxant effect through NO production [32]. In our preliminary experiment, we found that luteolin suppressed hypertensive vascular remodeling to protect blood vessels and inhibited TNF-α-induced vascular endothelial inflammation, suggesting luteolin may have a potential effect for improving microcirculatory disturbance.
LPS shredded from Gram-negative bacteria is a factor for microcirculation disturbance, which started with leukocyte recruitment in the microvessels. Leukocyte recruitment is controlled by adhesion molecules on leukocytes and ECs [27]. LPS could activate and up-regulate the expression of adhesion molecules from several families, resulting in rolling and adhesion of leukocytes to ECs [33]. Firstly, leukocytes rolling along the vessel wall is caused by the activation of endothelial and leukocytic selectins, whereas subsequent activation of ICAM-1 and VCAM-1 on ECs and CD11b, CD18 on leukocytes leads to the firm adhesion of leukocytes on the endothelial surface [34, 35]. In addition, the release of pro-inflammatory cytokines such as TNF-α, IL-6 and IL-8 could be stimulated by LPS [27, 36], which further exaggerates the adhesion between leukocytes and ECs followed by leukocytes extravasation and inflammatory reaction. The release of ROS [37] and mast cell degranulation [35] are promoted by the interactions between leukocytes and ECs, which exacerbates microcirculatory disturbance. Accordingly, inhibiting the adhesion between leukocytes and ECs is regarded to be the key to alleviating the disturbance of microcirculation induced by LPS.
Injection of LPS on animals is widely used as a model in the field of microcirculation disturbance research. With this model, the present experiment studied the effects of luteolin on the microcirculatory disturbance in vivo by examining the venular diameter, RBC velocity, leukocyte rolling, leukocyte adhesion, hydrogen peroxide production, mast cell degranulation, intestinal microcirculation blood flow and so on. Additionally, to identify the molecular mechanisms, the expression of adhesion molecules and pro-inflammatory cytokines were studied in an in vitro experiment.
The results from the present study demonstrated that luteolin could significantly inhibit leukocyte adhesion to the venular wall in LPS-induced rats although has no significant effect on venular diameter, RBC velocity and leukocytes rolling. Meanwhile, luteolin could also suppress the adhesion of monocytes to human EA.hy926 ECs in vitro induced by LPS, implying an direct effect of luteolin on inhibiting the leukocytes adhesion to the endothelium. As mentioned above, leukocytes adhesion to endothelium is the key step to inducing microcirculatory disturbance by LPS stimulation. This process is controlled by adhesion molecules on ECs and leukocytes. Furthermore, pro-inflammatory cytokines could exaggerate leukocyte adhesion.
Our results showed that luteolin attenuates the LPS-induced enhancement of adhesion molecules ICAM-1 and VCAM-1 expression in EA.hy926 cells, which was accordant with the reported effect of luteolin in inhibiting the expression of ICAM-1 and VCAM-1 in TNF-α-induced EA.hy926 cells [26]. In addition, some researchers found that luteolin significantly inhibited the expression of VCAM-1 and ICAM-1 in intestinal epithelial cells [38] or pulmonary microvascular ECs [39]. Moreover, luteolin markedly diminished LPS-stimulated TNF-α and IL-8 levels in EA.hy926 cell supernatants. In conclusion, luteolin protects against LPS-induced adhesion between leukocytes and ECs, in both in vitro and in vivo models. This effect of luteolin may be mediated by inhibiting the expression of adhesion molecules and the secretion of pro-inflammatory cytokines from ECs. However, the effect of luteolin on other adhesion molecules and pro-inflammatory cytokines remains to be explored.
It is widely known that LPS-enhanced adhesion molecules expression and pro-inflammatory cytokines secretion are primarily mediated by TLR4/NF-κB signaling pathway [13]. TLR4 is not only an important member of the TLR family but also a receptor for LPS. Myd88 is an indispensable component of the signaling cascade of TLR4 [40]. Activation of TLR4 can trigger downstream Myd88-dependent signaling pathway [41]. The final event is the activation of NF-κB, then the activated NF-κB protein transfers from cytoplasm to nucleus and binds to DNA reaction elements in gene promoter region to promote the gene transcription [42]. The phosphorylation of IκB typically gets involved in the activation of NF-κB. IκB, an inhibitor of NF-κB, is known to mainly bind to P65/P50 heterodimer in the cytoplasm [43, 44]. It is the phosphorylation of IκB that results in its ubiquitylation and subsequent degradation, which allows the release of NF-κB and its translocation to nucleus. Eventually, activation of the NF-κB leads to the production of cell adhesion molecules, such as ICAM-1 and VCAM-1 [45], and secretion of pro-inflammatory cytokines [27, 46], such as TNF-α and IL-8. These cytokines will then cause adhesion between leukocytes and ECs.
Western blot was used to analyze the expression of molecules in TLR4/NF-κB signaling pathway. Our results showed that LPS increased TLR4 expression in EA.hy926 cells and promoted the activation of its downstream signaling pathways. Luteolin could inhibit the expression of TLR4 and Myd88, which consequently suppressed activation of IκB and NF-κB. To further investigate the effect of luteolin on activation of IκB and NF-κB, IκB-α and NF-κB/p65 protein and their phosphorylated form were examined after LPS stimulation. We found that luteolin obviously inhibited LPS-induced phosphorylation and degradation of IκB-α and the activation of NF-κB/p65. In addition, it was reported that luteolin could improve TNF-α-induced vascular inflammation by inhibiting the NF-κB-mediated pathway [26]. Some researchers found that luteolin attenuated the activation of NF-κB and AP-1 in LPS-induced RAW 264.7 cells [47], and ameliorated LPS-mediated nephrotoxicity via decreasing NF-κB activation [48]. These indicated that luteolin could decrease the expression of adhesion molecules and the secretion of pro-inflammatory cytokines by inhibiting TLR4/NF-κB signaling pathway.
The interaction between leukocytes and ECs promotes ROS production [49]. ROS, markedly by the treatment of lute including hydrogen peroxide, causes cytotoxicity in ECs and induces pro-inflammatory cytokine production, which subsequently exacerbates microcirculatory disturbance. DHR fluorescence intensity was detected to quantify the effect of luteolin on hydrogen peroxide production in venular wall. The results showed that luteolin markedly inhibited LPS-induced hydrogen peroxide production on venular wall. Previous studies have reported that luteolin could protect against H2O2-induced oxidative stress and ameliorated ROS and superoxide generation in HUVECs, and prevent endothelial dysfunction induced by oxidative stress [50]. Therefore, the results of this study indicate that the improvement of luteolin on LPS-induced microcirculatory disorders is not only by inhibition of adhesion between leukocytes and ECs but also by scavenging ROS.
It is acknowledged that LPS makes mast cells degranulate, which leads to the release of cytokines such as histamine and TNF-α. These cytokines launch an extravascular attack on microvessels, promote adhesion molecules expression on ECs, and subsequently enhance leukocytes adhesion and venular hyperpermeability. In the present study, luteolin markedly decreased the percent of LPS-induced degranulated mast cells, suggesting that luteolin can protect against the microcirculatory disturbance by blockage of vasoactive substances from outside. Moreover, luteolin could improve the reduction of intestinal microcirculation blood flow, and it may be associated with the inhibition of leukocyte adhesion. The inhibition of leukocyte adhesion may enlarge the vascular lumen and increase the blood flow. Luteolin also could prevent the inflammatory infiltrates in the mesenteric artery, small intestine edema and fluff disintegration. These pathological changes are all involved in microcirculatory disturbance.
To sum up, this study used the rat inflammation model induced by LPS to evaluate the effect of luteolin on microcirculation disturbance. The results revealed that luteolin could significantly ameliorate microcirculatory disturbance. Interestingly, we found that luteolin could significantly inhibit the leukocyte adhesion to the venular wall, but it had no significant effect on leukocyte rolling. The results of further cell adhesion experiments in vitro were consistent with those in vivo. Thus, we focused on the modulation of the adhesion between leukocytes and ECs and its possible mechanism. As well accepted, LPS-enhanced adhesion molecules expression and pro-inflammatory cytokines secretion, which leads to the firm adhesion of leukocytes to the endothelial surface, are primarily mediated by TLR4/NF-κB signaling pathway. Our results further showed that luteolin could inhibit the expression of TLR4 and Myd88, phosphorylation of IκB-α and NF-κB/p65, and the expression of ICAM-1, VCAM-1, TNF-α and IL-8, suggesting luteolin could decrease the expression of adhesion molecules and pro-inflammatory cytokines by inhibiting LPS/TLR4/NF-κB signaling pathway. These results are consistent with the anti-inflammatory effects of luteolin previously reported.
Microcirculatory disturbance induced by LPS is a complex pathological process, which may initiates with leukocyte-EC interaction, followed by reactive oxygen species production, mast cell degranulation, microvessel hyperpermeability, albumin extravasation and so on. These processes could interact. However, leukocytes adhesion to endothelial cells is a crucial contribution to the development of microcirculatory disturbance. Based on the results of our study, we speculated luteolin improved LPS-induced microcirculation disturbance possibly by inhibiting leukocyte adhesion through the regulation of LPS/TLR4/NF-κB signaling pathway, as shown in Fig.6. However, due to existing deficiencies such as limited methods and data, more researches are needed to corroborate this mechanism of luteolin, and other mechanisms remain to be explored.
Fig. 6.

The proposed mechanism of luteolin on suppressing microcirculatory disturbance caused by cell adhesion. Luteolin could decrease the expression of adhesion molecules and pro-inflammatory cytokines by inhibiting LPS/TLR4/NF-κB signaling pathway, which would suppress LPS-induced leukocyte adhesion. Subsequently, the other processes of microcirculatory disturbance induced by LPS such as hydrogen peroxide production, mast cell degranulation, blood flow reduced and mesenteric artery pathological changes were also improved
Conclusions
Our studies concluded that luteolin inhibited the LPS-induced activation of the TLR4/Myd88/NF-κB signaling pathway. It down-regulated the expression of TLR4, Myd88 and the phosphorylation of IκB-a and NF-κB, inhibited the expression of cell adhesion molecules including ICAM-1 and VCAM-1, and restrained pro-inflammatory cytokine production including TNF-α and IL-8. Furthermore, luteolin suppressed LPS-induced adhesion between leukocytes and ECs, as well as hydrogen peroxide production. In addition, luteolin inhibited the venular hyperpermeability, decreased mast cell degranulation, increased intestinal microcirculation blood flow, and improved mesenteric artery and small intestine pathological changes. Finally, luteolin ameliorated microcirculatory disturbance. The findings in our studies indicate that luteolin is a potential inhibitor of cell adhesion, and may have potential application in treating microcirculatory disturbance.
Supplementary Information
Acknowledgements
Not applicable.
Abbreviations
- LPS
Lipopolysaccharide
- TLR4
Toll-like receptor 4
- ICAM-1
Intercellular adhesion molecule 1
- VCAM-1
Vascular cell adhesion molecule 1
- TNF-α
Tumor necrosis factor-alpha
- NF-κB
Nuclear factor-kappa B
- MyD88
Myeloid differentiation factor 88
- IκB
Inhibitor of Nuclear factor-kappa B
- IL-8
Interleukin-8
- LUT
Luteolin
- TLUT
Luteolin enriched extracts
- DHR
Dihydrorhodamine 123
- RBC
Red blood cell
- EA.hy926
Human umbilical vein cell fusion cell
- THP-1
Human monocytic leukemia cell
Authors’ contributions
GYL and SHC conceived and designed the study. JS, HTX, TW, YJW and CW performed the experiments. BL and SSL contributed to the analysis of data. JS and HTX wrote the manuscript. JJY, MQY, WJL and SMC reviewed and edited the manuscript. All authors read and approved the manuscript.
Funding
This research was supported by the Zhejiang Provincial Science Foundation of China under Grant No.LQ18H280003, the National Science Foundation of China under Grant No.82003977, No.81873036 and No.81803819, the Key Research and Development Project of Zhejiang province (No.2017C03052), the Zhejiang Provincial Key Laboratory Project (No.2012E10002) and Opening Project of Zhejiang Provincial First-rate Subject (Chinese Traditional Medicine) (No. Ya 2017010).
Availability of data and materials
The data used and/or investigated during the present study are accessible from the corresponding author on reasonable request.
Ethics approval and consent to participate
All animal care and experimental procedures were approved by the Institutional Animal Care and Use Committee (IACUC) at the Zhejiang Chinese Medical University (permission number: ZSLL-2015-84).
Consent for publication
Not applicable.
Competing interests
The authors declare no conflict of interest.
Footnotes
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Jie Su and Han-Ting Xu contributed equally to this work.
References
- 1.Colbert JF, Schmidt EP. Endothelial and Microcirculatory Function and Dysfunction in Sepsis. Clin Chest Med. 2016;37(2):263–275. doi: 10.1016/j.ccm.2016.01.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Sun K, Wang CS, Guo J, Horie Y, Fang SP, Wang F, et al. Protective effects of ginsenoside Rb1, ginsenoside Rg1, and notoginsenoside R1 on lipopolysaccharide-induced microcirculatory disturbance in rat mesentery. Life Sci. 2007;81(6):509–518. doi: 10.1016/j.lfs.2007.06.008. [DOI] [PubMed] [Google Scholar]
- 3.Davies T, Wythe S, O'Beirne J, Martin D, Gilbert-Kawai E. Review article: the role of the microcirculation in liver cirrhosis. Aliment Pharmacol Ther. 2017;46(9):825–835. doi: 10.1111/apt.14279. [DOI] [PubMed] [Google Scholar]
- 4.Sumagin R, Sarelius IH. Intercellular adhesion molecule-1 enrichment near tricellular endothelial junctions is preferentially associated with leukocyte transmigration and signals for reorganization of these junctions to accommodate leukocyte passage. J Immunol. 2010;184(9):5242–5252. doi: 10.4049/jimmunol.0903319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Kim MB, Sarelius IH. Role of shear forces and adhesion molecule distribution on P-selectin-mediated leukocyte rolling in postcapillary venules. Am J Physiol Heart Circ Physiol. 2004;287(6):H2705–H2711. doi: 10.1152/ajpheart.00448.2004. [DOI] [PubMed] [Google Scholar]
- 6.Kubes P, Kanwar S. Histamine induces leukocyte rolling in post-capillary venules. A P-selectin-mediated event. J Immunol. 1994;152(7):3570–3577. [PubMed] [Google Scholar]
- 7.Sumagin R, Sarelius IH. A role for ICAM-1 in maintenance of leukocyte-endothelial cell rolling interactions in inflamed arterioles. Am J Physiol Heart Circ Physiol. 2007;293(5):H2786–H2798. doi: 10.1152/ajpheart.00720.2007. [DOI] [PubMed] [Google Scholar]
- 8.Afshar-Kharghan V, Thiagarajan P. Leukocyte adhesion and thrombosis. Curr Opin Hematol. 2006;13(1):34–39. doi: 10.1097/01.moh.0000190107.54790.de. [DOI] [PubMed] [Google Scholar]
- 9.Zhang JH, Shangguan ZS, Chen C, Zhang HJ, Lin Y. Anti-inflammatory effects of guggulsterone on murine macrophage by inhibiting LPS-induced inflammatory cytokines in NF-kappaB signaling pathway. Drug Des Devel Ther. 2016;10:1829–1835. doi: 10.2147/DDDT.S104602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Hartmann DA, Hyacinth HI, Liao FF, Shih AY. Does pathology of small venules contribute to cerebral microinfarcts and dementia? J Neurochem. 2018;144(5):517–526. doi: 10.1111/jnc.14228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Mutlu U, Swanson SA, Klaver CCW, Hofman A, Koudstaal PJ, Ikram MA, et al. The mediating role of the venules between smoking and ischemic stroke. Eur J Epidemiol. 2018;33(12):1219–1228. doi: 10.1007/s10654-018-0436-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Richards EM, Pepine CJ, Raizada MK, Kim S. The Gut, Its Microbiome, and Hypertension. Curr Hypertens Rep. 2017;19(4):36. doi: 10.1007/s11906-017-0734-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Lu YC, Yeh WC, Ohashi PS. LPS/TLR4 signal transduction pathway. Cytokine. 2008;42(2):145–151. doi: 10.1016/j.cyto.2008.01.006. [DOI] [PubMed] [Google Scholar]
- 14.Darley-Usmar V, Halliwell B. Blood radicals: reactive nitrogen species, reactive oxygen species, transition metal ions, and the vascular system. Pharm Res. 1996;13(5):649–662. doi: 10.1023/A:1016079012214. [DOI] [PubMed] [Google Scholar]
- 15.Han JY, Horie Y, Miura S, Akiba Y, Guo J, Li D, et al. Compound Danshen injection improves endotoxin-induced microcirculatory disturbance in rat mesentery. World J Gastroenterol. 2007;13(26):3581–3591. doi: 10.3748/wjg.v13.i26.3581. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Kurose I, Argenbright LW, Wolf R, Lianxi L, Granger DN. Ischemia/reperfusion-induced microvascular dysfunction: role of oxidants and lipid mediators. Am J Physiol. 1997;272(6 Pt 2):H2976–H2982. doi: 10.1152/ajpheart.1997.272.6.H2976. [DOI] [PubMed] [Google Scholar]
- 17.Kulka M, Sheen CH, Tancowny BP, Grammer LC, Schleimer RP. Neuropeptides activate human mast cell degranulation and chemokine production. Immunology. 2008;123(3):398–410. doi: 10.1111/j.1365-2567.2007.02705.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Lopez-Lazaro M. Distribution and biological activities of the flavonoid luteolin. Mini Rev Med Chem. 2009;9(1):31–59. doi: 10.2174/138955709787001712. [DOI] [PubMed] [Google Scholar]
- 19.Qiu J, Chen L, Zhu Q, Wang D, Wang W, Sun X, et al. Screening natural antioxidants in peanut shell using DPPH-HPLC-DAD-TOF/MS methods. Food Chem. 2012;135(4):2366–2371. doi: 10.1016/j.foodchem.2012.07.042. [DOI] [PubMed] [Google Scholar]
- 20.Nabavi SF, Braidy N, Gortzi O, Sobarzo-Sanchez E, Daglia M, Skalicka-Wozniak K, et al. Luteolin as an anti-inflammatory and neuroprotective agent: A brief review. Brain Res Bull. 2015;119(Pt A):1–11. doi: 10.1016/j.brainresbull.2015.09.002. [DOI] [PubMed] [Google Scholar]
- 21.Gu J, Cheng X, Luo X, Yang X, Pang Y, Zhang X, et al. Luteolin ameliorates cognitive impairments by suppressing the expression of inflammatory cytokines and enhancing synapse-associated proteins GAP-43 and SYN levels in Streptozotocin-induced diabetic rats. Neurochem Res. 2018;43(10):1905–1913. doi: 10.1007/s11064-018-2608-6. [DOI] [PubMed] [Google Scholar]
- 22.Aziz N, Kim M, Cho J. Anti-inflammatory effects of luteolin: A review of in vitro, in vivo, and in silico studies. J Ethnopharmacol. 2018;225:342–358. doi: 10.1016/j.jep.2018.05.019. [DOI] [PubMed] [Google Scholar]
- 23.Su J, Xu HT, Yu JJ, Gao JL, Lei J, Yin QS, et al. Luteolin Ameliorates Hypertensive Vascular Remodeling through Inhibiting the Proliferation and Migration of Vascular Smooth Muscle Cells. Evid Based Complement Alternat Med. 2015;2015:364876. doi: 10.1155/2015/364876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Lv GY, Zhang YP, Gao JL, Yu JJ, Lei J, Zhang ZR, et al. Combined antihypertensive effect of luteolin and buddleoside enriched extracts in spontaneously hypertensive rats. J Ethnopharmacol. 2013;150(2):507–513. doi: 10.1016/j.jep.2013.08.058. [DOI] [PubMed] [Google Scholar]
- 25.Vogl S, Atanasov AG, Binder M, Bulusu M, Zehl M, Fakhrudin N, et al. The Herbal Drug Melampyrum pratense L. (Koch): Isolation and Identification of Its Bioactive Compounds Targeting Mediators of Inflammation. Evid Based Complement Alternat Med. 2013;2013:395316. doi: 10.1155/2013/395316. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Jia Z, Nallasamy P, Liu D, Shah H, Li JZ, Chitrakar R, et al. Luteolin protects against vascular inflammation in mice and TNF-alpha-induced monocyte adhesion to endothelial cells via suppressing IKappaBalpha/NF-kappaB signaling pathway. J Nutrit Biochem. 2015;26(3):293–302. doi: 10.1016/j.jnutbio.2014.11.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Sun K, Wang CS, Guo J, Liu YY, Wang F, Liu LY, et al. Effect of Panax notoginseng saponins on lipopolysaccharide-induced adhesion of leukocytes in rat mesenteric venules. Clin Hemorheol Microcirc. 2006;34(1–2):103–108. [PubMed] [Google Scholar]
- 28.Yuan Q, Liu YY, Sun K, Chen CH, Zhou CM, Wang CS, et al. Improving effect of pretreatment with yiqifumai on LPS-induced microcirculatory disturbance in rat mesentery. Shock (Augusta, Ga) 2009;32(3):310–316. doi: 10.1097/SHK.0b013e31819c36e8. [DOI] [PubMed] [Google Scholar]
- 29.Stucker M, Esser M, Hoffmann M, Memmel U, Hirschmuller A, von Bormann C, et al. High-resolution laser Doppler perfusion imaging aids in differentiating between benign and malignant melanocytic skin tumours. Acta Derm Venereol. 2002;82(1):25–29. doi: 10.1080/000155502753600849. [DOI] [PubMed] [Google Scholar]
- 30.Esser D, Oosterink E, op ‘t Roodt J, Henry RM, Stehouwer CD, Muller M, et al. Vascular and inflammatory high fat meal responses in young healthy men; a discriminative role of IL-8 observed in a randomized trial. PloS one. 2013;8(2):e53474. doi: 10.1371/journal.pone.0053474. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Si H, Wyeth RP, Liu D. The flavonoid luteolin induces nitric oxide production and arterial relaxation. Eur J Nutr. 2014;53(1):269–275. doi: 10.1007/s00394-013-0525-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Gentile D, Fornai M, Pellegrini C, Colucci R, Benvenuti L, Duranti E, et al. Luteolin Prevents Cardiometabolic Alterations and Vascular Dysfunction in Mice With HFD-Induced Obesity. Front Pharmacol. 2018;9:1094. doi: 10.3389/fphar.2018.01094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Kim S, Joo YE. Theaflavin Inhibits LPS-Induced IL-6, MCP-1, and ICAM-1 Expression in Bone Marrow-Derived Macrophages Through the Blockade of NF-kappaB and MAPK Signaling Pathways. Chonnam Med J. 2011;47(2):104–110. doi: 10.4068/cmj.2011.47.2.104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Xu H, Wang D, Peng C, Huang X, Ou M, Wang N, et al. Rabbit sera containing compound danshen dripping pill attenuate leukocytes adhesion to TNF-alpha--activated human umbilical vein endothelial cells by suppressing endothelial ICAM-1 and VCAM-1 expression through NF-kappaB signaling pathway. J Cardiovasc Pharm. 2014;63(4):323–332. doi: 10.1097/FJC.0000000000000046. [DOI] [PubMed] [Google Scholar]
- 35.Davenpeck KL, Zagorski J, Schleimer RP, Bochner BS. Lipopolysaccharide-induced leukocyte rolling and adhesion in the rat mesenteric microcirculation: regulation by glucocorticoids and role of cytokines. J Immunol. 1998;161(12):6861–6870. [PubMed] [Google Scholar]
- 36.Li A, Dong L, Duan ML, Sun K, Liu YY, Wang MX, et al. Emodin improves lipopolysaccharide-induced microcirculatory disturbance in rat mesentery. Microcirculation (New York, NY : 1994) 2013;20(7):617–628. doi: 10.1111/micc.12061. [DOI] [PubMed] [Google Scholar]
- 37.Wan BY, Peh KH, Ho M, Assem ES. Effects of nitric oxide and hydrogen peroxide on histamine release from RBL-2H3 cells. Biochem Pharmacol. 2001;62(11):1537–1544. doi: 10.1016/S0006-2952(01)00770-5. [DOI] [PubMed] [Google Scholar]
- 38.Kim JS, Jobin C. The flavonoid luteolin prevents lipopolysaccharide-induced NF-kappaB signalling and gene expression by blocking IkappaB kinase activity in intestinal epithelial cells and bone-marrow derived dendritic cells. Immunology. 2005;115(3):375–387. doi: 10.1111/j.1365-2567.2005.02156.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Kong X, Huo G, Liu S, Li F, Chen W, Jiang D. Luteolin suppresses inflammation through inhibiting cAMP-phosphodiesterases activity and expression of adhesion molecules in microvascular endothelial cells. Inflammopharmacology. 2019;27(4):773–780. doi: 10.1007/s10787-018-0537-2. [DOI] [PubMed] [Google Scholar]
- 40.De Nardo D. Toll-like receptors: activation, signalling and transcriptional modulation. Cytokine. 2015;74(2):181–189. doi: 10.1016/j.cyto.2015.02.025. [DOI] [PubMed] [Google Scholar]
- 41.Cronin JG, Turner ML, Goetze L, Bryant CE, Sheldon IM. Toll-like receptor 4 and MYD88-dependent signaling mechanisms of the innate immune system are essential for the response to lipopolysaccharide by epithelial and stromal cells of the bovine endometrium. Biol Reprod. 2012;86(2):51. doi: 10.1095/biolreprod.111.092718. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Joh EH, Kim DH. Lancemaside A inhibits lipopolysaccharide-induced inflammation by targeting LPS/TLR4 complex. J Cell Biochem. 2010;111(4):865–871. doi: 10.1002/jcb.22773. [DOI] [PubMed] [Google Scholar]
- 43.Xu D, Yan S, Wang H, Gu B, Sun K, Yang X, et al. IL-29 Enhances LPS/TLR4-Mediated Inflammation in Rheumatoid Arthritis. Cell Physiol Biochem. 2015;37(1):27–34. doi: 10.1159/000430330. [DOI] [PubMed] [Google Scholar]
- 44.Mobarak E, Haversen L, Manna M, Rutberg M, Levin M, Perkins R, et al. Glucosylceramide modifies the LPS-induced inflammatory response in macrophages and the orientation of the LPS/TLR4 complex in silico. Sci Rep. 2018;8(1):13600. doi: 10.1038/s41598-018-31926-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Liu L, Li YH, Niu YB, Sun Y, Guo ZJ, Li Q, et al. An apple oligogalactan prevents against inflammation and carcinogenesis by targeting LPS/TLR4/NF-kappaB pathway in a mouse model of colitis-associated colon cancer. Carcinogenesis. 2010;31(10):1822–1832. doi: 10.1093/carcin/bgq070. [DOI] [PubMed] [Google Scholar]
- 46.Zhang J, Li Q, Shao Q, Song J, Zhou B, Shu P. Effects of panax notoginseng saponin on the pathological ultrastructure and serum IL-6 and IL-8 in pulmonary fibrosis in rabbits. J Cell Biochem. 2018;119(10):8410–8418. doi: 10.1002/jcb.27045. [DOI] [PubMed] [Google Scholar]
- 47.Park CM, Song YS. Luteolin and luteolin-7-O-glucoside inhibit lipopolysaccharide-induced inflammatory responses through modulation of NF-kappaB/AP-1/PI3K-Akt signaling cascades in RAW 264.7 cells. Nutr Res Pract. 2013;7(6):423–429. doi: 10.4162/nrp.2013.7.6.423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Xin SB, Yan H, Ma J, Sun Q, Shen L. Protective Effects of Luteolin on Lipopolysaccharide-Induced Acute Renal Injury in Mice. Med Sci Monit. 2016;22:5173–5180. doi: 10.12659/MSM.898177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Guo J, Sun K, Wang CS, Fang SP, Horie Y, Yang JY, et al. Protective effects of dihydroxylphenyl lactic acid and salvianolic acid B on LPS-induced mesenteric microcirculatory disturbance in rats. Shock (Augusta, Ga) 2008;29(2):205–211. doi: 10.1097/SHK.0b013e318070c61a. [DOI] [PubMed] [Google Scholar]
- 50.Yang SC, Chen PJ, Chang SH, Weng YT, Chang FR, Chang KY, et al. Luteolin attenuates neutrophilic oxidative stress and inflammatory arthritis by inhibiting Raf1 activity. Biochem Pharmacol. 2018;154:384–396. doi: 10.1016/j.bcp.2018.06.003. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The data used and/or investigated during the present study are accessible from the corresponding author on reasonable request.