
Figure 2. Meiotic prophase I and maturation are disrupted by female infertility-associated gene variants.
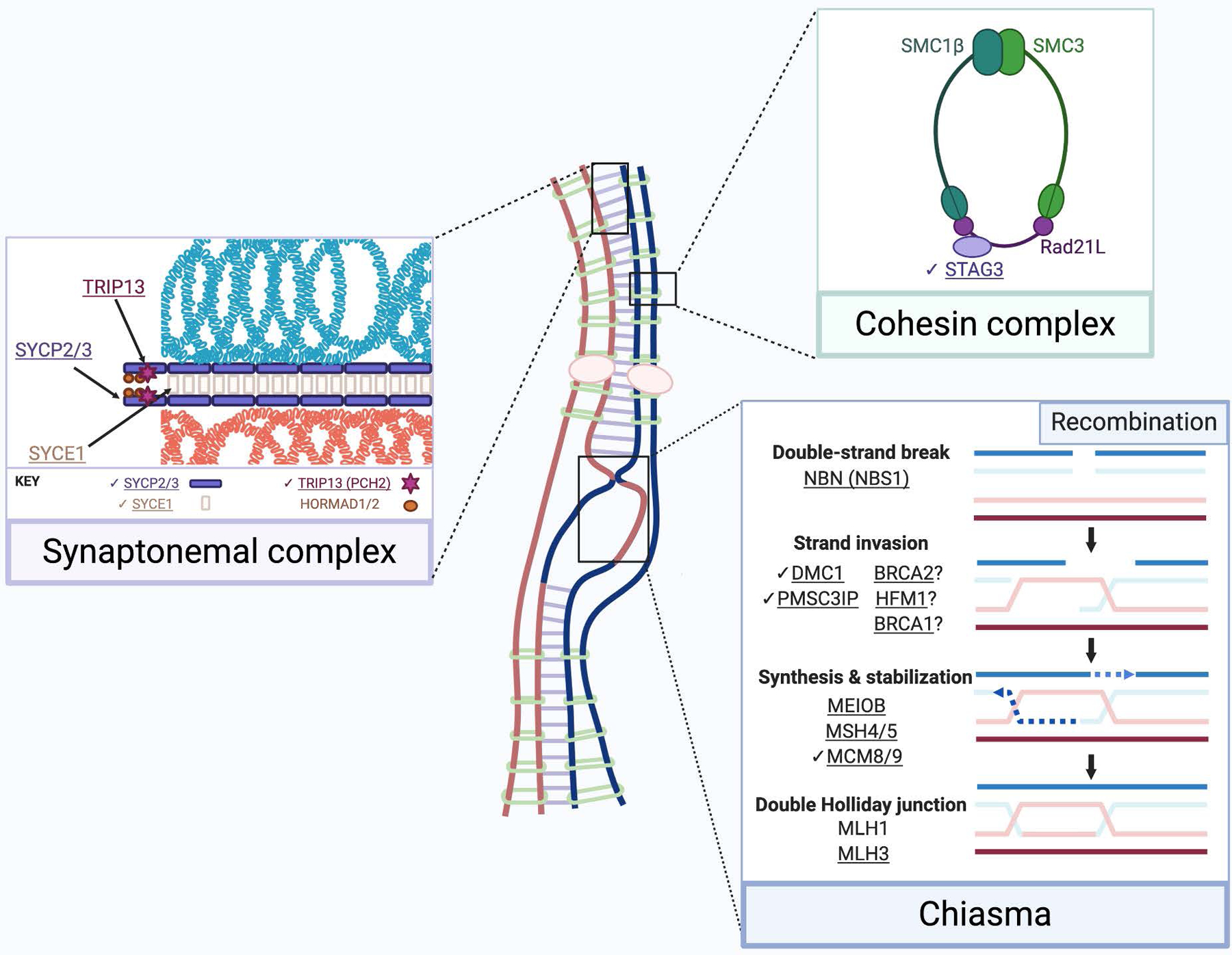
The synaptonemal complex (purple box) scaffolds homologous chromosomes and is composed of two lateral elements (SYCP2/3) and central elements SYCE1 (shown), SYCP1, SYCE2, SYCE3, and TEX12 (not shown). TRIP13 removes HORMA domain proteins (HORMADs), permitting synaptonemal complex assembly. The meiotic cohesin complex (green box) joins sister chromatids. In parallel with synaptonemal complex assembly, recombination (blue box) begins with double-strand breaks via SPO11. This requires the MRE11-RAD50-NBS1 (MRN) complex. Recombinases RAD51 (not shown) and DMC1 facilitate strand invasion into a chromosome’s homolog. HFM1 may also be essential for stable strand invasion (Guiraldelli et al., 2013). At double-strand breaks, the MCM8/9 helicase facilitates RAD51 recruitment and DNA synthesis. BRCA2 is proposed to localize to DSBs via MEILB2, and recruits RAD51 and DMC1 (Zhang et al., 2019a). A PMSC3IP-MND1 heterodimer stabilizes the RAD51 filament. BRCA1 may also stabilize the RAD51 filament in meiosis (Scully et al., 1997, Li et al., 2018). MEIOB is essential for maintenance of RAD51 and DMC1 foci (Souquet et al., 2013). The resulting double Holliday junction is stabilized by the MSH4/5 heterodimer. The MSH4/5 complex recruits the MLH1/MLH3 heterodimeric endonuclease, which processes most crossovers. Exact stoichiometry is not depicted. Underlined gene products are implicated in infertility. Check marks (✔) indicate gene products discussed in this review. Created using BioRender.com.