

Biofilms are increasingly implicated as playing a major role in waterborne cryptosporidiosis. This review aims to synthesize all currently available data on interactions between Cryptosporidium oocysts and biofilms.
KEYWORDS: biofilms, Cryptosporidium, waterborne pathogens
ABSTRACT
Biofilms are increasingly implicated as playing a major role in waterborne cryptosporidiosis. This review aims to synthesize all currently available data on interactions between Cryptosporidium oocysts and biofilms. Initially described following a waterborne outbreak, the integration of Cryptosporidium oocysts in biofilm has been well demonstrated. Biofilms appear important in the dissemination/protection of oocysts in the environment. Consequently, it has been suggested that substrate-associated biofilms should be systematically considered in oocyst water quality assessment. The influence of physicochemical parameters has been studied on oocyst biofilm retention. Biofilm surface roughness, ionic concentration (especially Ca2+), laminar/turbulent flow, shear stress, and electrostatic repulsion forces appear important to consider regarding oocyst release from biofilm. However, data analysis carried out during this review also revealed important gaps in biological interactions within biofilms, offering many perspectives for future work.
INTRODUCTION
Cryptosporidium spp. are anthropo-zoonotic parasitic protozoa belonging to the phylum Apicomplexa. The genus Cryptosporidium was recently classified in the subclass Cryptogregaria in the class of Gregarinomorphea (1). Cryptosporidiosis is currently classified as an emerging disease (2), and Cryptosporidium has been reported as the second most implicated pathogen (in both morbidity and mortality) in infants with diarrhea in developing countries (3). Several outbreaks of Cryptosporidium spp. have been reported. One of the main waterborne outbreaks occurred in 1993 in Milwaukee, Wisconsin, United States, where the drinking water network was contaminated, leading to more than 400,000 infections in humans and 69 deaths (4). More recently, between 2011 and 2016, at least 381 outbreaks related to protozoan waterborne transmission were described (63% due to Cryptosporidium). However, these outbreaks mainly occurred in New Zealand (49%) and North America (41%), while notifications from Europe represented 9% of them and there were almost none from regions of endemicity (Africa, Asia, and South America) (5). These data highlight huge variations in surveillance systems. Many parasitic outbreaks have occurred worldwide, especially in developing countries, but without any notification. An effective surveillance system requires an associated budget and clearly stated guidelines. Many differences are observed from country to country and pathogen to pathogen with a better focus on bacteria and viruses than parasites. Consequently, many protozoan parasitic waterborne outbreaks occur worldwide, but they are not reported because of a lack of parasitological investigations, diagnosis difficulties, or a lack of official declarations.
Cryptosporidium foodborne outbreaks are documented less often than waterborne ones and also are probably widely underestimated. Many reasons are given, including underreporting, lack of national/international food safety standards, and difficulties in tracing back to food items due to prolonged incubation time for cryptosporidiosis (around 7 to 10 days). However, foodborne outbreaks of cryptosporidiosis are increasingly reported thanks to the development of surveillance systems such as FoodNet, National Notifiable Diseases Surveillance System, European Centre for Disease Prevention and Control, etc. Cryptosporidium oocysts have already been reported as responsible for outbreaks from fruits (apples, cider, etc.), vegetables (salads, spinach, etc.) dairy products, meat, and shellfish (6). Several factors could explain the frequent implication of Cryptosporidium in outbreaks, including (i) the low 50% infective dose (ID50). ID50 was calculated at 132 oocysts in healthy volunteers for Cryptosporidium parvum (7) and was estimated between 10 and 83 oocysts for Cryptosporidium hominis (8) even if a single C. hominis oocyst could cause an infection (1, 8–10); and (ii) resistance to disinfectant. Oocysts are highly resistant to chlorine, allowing a high number to escape into wastewater treatment devices (11, 12). All considered, the impact of cryptosporidiosis on the environment is important, and special attention should be given to prevent this pathology since no real therapeutic options are available for infected humans or animals. Consequently, understanding the survival and dissemination of Cryptosporidium oocysts is crucial. Among potential environmental sources of contamination, the role of biofilm must be considered.
A biofilm is defined as a microbial community irreversibly associated with a surface and enclosed in an extracellular polymeric substance matrix (13). Many microbiological waterborne outbreaks, especially bacterial ones, have been linked to biofilms. For example, Legionella was reported as the most frequently reported etiology among drinking water-associated outbreaks, and Legionella replication occurs in free-living amoebae that are particularly encountered in biofilm. Pseudomonas aeruginosa is also frequently implicated in outbreaks due to water contamination and especially due to the development of biofilms in water distribution systems (14–16). In a recent publication, C. hominis was encountered in a freshwater sponge, underlining the importance of considering substrate-associated biofilms in natural aquatic environments (17). In the literature, some data are available regarding Cryptosporidium implicated in biofilms (mono- or polymicrobial) but without any overview. Consequently, we propose here a state-of-the-art review of current knowledge on Cryptosporidium spp. implicating biofilms.
Physicochemical interactions.
(i) Cryptosporidium oocysts and biofilm attachment ability. The ability of Cryptosporidium oocysts to attach to biofilm was first suggested after a cryptosporidiosis waterborne outbreak that occurred in northwest England in March 2000. The outbreak control team suggested changing the water supplying the area affected by the outbreak to remove any oocyst water contamination. However, oocysts remained microscopically detectable in water for up to 19 days after the change. Authors hypothesized oocyst release from biofilm developed on the surface of distribution pipes (18). Evidence of potential Cryptosporidium oocyst distribution in biofilm was first provided by Skraber et al., showing the presence of oocysts in polymicrobial biofilms developed on polyvinylchloride (PVC) coupons submerged for at least 2 months in wastewater (19). Recently, Masangkay et al. compared the prevalence of oocyst detection in substrate-associated biofilms, with detection in both surface and bottom water with sediment. They observed a higher oocyst prevalence in substrate-associated biofilms and concluded that water quality assessment strategies could be strongly improved by systematically considering substrate-associated biofilms for environmental oocyst screening (20). In 2009, Wolyniak et al. reproduced biofilms in single-channel flow chambers by scraping biofilms naturally developed on rocks in creek waters. Flow chambers were previously incubated with biofilm cells for 24 h before starting a constant laminar flow. Influent water was supplemented in C. parvum oocysts for 3 days and was free of oocyst during several days to evaluate the sloughing of oocysts from the biofilm. Results show the ability of oocysts to integrate biofilm. Oocysts rapidly reached a steady state after inoculation and decreased to a new steady state within 5 days when oocyst supply was removed (21, 22). Oocyst biofilm integration was saturated; no increase in the number of oocysts attached to the biofilm was observed when the loading rate was increased from 104 to 105 oocysts/day (11). Helmi et al. studied oocyst attachment and detachment from two developed natural biofilms. After 7 months of developing biofilms on polycarbonate coupons placed inside a rotating annular reactor fed with drinking water or wastewater under laminar regiment, 106 C. parvum oocysts were spiked in the reactor, and oocyst biofilm attachment and detachment were monitored in time. Most attachment occurred within the first hour of biofilm exposure to oocysts. Oocysts were released from the biofilm during the entire experiment over 34 days (23). In contrast, Searcy et al. reported no significant release of oocysts from the biofilms after initial deposition. Searcy et al. developed biofilms under laminar conditions in flow cells fed with P. aeruginosa grown in Jensen's medium or Luria-Bertani broth. C. parvum oocysts were injected after obtaining confluent P. aeruginosa biofilms. No significant difference in the number of oocysts occurring in biofilms was observed 24 h after oocyst injection compared to 1 h or even after a 40-fold increase in flow velocity. Microbial species constituting biofilms probably strongly influence oocyst release; however, Searcy et al. did not look for oocysts released in effluent water, which could have changed their conclusions of no oocyst release from biofilms (24). The main characteristics, strengths, and weaknesses of each model are summarized in Table 1.
TABLE 1.
Summary of characteristics of biofilm reported in in vitro models
| Reactor type | Culture media | Oocyst inclusion | Strengths of the model | Weaknesses of the model | Reference no. |
|---|---|---|---|---|---|
| Flow cells | Creek water | Added to influent water after 24 h of biofilm cell incubation | Dynamic, polymicrobial, and representative of environmental conditions | Uncharacterized microorganisms, unidirectional flow, and oocysts added after biofilm development | 21, 22 |
| Flow cells | Both Jensen's and Luria-Bertani broths | Spiked after P. aeruginosa confluence | Dynamic model, well-characterized biofilm | Not representative of environmental conditions, monomicrobial, unidirectional flow, oocysts added after biofilm development | 24 |
| Rotating disk reactor | Both drinking water and wastewater | Spiked after 7 mo of running | Dynamic model, turbulent flow, representative of environmental conditions | Oocysts added after biofilm development, uncharacterized microorganisms | 23 |
(ii) Biofilm thickness and oocyst attachment location. Wolyniak et al. first observed high oocyst attachment to summer biofilm cultures, which was correlated with the highest thickness of developed biofilms (22). In a subsequent complementary study, Wolyniak et al. also observed a statistically higher biofilm thickness in biofilms developed from summer sampling than winter sampling, but it was not correlated with an increase in the number of oocysts attached to the biofilm. Finally, they concluded that it was unlikely that biofilm thickness explained increased oocyst attachment because all oocysts were observed to attach at the biofilm surface, and no oocysts were ever observed within biofilm depths (21, 22).
Regarding methods, Wolyniak et al. introduced oocysts after at least 24 h of previous biofilm growth, which could at least partially explain why oocysts were always observed in superficial layers of biofilms. However, no difference in oocyst attachment location was observed if oocysts were added to the water supply during all of the experiment or only after a few days (22). In addition, Searcy et al. also always observed oocysts at the biofilm surface, introducing oocysts after obtaining a confluent P. aeruginosa biofilm (24). The hypothesis put forward was that the inner biofilm was denser, with less pore space, while the biofilm surface had more water channels, providing more surface area for particle attachment. Consequently, large microorganisms such as Cryptosporidium oocysts could more easily integrate the surface of biofilm than the deep layers (22). Searcy et al. also investigated the influence of extracellular polymeric substances on oocyst attachment to biofilm. In developing P. aeruginosa biofilms from both a wild-type strain and a strain overproducing exopolysaccharide (EPS) alginate, no difference in oocyst retention was observed, suggesting no influence of EPS (24). However, such conclusions were obtained by introducing oocysts after the development of confluent biofilms from the two strains, and results could be different when introducing oocysts at the beginning of the experiment. EPS structure strongly depends on constitutive microorganisms, and further investigations considering polymicrobial communities could modify interpretations.
(iii) Biofilm roughness influence. Rather than thickness, oocyst retention appeared strongly correlated with biofilm surface roughness (11, 24). The explanation appears to be physical since a rough surface provides more surface area for attachment. Okabe et al. proved this hypothesis studying fluorescent microparticle diffusion in mixed biofilms; polymicrobial biofilms were developed on disks immersed in wastewater (or synthetic culture media) in a rotating disk reactor; tracer beads were added to young, intermediate, and mature biofilms, and tracer bead spatial distribution was measured microscopically by sectioning the biofilm. Results show that an increased surface roughness provides more surface area for bead attachment (25). Wu et al. studied the adhesion of C. parvum oocysts to a large number of polymeric materials. They observed an optimal adhesion of oocysts to low-surface-roughness materials (close to the surface roughness of oocysts) and suggested that, as already known for bacteria, irregularities that conform to the size of oocysts could increase the adhesion due to maximizing parasite-surface contact area (26).
(iv) Water parameter influence. Based on a preliminary study showing higher oocyst attachment from summer biofilm cultures than winter ones, Wolyniak et al. evaluated the influence of water temperature on C. parvum oocyst biofilm attachment (21, 22). Regarding the cumulative percentage of oocysts associated with the biofilm (grown at 5°C versus 20 to 25°C), temperature did not affect oocyst attachment. However, temperature influenced the growth of microorganisms and, consequently, the microbial biofilm composition. It has been shown that an increasing temperature results in a decreasing concentration of dissolved oxygen in water, also probably influencing microbial biofilm composition (27). The physical characteristics of the biofilm (roughness, thickness, etc.) also depend on constituting microorganisms. Consequently, temperature could indirectly influence oocyst attachment due to microorganism selection, even if such evidence needs further investigation (21). The influence of biofilm oocyst attachment was also investigated for other water parameters. Neither pH, conductivity, or dissolved organic carbon concentration appeared significantly correlated with oocyst attachment to biofilm, even if a trend of decreasing oocyst attachment with increasing pH was observed (21). Electrostatic forces have been proposed to explain, at least in part, this pH influence. C. parvum oocysts are neutral around pH 2 to 3 and become increasingly negatively charged as the pH increases (28). Biofilm surfaces are generally negatively charged as a result of the presence of anionic carboxyl, sulfate, and phosphoryl groups in the biofilm matrix (21, 28). Consequently, the more pH increases, the more negative repulsion forces are reinforced, probably boosting oocyst detachment. Mechanical forces probably also play a role in the observed effect of increased pH; Simoes et al. showed the facilitation of biofilm removal after sodium hydroxide exposure due to mechanical action (29). In addition, Dai et al. evaluated the effects of biofilm and natural organic matter (NOM) on the removal of Cryptosporidium parvum oocysts from water by filtration and reported less efficient removal of oocysts from biofilm-coated bed and NOM exposure. Hypotheses were (i) pore clogging and short-circuiting of the filter bed by the biofilm; (ii) increased negative charge of oocysts due to NOM, leading to enhanced electrostatic repulsion forces; and (iii) hydrophobic interactions with oocysts (30).
(v) Ion-mediated oocyst attachment. In developing biofilms on polycarbonate coupons fed with stream water, C. parvum oocyst attachment appeared independent of solution ionic strength, little influenced by magnesium but strongly influenced by calcium. Luo et al. demonstrated that the attachment of oocysts to biofilms responds to a pseudo-second-order calcium-mediated process; calcium was identified as the predominant factor that bridges the carboxylic functional groups on biofilm and oocyst surfaces to cause attachment (31). Sarkhosh et al. confirmed that oocyst binding was related to a calcium-mediated mechanism and added that it was dependent on oocyst age; the binding strength increases over a 5-week period, followed by a decrease in binding strength. Oocyst wall structure probably changes over time; the outer layer is mainly composed of glycoproteins and glucose-rich glycocalyx, whereas the inner layer is composed of glycolipids and lipoproteins. Ion-mediated interactions appeared variable over time, probably associated with oocyst wall structure modifications, but more characterization is needed (32).
(vi) Role of laminar/turbulent flow and wall shear stress. Some observations have been reported in the literature regarding the influence of flow and shear stress on oocyst detachment. First, Searcy et al. reported no release of oocysts from biofilm, even after a 40-fold increase in the laminar flow rate (2.5 ml/min). However, in their experiment, they developed a confluent Pseudomonas aeruginosa biofilm before oocyst loading, and finally, they observed no statistically significant detachment of Pseudomonas cells at the higher flow rate. They concluded that the maximum flow rate achieved in their experiment was insufficient to generate sufficient shear stress to observe microorganism release from biofilm (24). Helmi et al. observed a massive transfer of parasites from biofilm to water phase after increasing the flow rate from a laminar to turbulent regimen (23). Helmi et al. developed biofilms from drinking water and wastewater origins; biofilms were developed in a rotating annular reactor for 7 months or 14 days with a flow rate of 100 ml/min. Such variations in methods could explain differences in observed conclusions on oocyst release from biofilm. Finally, Luo et al. evaluated the impact of hydrodynamic conditions on oocyst attachment to biofilms developed in an annular rotating reactor from stream water sampling. They showed a different influence of shear stress on oocyst attachment over time; higher shear (to a certain limit) may be associated with faster initial oocyst attachment, but lower shear is associated with greater numbers of oocysts attached at equilibrium (33).
Biological interactions.
(i) Biofilm oocyst replication. The development of C. parvum in host-cell free culture was first reported by Hijijawi et al. in 2004 and was based on microscopy (34). Based on quantitative PCR (qPCR), flow cytometry, and microscopic observations, Koh et al. reported that biofilm can support the multiplication of Cryptosporidium over time in an aquatic environment (35, 36). They developed P. aeruginosa biofilms during 24 h before continuously adding C. parvum oocysts into an influent medium (60 ml/h; 106 oocysts/24 h) for 6 days. Various Cryptosporidium developmental stages (sporozoites, trophozoites, type I and II meronts) were identified from the biofilm, suggesting that Cryptosporidium can complete much of its life cycle in a biofilm. Regarding qPCR, C. parvum DNA was detected targeting glyceraldehyde-3-phosphate dehydrogenase (GAPDH) gene. They observed a 2- to 3-fold multiplication of C. parvum DNA after 6 days when oocysts were exposed to biofilm versus without biofilm exposure. However, the production of new oocysts in aquatic biofilms has not been reported, and the observed C. parvum multiplication targeting the GAPDH gene could be discussed because the observed overexpression of the GAPDH gene could be explained by an increase in the metabolic activity of oocysts when exposed to biofilm. Finally, the ability of Cryptosporidium to replicate in biofilm needs further investigation.
(ii) Effect of UV on oocysts embedded in biofilm. UV irradiation on Cryptosporidium oocyst viability has already been reported in the literature, but biofilm influence is rarely documented (37–40). DiCesare et al. compared (i) the infectivity of oocysts attached to the top of a biofilm, (ii) the infectivity of oocysts attached to the bottom of a 30-μm biofilm, and (iii) the infectivity of resuspended oocysts. Sun-exposed oocysts were less infectious than oocysts in the dark. The infectivity of oocysts attached to the top of the biofilm was significantly less than the infectivity of oocysts at the bottom of the biofilm, suggesting that the biofilm provides a protective barrier for oocysts against solar radiation. The density of the biofilm could influence solar protection as well as the microbial community constituting the biofilm; the ability of the biofilm to absorb UV wavelengths may be a result of microbial production of sunscreen-like compounds (41).
Currently, biological investigations on oocysts embedded in biofilm are very limited. Many questions remain, especially on public health impact and outbreak prevention. How long do oocysts stay infectious in biofilm? Is oocyst release from biofilm a risk for human health? Are there any interactions with microorganisms constituting biofilms? Are natural microorganism predators, such as free-living amoebae, able to phagocyte oocysts? Are environmental disinfection methods, such as chlorine disinfection, ozone therapy etc., effective on biofilm, or should they be associated with antibiofilm strategies (flush, anti-EPS, impregnated nanoparticles, etc.)? Does mineral rock/soil composition play a role in biofilm architecture/properties? Many aspects remain to be studied.
In conclusion, the integration of Cryptosporidium oocysts in biofilm has been clearly demonstrated. It has also been suggested that substrate-associated biofilms should be systematically considered for environmental oocyst investigations. The release of oocysts from biofilm has also been clearly demonstrated and seems dependent on several factors (Fig. 1). Some physicochemical parameters have been studied. Among them, roughness, ion-mediated attachment (especially Ca2+), and microorganisms constituting the biofilm appeared important considering oocyst integration and release from the biofilm. However, much work remains to be done, especially considering the microbial composition of biofilm and polymicrobial interactions. In addition, data analysis within the framework of this review has clearly demonstrated gaps in biological investigations between Cryptosporidium oocysts and biofilm integration. Consequently, future works on oocyst biofilm interactions are expected for a better understanding of the potential infectious risk for humans and its management.
FIG 1.
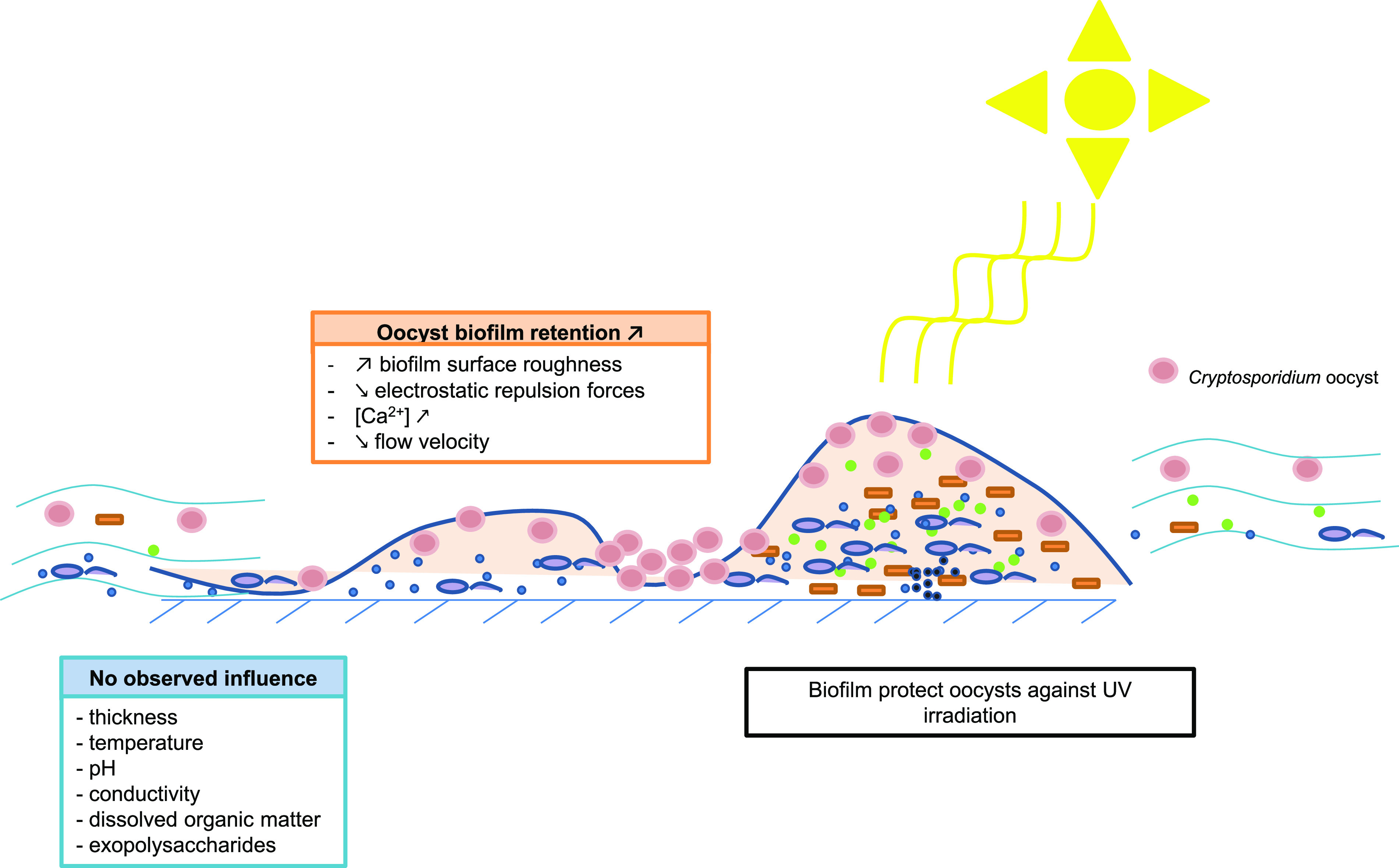
Schematic representation of parameters influencing oocyst retention/dissemination from biofilms.
ACKNOWLEDGMENT
We are grateful to Nikki Sabourin-Gibbs, Rouen University Hospital, for her help in editing the manuscript.
REFERENCES
- 1.Ryan U, Paparini A, Monis P, Hijjawi N. 2016. It’s official – Cryptosporidium is a gregarine: what are the implications for the water industry? Water Res 105:305–313. doi: 10.1016/j.watres.2016.09.013. [DOI] [PubMed] [Google Scholar]
- 2.Bamaiyi PH, Redhuan NEM. 2017. Prevalence and risk factors for cryptosporidiosis: a global, emerging, neglected zoonosis. Asian Biomed 10. doi: 10.5372/1905-7415.1004.493. [DOI] [Google Scholar]
- 3.Kotloff KL, Nataro JP, Blackwelder WC, Nasrin D, Farag TH, Panchalingam S, Wu Y, Sow SO, Sur D, Breiman RF, Faruque AS, Zaidi AK, Saha D, Alonso PL, Tamboura B, Sanogo D, Onwuchekwa U, Manna B, Ramamurthy T, Kanungo S, Ochieng JB, Omore R, Oundo JO, Hossain A, Das SK, Ahmed S, Qureshi S, Quadri F, Adegbola RA, Antonio M, Hossain MJ, Akinsola A, Mandomando I, Nhampossa T, Acácio S, Biswas K, O'Reilly CE, Mintz ED, Berkeley LY, Muhsen K, Sommerfelt H, Robins-Browne RM, Levine MM. 2013. Burden and aetiology of diarrhoeal disease in infants and young children in developing countries (the Global Enteric Multicenter Study, GEMS): a prospective, case-control study. Lancet Lond Engl 382:209–222. doi: 10.1016/S0140-6736(13)60844-2. [DOI] [PubMed] [Google Scholar]
- 4.Mac Kenzie WR, Schell WL, Blair KA, Addiss DG, Peterson DE, Hoxie NJ, Kazmierczak JJ, Davis JP. 1995. Massive outbreak of waterborne Cryptosporidium infection in Milwaukee, Wisconsin: recurrence of illness and risk of secondary transmission. Clin Infect Dis 21:57–62. doi: 10.1093/clinids/21.1.57. [DOI] [PubMed] [Google Scholar]
- 5.Efstratiou A, Ongerth JE, Karanis P. 2017. Waterborne transmission of protozoan parasites: review of worldwide outbreaks - an update 2011–2016. Water Res 114:14–22. doi: 10.1016/j.watres.2017.01.036. [DOI] [PubMed] [Google Scholar]
- 6.Ryan U, Hijjawi N, Xiao L. 2018. Foodborne cryptosporidiosis. Int J Parasitol 48:1–12. doi: 10.1016/j.ijpara.2017.09.004. [DOI] [PubMed] [Google Scholar]
- 7.DuPont HL, Chappell CL, Sterling CR, Okhuysen PC, Rose JB, Jakubowski W. 1995. The infectivity of Cryptosporidium parvum in healthy volunteers. N Engl J Med 332:855–859. doi: 10.1056/NEJM199503303321304. [DOI] [PubMed] [Google Scholar]
- 8.Chappell CL, Tzipori S, Akiyoshi DE, Okhuysen PC, Tanriverdi S, Langer-Curry R, Widmer G. 2006. Cryptosporidium hominis: experimental challenge of healthy adults. Am J Trop Med Hyg 75:851–857. doi: 10.4269/ajtmh.2006.75.851. [DOI] [PubMed] [Google Scholar]
- 9.Benamrouz S, Guyot K, Gazzola S, Mouray A, Chassat T, Delaire B, Chabé M, Gosset P, Viscogliosi E, Dei-Cas E, Creusy C, Conseil V, Certad G. 2012. Cryptosporidium parvum infection in SCID mice infected with only one oocyst: qPCR assessment of parasite replication in tissues and development of digestive cancer. PLoS One 7:e51232. doi: 10.1371/journal.pone.0051232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Chappell CL, Okhuysen PC, Langer-Curry RC, Akiyoshi DE, Widmer G, Tzipori S. 2011. Cryptosporidium meleagridis: infectivity in healthy adult volunteers. Am J Trop Med Hyg 85:238–242. doi: 10.4269/ajtmh.2011.10-0664. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Wolyniak DiCesare EA, Hargreaves BR, Jellison KL. 2012. Biofilm roughness determines Cryptosporidium parvum retention in environmental biofilms. Appl Environ Microbiol 78:4187–4193. doi: 10.1128/AEM.08026-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Angles ML, Chandy JP, Cox PT, Fisher IH, Warnecke MR. 2007. Implications of biofilm-associated waterborne Cryptosporidium oocysts for the water industry. Trends Parasitol 23:352–356. doi: 10.1016/j.pt.2007.06.001. [DOI] [PubMed] [Google Scholar]
- 13.Donlan RM. 2002. Biofilms: microbial life on surfaces. Emerg Infect Dis 8:881–890. doi: 10.3201/eid0809.020063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Benedict KM, Reses H, Vigar M, Roth DM, Roberts VA, Mattioli M, Cooley LA, Hilborn ED, Wade TJ, Fullerton KE, Yoder JS, Hill VR. 2017. Surveillance for waterborne disease outbreaks associated with drinking water - United States, 2013–2014. MMWR Morb Mortal Wkly Rep 66:1216–1221. doi: 10.15585/mmwr.mm6644a3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Costa D, Bousseau A, Thevenot S, Dufour X, Laland C, Burucoa C, Castel O. 2015. Nosocomial outbreak of Pseudomonas aeruginosa associated with a drinking water fountain. J Hosp Infect 91:271–274. doi: 10.1016/j.jhin.2015.07.010. [DOI] [PubMed] [Google Scholar]
- 16.Hota S, Hirji Z, Stockton K, Lemieux C, Dedier H, Wolfaardt G, Gardam MA. 2009. Outbreak of multidrug-resistant Pseudomonas aeruginosa colonization and infection secondary to imperfect intensive care unit room design. Infect Control Hosp Epidemiol 30:25–33. doi: 10.1086/592700. [DOI] [PubMed] [Google Scholar]
- 17.Masangkay FR, Milanez GD, Tsiami A, Somsak V, Kotepui M, Tangpong J, Karanis P. 2020. First report of Cryptosporidium hominis in a freshwater sponge. Sci Total Environ 700:134447. doi: 10.1016/j.scitotenv.2019.134447. [DOI] [PubMed] [Google Scholar]
- 18.Howe AD, Forster S, Morton S, Marshall R, Osborn KS, Wright P, Hunter PR. 2002. Cryptosporidium oocysts in a water supply associated with a cryptosporidiosis outbreak. Emerg Infect Dis 8:619–624. doi: 10.3201/eid0806.010271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Skraber S, Helmi K, Willame R, Ferréol M, Gantzer C, Hoffmann L, Cauchie HM. 2007. Occurrence and persistence of bacterial and viral faecal indicators in wastewater biofilms. Water Sci Technol 55:377–385. doi: 10.2166/wst.2007.280. [DOI] [PubMed] [Google Scholar]
- 20.Masangkay FR, Milanez GD, Tsiami A, Hapan FZ, Somsak V, Kotepui M, Tangpong J, Karanis P. 2020. Waterborne protozoan pathogens in environmental aquatic biofilms: implications for water quality assessment strategies. Environ Pollut 259:113903. doi: 10.1016/j.envpol.2019.113903. [DOI] [PubMed] [Google Scholar]
- 21.Wolyniak EA, Hargreaves BR, Jellison KL. 2010. Seasonal retention and release of Cryptosporidium parvum oocysts by environmental biofilms in the laboratory. Appl Environ Microbiol 76:1021–1027. doi: 10.1128/AEM.01804-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Wolyniak EA, Hargreaves BR, Jellison KL. 2009. Retention and release of Cryptosporidium parvum oocysts by experimental biofilms composed of a natural stream microbial community. Appl Environ Microbiol 75:4624–4626. doi: 10.1128/AEM.02916-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Helmi K, Skraber S, Gantzer C, Willame R, Hoffmann L, Cauchie H-M. 2008. Interactions of Cryptosporidium parvum, Giardia lamblia, vaccinal poliovirus type 1, and bacteriophages X174 and MS2 with a drinking water biofilm and a wastewater biofilm. Appl Environ Microbiol 74:2079–2088. doi: 10.1128/AEM.02495-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Searcy KE, Packman AI, Atwill ER, Harter T. 2006. Capture and retention of Cryptosporidium parvum oocysts by Pseudomonas aeruginosa biofilms. Appl Environ Microbiol 72:6242–6247. doi: 10.1128/AEM.00344-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Okabe S, Yasuda T, Watanabe Y. 1997. Uptake and release of inert fluorescence particles by mixed population biofilms. Biotechnol Bioeng 53:459–469. doi: 10.1002/(SICI)1097-0290(19970305)53:5<459::AID-BIT3>3.0.CO;2-G. [DOI] [PubMed] [Google Scholar]
- 26.Wu M, Bridle H, Bradley M. 2012. Targeting Cryptosporidium parvum capture. Water Res 46:1715–1722. doi: 10.1016/j.watres.2011.12.041. [DOI] [PubMed] [Google Scholar]
- 27.Moss JA, Nocker A, Lepo JE, Snyder RA. 2006. Stability and change in estuarine biofilm bacterial community diversity. Appl Environ Microbiol 72:5679–5688. doi: 10.1128/AEM.02773-05. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Kuznar ZA, Elimelech M. 2004. Adhesion kinetics of viable Cryptosporidium parvum oocysts to quartz surfaces. Environ Sci Technol 38:6839–6845. doi: 10.1021/es0494104. [DOI] [PubMed] [Google Scholar]
- 29.Simões M, Pereira MO, Vieira MJ. 2005. Effect of mechanical stress on biofilms challenged by different chemicals. Water Res 39:5142–5152. doi: 10.1016/j.watres.2005.09.028. [DOI] [PubMed] [Google Scholar]
- 30.Dai X, Hozalski RM. 2002. Effect of NOM and biofilm on the removal of Cryptosporidium parvum oocysts in rapid filters. Water Res 36:3523–3532. doi: 10.1016/s0043-1354(02)00045-3. [DOI] [PubMed] [Google Scholar]
- 31.Luo X, Jedlicka S, Jellison K. 2017. Pseudo-second-order calcium-mediated Cryptosporidium parvum oocyst attachment to environmental biofilms. Appl Environ Microbiol 83:e02339-16. doi: 10.1128/AEM.02339-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Sarkhosh T, Zhang XF, Jellison KL, Jedlicka SS. 2019. Calcium-mediated biophysical binding of Cryptosporidium parvum oocysts to surfaces is sensitive to oocyst age. Appl Environ Microbiol 85:e00816-19. doi: 10.1128/AEM.00816-19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Luo X, Jedlicka SS, Jellison KL. 2017. Role of wall shear stress in Cryptosporidium parvum oocyst attachment to environmental biofilms. Appl Environ Microbiol 83:e01533-17. doi: 10.1128/AEM.01533-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Hijjawi NS, Meloni BP, Ng'anzo M, Ryan UM, Olson ME, Cox PT, Monis PT, Thompson RCA. 2004. Complete development of Cryptosporidium parvum in host cell-free culture. Int J Parasitol 34:769–777. doi: 10.1016/j.ijpara.2004.04.001. [DOI] [PubMed] [Google Scholar]
- 35.Koh W, Clode PL, Monis P, Thompson R. 2013. Multiplication of the waterborne pathogen Cryptosporidium parvum in an aquatic biofilm system. Parasit Vectors 6:270. doi: 10.1186/1756-3305-6-270. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Koh W, Thompson A, Edwards H, Monis P, Clode PL. 2014. Extracellular excystation and development of Cryptosporidium: tracing the fate of oocysts within Pseudomonas aquatic biofilm systems. BMC Microbiol 14:281. doi: 10.1186/s12866-014-0281-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Adeyemo FE, Singh G, Reddy P, Bux F, Stenström TA. 2019. Efficiency of chlorine and UV in the inactivation of Cryptosporidium and Giardia in wastewater. PLoS One 14:e0216040. doi: 10.1371/journal.pone.0216040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Bones AJ, Jossé L, More C, Miller CN, Michaelis M, Tsaousis AD. 2019. Past and future trends of Cryptosporidium in vitro research. Exp Parasitol 196:28–37. doi: 10.1016/j.exppara.2018.12.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Morita S, Namikoshi A, Hirata T, Oguma K, Katayama H, Ohgaki S, Motoyama N, Fujiwara M. 2002. Efficacy of UV irradiation in inactivating Cryptosporidium parvum oocysts. Appl Environ Microbiol 68:5387–5393. doi: 10.1128/aem.68.11.5387-5393.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Soliman A, El-Adawy A, Abd El-Aal AA, Elmallawany MA, Nahnoush RK, Eiaghni ARA, Negm MS, Mohsen A. 2018. Usefulness of sunlight and artificial UV radiation versus chlorine for the inactivation of Cryptosporidium oocysts: an in vivo animal study. Open Access Maced J Med Sci 6:975–981. doi: 10.3889/oamjms.2018.180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Wolyniak DiCesare EA, Hargreaves BR, Jellison KL. 2012. Biofilms reduce solar disinfection of Cryptosporidium parvum oocysts. Appl Environ Microbiol 78:4522–4525. doi: 10.1128/AEM.08019-11. [DOI] [PMC free article] [PubMed] [Google Scholar]