
FIGURE 1. A model indicating roles for MLH proteins in meiotic recombination.
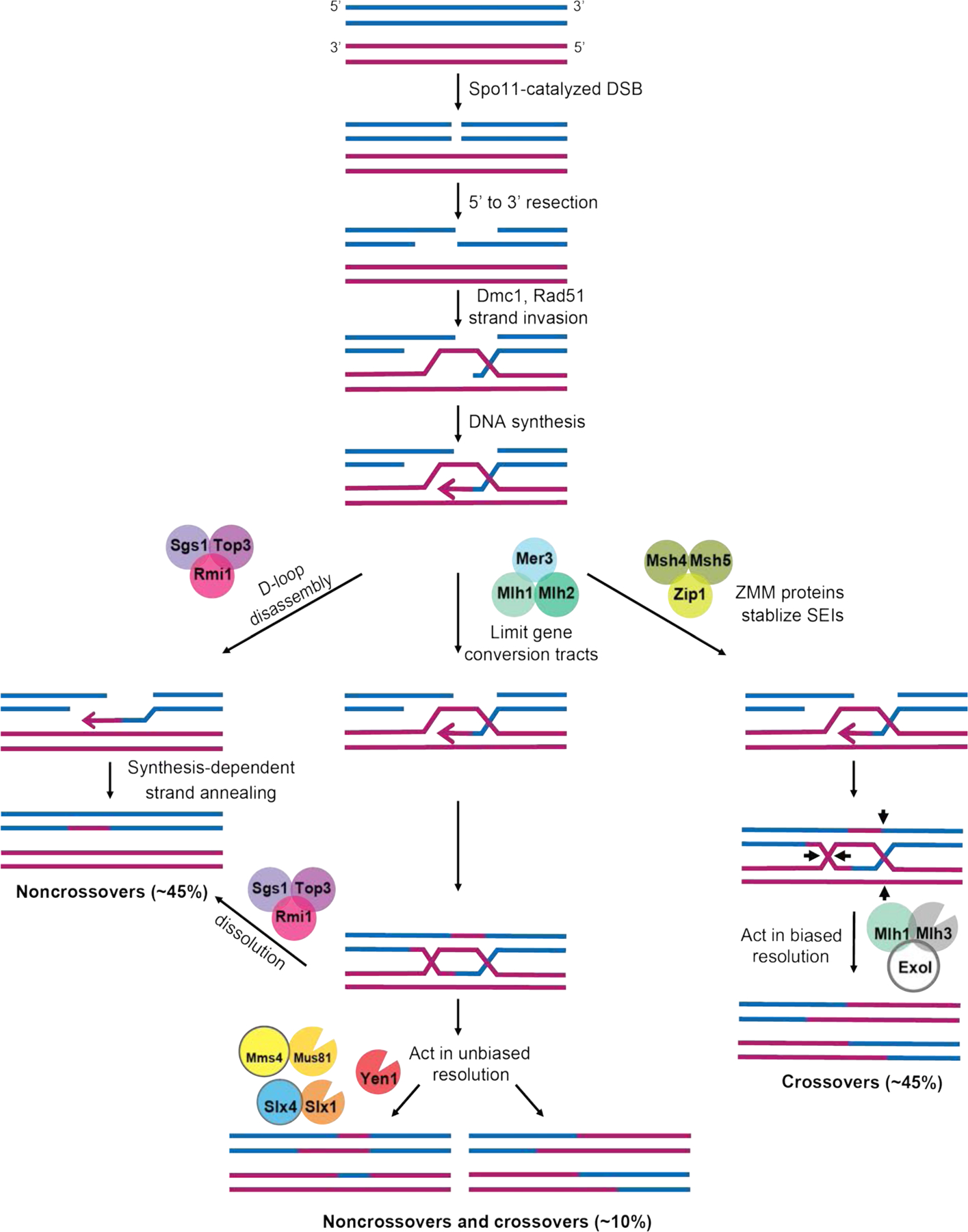
In diploid yeast induced to undergo meiosis, Spo11 catalyzes double-strand breaks (DSBs) throughout the genome, which undergo 5’ to 3’ resection. The resulting 3’ single stranded tail can invade the homologous chromosome to form a D-loop intermediate. In the ZMM-stabilized (Class I) pathway, the single end invasion intermediate (SEI) is stabilized by the actions of Msh4-Msh5 and Zip3 to promote D-loop extension through DNA repair synthesis. A double Holliday junction (dHJ) is then formed by second end capture that is asymmetrically cleaved in an Mlh1-Mlh3-dependent step to yield primarily crossover products. Note that in this model branch migration of Holliday junctions can occur but is not shown. Noncrossovers and a minority of crossovers (class II) are thought to occur through the other pathways shown (reviewed in Manhart & Alani, 2016; Wild & Matos, 2016; Zakharyevich et al., 2012). Roles for the Mlh1-Mlh2 complex in limiting gene conversion tract length, and Mlh1-Mlh3 in biased resolution of double-Holliday junctions to form crossovers, are shown. Mlh1-Pms1 acts to repair DNA mismatches that form in heteroduplex DNA in all pathways. The distribution of the types of meiotic recombination events in baker’s yeast is approximate and was calculated based on studies showing that Spo11 catalyzed DSBs produce equal numbers of crossovers and noncrossovers (Mancera, Bourgon, Brozzi, Huber, & Steinmetz, 2008; Martini, Diaz, Hunter, & Keeney, 2006; Marsolier-Kergoat et al., 2018). Of the crossovers, 85% are estimated to be class I, and 15% class II (Cooper et al., 2018; Jessop & Lichten, 2008; Oh, Lao, Taylor, Smith, & Hunter, 2008). Of the noncrossovers, 15% are estimated to result from the resolution of dHJs in the class II pathway (DeMuyt et al., 2012), with the remainder occurring through other mechanisms such as synthesis-dependent strand annealing. See text for details and Marsolier-Kergoat et al. (2018) and Pyatnitskaya, Borde, & De Muyt (2019) for more detailed models.