

Abstract
Insulin-like growth factor 1 (IGF-1) is a dichotomous hormone. While beneficial for growth/repair, and regulating muscle hypertrophy, high concentrations of IGF-1 are associated with increased risk of cancer and mortality. Factors thought to mediate IGF-1 include dietary protein and exercise. The purpose of this study was to analyze acute effects of dietary protein and/or exercise on plasma free IGF-1 and the time-course thereof to inform individuals who may benefit from increased IGF-1 (muscle growth/repair) or reduced IGF-1 (risk/diagnosis of cancer). Twenty-four participants (11 females, 24.9±4.6y) completed the three-way crossover study consisting of: (1)a high protein (42g) meal; (2)exercise (20min with four 30sec sprints); and (3)exercise followed by a high protein meal. Blood samples were collected fasted at rest, immediately after rest (or 5min after exercise), and at regular intervals throughout a 5h recovery. An additional fasted venipuncture was performed the morning following each condition (24h after baseline). Free IGF-1 was higher at immediately after exercise in the exercise condition (p=0.04). In the protein condition the 24h IGF-1 was 17.5% higher (p=0.02) than baseline. IGF-1 did not change over time in response to exercise with protein. The data gleaned from this study can enhance the knowledge of the time-course effects from protein and/or exercise on IGF-1. This study can provide a foundation for future research to investigate optimal timing and dosage to enhance muscle protein synthesis for athletes, as well as investigate whether consistent high protein meals may chronically elevate IGF-1 and increase the risk of deleterious health outcomes.
Keywords: insulin-like growth factor, somatotropin, physical activity, diet
INTRODUCTION
Insulin-like growth factor 1 (IGF-1) stimulates growth, repair, and is an important regulator of muscle hypertrophy (1). However, high concentration of IGF-1 is associated with increased risk of some types of cancer (2, 3) and mortality (4). Factors that are thought to mediate blood IGF-1 concentration include dietary protein (5, 6) and exercise (5, 6). Could it be possible to manipulate diet and exercise to produce acute increases in IGF-1 that maximize the benefits of exercise but stave off the deleterious effects of long term increases in IGF-1? In order to answer this question, we first need to better understand the acute response of IGF-1 to a single dose of dietary protein, exercise, and protein with exercise. In this way, future research can expand upon our findings and analyze the long-term effects of each mediating factor.
Previous research investigating the effects of dietary protein on IGF-1 in cross sectional studies has shown that chronic elevations in dietary protein are associated with increased IGF-1 concentration (7) and increased cancer risk (8, 9). However, no study has tracked the acute IGF-1 response to a single dose of dietary protein.
While numerous studies have examined the effect of exercise on IGF-1 concentration (10, 11), the results appear to be contingent upon exercise type. Resistance (12, 13) and sprint (14) exercises have been shown to elevate IGF-1 concentration and endurance exercise (15, 16) has been shown to decrease IGF-1 concentration.
Studies investigating the acute IGF-1 response to the combined effect of exercise and protein ingestion have predominantly used resistance exercise (17, 18) or endurance running (19) and have failed to show significant changes in IGF-1 concentration, indicating a possible interaction effect between protein and exercise. Thus, this study seeks to determine to identify possible interactions between dietary protein and sprint exercise, the condition which is most applicable to athletes that routinely consume protein after exercise to increase muscle anabolic properties. Furthermore, there are no studies that have investigated the effects of all three conditions (protein, exercise and protein with exercise) in the same individuals.
The present study was designed to better understand the impact a single dose of protein and exercise, can have on IGF-1 and, specifically, if exercise modulates any protein-induced increase in IGF-1 concentration. Therefore, the purpose of this study was to analyze acute effects of dietary protein and exercise, independently and combined, on free IGF-1 concentration and the time-course thereof.
METHODS
Participants.
Participants were recruited locally via flyers, social media postings and Otago University email lists. All participants (males = 13, females = 11) were healthy, not on medication (other than birth control pills or vitamins/minerals), not diabetic nor in any identifiable disease state, BMI<30 not pregnant or lactating nor planning pregnancy, and were physically capable of participating in physical activity (Table 1).
Table No. 1:
Participant characteristics (n=24; m=13, f=11)
| Age (year) | 24.9 ± 4.6 |
| Height (cm) | 173.9 ± 10.6 |
| Body mass (kg) | 71.9 ± 12.6 |
| Body fat (%) | 18.5 ± 8.4 |
| VO2max (ml/kg/min) | 47.0 ± 9.9 |
Values are means ± SD
Experimental Design
This randomized, three-way crossover study was performed in Dunedin, New Zealand between January and March 2019. Trials were conducted at the same time of day for each participant, separated by a minimum of one week, and maximum of 17 days (mdn = 7 days). The three conditions consisted of: (1) a high protein (42 g) meal; (2) exercise (20 min with four 30 second sprints); and (3) exercise followed by a high protein meal. Participants were randomly assigned to complete the three conditions in one of six possible orders via a random number generator (Microsoft Excel for Office 365 MSO 16.0). The trials were not blinded; however, participants were not told the condition in which they participated until they reported to the facility the morning of testing and the subsequent analysis of the samples was blinded. The study was approved by the University of Otago Ethics Committee (H18/133), conformed to the standards set by the Declaration of Helsinki, and written informed consent was obtained from all participants. The study was registered with the Australian New Zealand Clinical Trials Registry (ACTRN12618001919202p).
Pre-condition Testing
After providing written informed consent, and at least one week prior to participating in the first condition, anthropometric measurements, bio-electrical impedance analysis (BIA; InBody230 GBC BioMed, Auckland, N.Z.) for body composition, and a maximal oxygen capacity (VO2max) test were conducted. Participants completed the Physical Activity Readiness Questionnaire (PAR-Q) to assess suitability to perform maximal exercise, and an exercise history questionnaire to determine the amount of exercise they have been performing in the three months preceding the experiment. No one was excluded based on the results of the PAR-Q. The VO2max test was performed on a cycle ergometer (Velotron, Seattle WA, USA) via a modified Andersen protocol (20) and was used to set the workload for the exercise condition of the study as well as to detect the training status of the participants (20).
Experimental Conditions
Protein Condition
Participants reported to the testing facility in the morning after a 10 h overnight fast. Participants confirmed that they did not perform exercise for 24 h prior to testing and body composition was assessed using BIA. Thirty min after the baseline assessment point (the first blood sample) (Figure 1), participants were immediately handed a high protein liquid meal (whey protein isolate and dextrose mixed in almond milk) to consume within 10 min. The high protein meal provided 45 g of protein, 28 g of carbohydrate, and 12 g of fat, totaling 1707 kJ. After consuming the high protein meal participants rested quietly for the remainder of the session, consuming only water. At the completion of the session, participants were provided with a meal consisting of mini pizzas (or allergen-free equivalent) (1410 kJ, 14.2 g protein, 7.8 g fat, and 50.9 g carbohydrates per pizza), which they consumed ad libitum at the competition of the first condition. The number of pizzas consumed was then replicated after the participants’ second and third sessions. Participants left the facility and were instructed to record their diet from the time they left the facility until 10 h before their final blood draw of the condition (24 h after the baseline blood draw). They were told to replicate this diet after each condition (including a fast for 10 h before their final blood draw of each condition). In addition, participants were asked to refrain from physical activity until after they had returned to the facility the next morning.
Figure 1: Protocol.
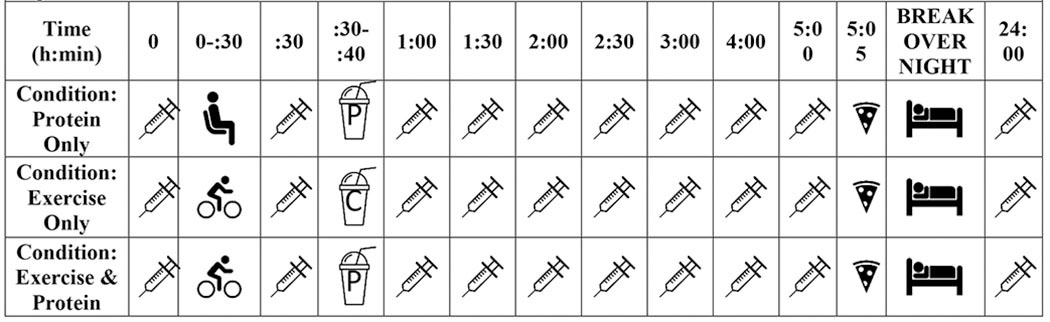
Timeline of protocol. P= protein meal, C= isocaloric placebo meal,  = blood draw,
= blood draw,  = seated and quiet rest,
= seated and quiet rest,  = cycling exercise protocol,
= cycling exercise protocol,  = overnight rest,
= overnight rest,  = pizza
= pizza
Exercise Condition
The protocol was the same as in the protein condition, however, instead of resting quietly prior to consuming the meal, participants performed a 20-minute exercise session on a cycle ergometer consisting of four cycles of: 4 min at 15% of aerobic threshold power followed by 30 seconds of maximum effort at 100% of aerobic threshold power. Upon completion of the exercise, participants received an isocaloric low-protein liquid meal, which they were instructed to consume within 10 min. The low-protein meal consisted of: dextrose and cocoa powder mixed in almond milk (1657 kJ, 65 g carbohydrate, 5.9 g protein, 11.6 g fat).
Protein and Exercise Condition
The protocol was the same as in the exercise condition. However, exercise was followed by consumption of the high protein liquid meal instead of the low-protein meal (compositions as noted in prior sections).
Blood Collection
A venous cannula (BD Adsyte Pro 20 GA 61 mL/min, Sandy, Ut) was inserted into the antecubital vein approximately 10 min after the participant arrived for testing. Blood samples were collected at rest, immediately after rest or within five min of concluding exercise (depending on the condition), and at regular intervals during the recovery period (Figure 1). Samples were collected from the cannula into syringes and immediately transferred to EDTA vacutainer tubes (K2E 10.8mg, Plymouth, UK), inverted 8 times, refrigerated, and within an hour of collection, centrifuged at 2000 g for 10 min (Digisystem Laboratory Instruments Inc., Table Top Centrifuge, New Taipei City, TW). Plasma was aliquoted into two microcentrifuge tubes (1.5 mL) and frozen at −80°C until later analysis. The cannula was removed after the five h of testing. The fasted 24 h blood samples were collected into EDTA vacutainer tubes (K2E 10.8mg, Plymouth, UK), which were treated and processed in the same manner as above.
Analytical Methods
Free IGF-1 concentration was measured using commercially available enzyme-linked immunosorbent assays (ab211651 Human IGF1 SimpleStep ELISA Kit, Cambridge, UK).
Absorbance was read on a microplate reader at 450 nm (BioRad Model680, Hercules, CA). All analyses were performed in duplicate, and all analyses for each participant were performed within the same assay batch to reduce inter-assay variance in compliance with recommended laboratory procedures (21). The intra-assay variance was 15.2%.
Statistical Analysis
Data are expressed as mean ± SD. Data were analyzed using STATA statistical software (STATA IC 15, College Station, TX, USA). A P<0.05 was considered statistically significant.
A sample size of 24 participants was chosen to provide 80% power to detect a treatment effect of a 0.5 standard deviation difference in mean IGF-1 area under the curve (AUC) response, assuming a within participant correlation of 0.7 and an alpha of 0.05. Total AUC was calculated by trapezoidal reconstruction for the time period between baseline and 300 min (i.e. total duration of a session). Differences in area under the curve between conditions was calculated by a within subject repeated measures analysis of variance (ANOVA). A within subject repeated measures ANOVA was also used to detect between condition difference in: baseline IGF-1 concentration, incremental AUC, peak IGF-1 concentration, time of peak concentration, IGF-1 concentration at 24 h, and change in IGF-1 concentration from baseline to peak.
A multilevel mixed-effects linear regression was run to investigate the change in IGF-1 concentration over time for all three conditions. Tests of simple main effects were run to determine which time points were significantly different from baseline in each condition.
A separate multilevel mixed-effects linear regression was utilized to investigate potential differences between the baseline measure and 24 h after baseline in each condition. Tests of simple main effects were conducted to determine if IGF-1 concentration at 24 h after baseline was significantly different from baseline in each condition.
RESULTS
Twenty-four recreationally active males and females completed the study. The characteristics of these participants are presented in Table 1. Participants that could not complete the study were excluded from the analysis. Reasons for the dropouts included: scheduling conflicts due to the time commitment necessary to complete the study (four), fainting when the cannula was inserted into their arm or being uncomfortable with blood draws (five), inability to find a suitable vein for cannulation (one), or family emergency (one).
Changes in IGF-1 Concentration over Time
In the protein condition, IGF-1 concentration did not change over the first 300 min (p=0.56). However, at 24 h IGF-1 concentration was 3.41 pg/mL higher (95% CI 0.64 to 6.07; p=0.02) than at baseline (Figure 2).
Figure 2:

Mean free IGF-1 concentration (pg/mL) from fasted, morning baseline through 300 min after baseline and next morning fasted IGF-1 concentration (1440 min). * significant change from baseline free IGF-1 concentration
In the exercise condition IGF-1 concentration was significantly higher at 30 min (immediately after exercise) (Δ 3.14 pg/mL; 95% CI 0.39 to 5.90; p=0.04) (Figure 2). At none of the other timepoints was IGF-1 different from baseline (including at 24 h (Δ 2.65 pg/mL; 95% CI 2.62 to 2.68; p=0.69)).
In the exercise and protein condition IGF-1 concentration did not change over time (first 300 min p=0.712) or from baseline to 24 h (Δ 1.63 pg/mL; 95% CI 1.60 to 1.66; p=0.391).
Differences between Conditions
The plasma IGF-1 AUC was not significantly different between conditions (p=0.69) (Figure 2). The baseline IGF-1 concentration (p=0.93), peak concentration (p=0.88), change from baseline to peak concentration (p=0.37), and change from baseline to 24 h concentration (p=0.63) were not statistically significant between conditions. The difference in IGF-1 concentration at 24 h between conditions approached significance (p=0.07). The time to peak concentration within the first 300 min was significantly longer in the protein condition than the exercise condition (85.00 ± 34.64 min; p=0.01) and the protein with exercise condition (66.41 ± 35.63 min; p=0.04). The time to peak concentration was similar in the protein with exercise and exercise only conditions (p=0.51).
DISCUSSION
To our knowledge, our study is the first to show that IGF-1 concentration is increased 24 h after ingesting protein (45 g). We also observed an increase in IGF-1 concentration immediately following high-intensity interval cycling, which is similar to that observed with previous research, indicating such exercise produces elevations in IGF-1 concentration immediately after exercise (22, 23). Interestingly, when exercise preceded protein ingestion no increase in IGF-1 concentration at 24 h was observed.
Differences Between Conditions and Potential Mechanisms:
When analyzing the differences between conditions for summary measures, the AUC was similar in all three conditions. However, the shapes of the IGF-1 concentration response curves were different (Figure 2). Therefore, it is possible that the different stimuli (dietary protein vs exercise vs protein with exercise) elicit different release mechanisms or a change in the sensitivity.
The mechanisms through which dietary protein affects free IGF-1 concentration have yet to be fully elucidated. However, studies investigating the effects of consecutive days of high protein diets have shown IGF-1 to increase linearly with protein dose (24). Interestingly, our results indicate no significant changes in IGF-1 concentration for up to 5 hours after ingestion, but a significant, increase at 24 h. It is possible that the hepatic release of IGF-1 is a slow process and any rise in IGF-1 caused by dietary protein may not be immediately observed in the circulation. The mechanisms underlying the release of IGF-1 after exercise are not fully understood either. However, there are three leading theories(25) : 1) exercise increases growth hormone secretion, which results in increased hepatic production of IGF-1 and circulating IGF-1(26); 2) growth hormone secretion stimulated by exercise induces endogenous production of IGF-1 by the muscle; 3) exercise stimulates the active muscle to produce IGF-1 independently of circulating growth hormone(27). Thus, differences in the time course response of IGF-1 concentration between our experimental conditions may be due to independent regulatory mechanisms associated with protein consumption and exercise. However, the mechanisms of IGF-1 release during this investigation was not a part of our intent.
Effects of Dietary Protein:
The most novel finding of our study is that IGF-1 concentration was significantly greater than baseline at the 24 h measure after ingesting protein. This increase has not been observed previously and may be indicative of a delayed effect caused by the consumption of protein. Since the half-life of IGF-1 is six h (28) and the stability of IGF-1 concentration can increase due to binding proteins (28), it is possible that dietary protein consumption causes changes in circulating IGF-1 by creating a complex with binding protein and extending the half-life to 15 h(28). Regrettably, we were unable to assess IGF binding proteins. The change in IGF-1 concentration 24 h after protein ingestion may have implications for long term IGF-1 regulation. Since an elevation was seen with a single high protein meal, it is possible that multiple high protein meals on a consistent basis may chronically or repeatedly elevate IGF-1 concentration. Future research should address this supposition and determine the validity of our speculation.
It should be noted that we chose to monitor IGF-1 concentration for five h after baseline and at one time point 24 h after baseline. These time points were chosen in order to monitor the IGF-1 response immediately following protein ingestion, where we would have expected to see the most change. We used the 24 h measure to reduce any potential effects of circadian IGF-1 release and reveal how the effects of each condition altered the fasted, morning IGF-1 concentration. Thus, we feel any change observed in IGF-1 was due to the condition and not the timing of sampling.
Effects of Exercise:
The observed effects of exercise on IGF-1 concentration, are in agreement with others who conducted two thirty second sprints on a cycle ergometer with four min recovery between sprints (29), demonstrating that high-intensity exercise acutely elevates IGF-1 concentration (30). Part of this observed response may be due to plasma volume shifts. Plasma volume shifts occur during the onset of exercise due to osmotic movement of water from the intravascular space to the active muscle (31). We did not measure hematocrit (nor hemoglobin) and thus do not know the magnitude of the hemoconcentration. However, we did measure an increase in IGF-1 concentration immediately after exercise of 16 percent. Based upon the observed hemoconcentration with this type of exercise, ~2 percent as reported by others (29), we feel confident to conclude that the post-exercise rise in IGF-1 was in response to the exercise bout and not solely hemoconcentration. Moreover, as free IGF-1 should move with the plasma out of the vascular space, any effect of plasma volume shifts should not affect the observed concentration in reflecting IGF-1 release (32). Furthermore, the physiological influence of hormones on target tissue receptors is affected by concentration change, regardless of the mechanism by which that change is implemented (33).
Despite an acute transient increase in IGF-1, exercise appears not to result in any lasting changes in IGF-1, as the concentration measured at 24 h was not different from baseline. This is consistent with several other previous studies that also found no change the morning after a 36 km run(15), one hour of single-leg kicking (in men(34) and women in both the luteal and follicular phase(35)), moderate duration resistance exercise(36), long duration resistance exercise(36), moderate duration cycling(36), and long duration cycling(36). However, it is possible that when exercise induces a large energy deficit, decreases in IGF-1 concentration can occur since reductions in IGF-1 24 h after an ultra-endurance race have been observed in male participants (37). In addition, decreases in body weight/fat mass and decreases in IGF-1 have been observed as a result of restricted energy intake in military personnel during simulated field operations (38).
Effects of Protein with Exercise:
Previous studies have investigated the relationship between protein consumed immediately after exercise and IGF-1. However, most of these studies involved resistance exercise (17, 18) or endurance running (19, 39) and failed to find a significant effect of protein with exercise on IGF-1 concentration. Our study is the first to monitor the effects of dietary protein with intermittent high-intensity sprint cycling exercise. Our results indicate that the combination of dietary protein and exercise dose not result in any statistically significant change in IGF-1 concentration over time. This is a surprising result given that previous studies have shown ingestion of protein after exercise results in greater skeletal muscle anabolism when compared to exercise and a placebo beverage(40, 41). Thus, we would have expected exercise and dietary protein to have an additive effect on IGF-1 concentration. This was not the case; but it is possible that under different conditions it may have occurred. Since no such additive effect was observed with our protocol, further research is needed to determine the optimal conditions (timing, dose type of protein) for protein supplementation after exercise to enhance IGF-1 and protein synthesis.
When protein ingestion was preceded by exercise there was no effect on IGF-1 the following morning. The effect of exercise on protein induced IGF-1 release may be indicative of a protective mechanism produced by exercise in which IGF-1 concentrations return to baseline faster. It is possible that an exercise induced increase in IGF-1 binding proteins reduced the quantity of free IGF-1 in circulation (42). However, without measuring the concentration of IGF-1 binding proteins we are unable to confirm this theory. To our knowledge, this is the first study with a subsequent 24 h, measure to investigate the effects of exercise with protein on IGF-1. An additive, or perhaps synergistic, effect when protein ingestion was preceded by exercise could have been expected. That is, specifically showing a significant increase after exercise (due to stimulator actions of the physical activity and plausible localized IGF-1 release) as well as at 24 h (due to the previously observed 24 h increase in the protein condition and plausible systemic IGF-1 release). Surprisingly, this was not observed. The total lack of response suggests a potential antagonism action exists from the interaction of protein ingestion and exercise (i.e., as manipulated in this study). We cannot explain why such an action would occur, but it could represent pre- or post-secretion regulatory events that control IGF-1 levels in the circulation and maintain homeostasis. Future research needs to pursue this interesting finding and elucidate the mechanism(s).
CONCLUSIONS:
Our results demonstrate that the timing and magnitude of plasma free IGF-1 concentration are influenced differently by exercise, than with protein ingestion, or when protein ingestion is preceded by exercise. Our results are the first to describe the time course response of free IGF-1 to dietary protein. While all three conditions resulted in a similar area under the curve for IGF-1 concentration, the timing and magnitude of change in IGF-1 concentration were different. Dietary protein did not produce significant changes in IGF-1 concentrations in the initial 5 h after ingestion, but it did significantly increase IGF-1 concentration 24 h after baseline. The exercise with protein condition, a single bout of intermittent sprint exercise followed by protein ingestion, most closely resembles the current practices of athletes seeking to maximize skeletal muscle growth by the consumption of protein directly after a workout. The 24 h increase in IGF-1 observed in the protein only condition was mitigated by performing exercise with protein. This finding may be important for the long-term health of athletes or individuals on a high protein diet. Since it appears that exercise prior to protein ingestion provides a protective mechanism it may be possible to mitigate the harmful effects of a high protein diet with exercise. The data gleaned from this study can enhance the knowledge of the time course of effects from protein and exercise on IGF-1 concentration. In addition, this study can provide a foundation for future research to investigate optimal timing and dosage combinations to enhance muscle protein synthesis for athletes, as well as investigate whether multiple high protein meals on a consistent basis may chronically elevate IGF-1 concentration and increase the risk of deleterious health outcomes.
Table No. 2:
Differences Between Trials
| Protein Condition | Exercise Condition | Protein & Exercise Condition | |
|---|---|---|---|
| Area Under the Curve (pg/mL/h) | 97.26 ± 42.12 | 99.15 ± 35.10 | 94.66 ± 38.56 |
| Peak IGF-1 Concentration (pg/mL) | 24.67 ± 8.37 | 24.91 ± 7.71 | 25.24 ± 8.36 |
| Time to Peak IGF-1 Concentration within first 300 min (min after baseline) | 182.50 ±102.37 | 97.50 ± 95.38 | 116.09 ± 94.57 |
| Baseline IGF-1 Concentration (pg/mL) | 19.80 ± 9.32 | 19.48 ± 8.08 | 19.18 ± 6.78 |
| 24 h IGF-1 Concentration (pg/mL) | 23.27 ± 8.76 | 21.95 ± 8.27 | 20.07 ± 9.68 |
| Change from Baseline to 24 h IGF-1 Concentration (pg/mL) | 3.41 ± 6.37* | 2.65 ± 7.99 | 1.63 ± 8.58 |
Values are means ± SD
Statistically different 24 h from baseline
ACKNOWLEDGMENTS
We would like to thank our participants. Thank you to the University of Otago Department of Microbiology for their blood analysis instruction.
Financial Support
This work was supported by the University of Otago School of Physical Education, Sport & Exercise Sciences (C.G., University of Otago Post Graduate Research Grant, Postgraduate Research Scholarship). Dr. Peddie’s position is supported by a Research Fellowship from the National Heart Foundation of New Zealand (Grant number 1754) with funding from the Southland Medical Foundation.
Footnotes
Conflict of Interest
The authors declare no conflicts of interest arising from the conclusions of this work.
Declaration of Interest Statement
We, the authors, declare this manuscript is original, has not been published before and is not currently being considered for publication elsewhere.
The manuscript and order of authors has been read/approved by all authors. There are no other personas who satisfied the authorship criteria.
There are no impediments to publication. Our institutions intellectual property regulations have been followed.
The involvement of human patients has met ethical approval (acknowledged in manuscript).
We understand the Corresponding Author is the sole contact for the Editorial process and is responsible for communicating with the other authors.
REFERENCES
- 1.Baserga R The IGF-I receptor in cancer research. Exp Cell Res. 1999;253:1–6. [DOI] [PubMed] [Google Scholar]
- 2.Renehan A, Zwahlen M, Minder C, O’Dwyer S, Shalet S, Egger M. Insulin-like growth factor (IGF)-I, IGF binding protein-3, and cancer risk: systematic review and meta-regression analysis. The Lancet. 2004;363(9418):1346–53. [DOI] [PubMed] [Google Scholar]
- 3.Yu H, Spitz M, Mistry J, Gu J, Hong W, Wu X. Plasma levels of insulin-like growth factor-I and lung cancer risk: a case-control analysis. J Natl Cancer Inst. 1999;91(2):151–6. [DOI] [PubMed] [Google Scholar]
- 4.Levine M, Suarez J, Brandhorst S, Balasubramanian P, Cheng C-W, Madia F, et al. Low Protein Intake is Associated with a Major Reduction in IGF-1, Cancer, and Overall Mortality in the 65 and Younger but Not Older Population. Cell Metab. 2014;19:407–17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Thissen JP, Ketelslegers JM, Underwood LE. Nutritional regulation of the insulin-like growth factors. Endocr Rev. 1994;15:80–101. [DOI] [PubMed] [Google Scholar]
- 6.Kraemer W, Volek J, Bush J, Putukian M, Sebastianelli W. Hormonal responses to consecutive days of heavy-resistance exercise with or without nutritional supplementation. J Appl Physiol. 1998;85(4). [DOI] [PubMed] [Google Scholar]
- 7.Crowe FL, Key TJ, Allen NE, Appleby PN, Roddam A, Overvad K, et al. The Association between Diet and Serum Concentrations of IGF-I, IGFBP-1, IGFBP-2, and IGFBP-3 in the European Prospective Investigation into Cancer and Nutrition. Cancer Epidemiol Biomarkers Prev. 2009;18(5):1333–40. [DOI] [PubMed] [Google Scholar]
- 8.Schernhammer E, Holly J, Pollak M, Hankinson S. Circulating levels of insulin-like growth factors, their binding proteins, and breast cancer risk. Cancer Epidemiol Biomarkers Prev. 2005;14:699–704. [DOI] [PubMed] [Google Scholar]
- 9.Hankinson S, Willett W, Colditz G, al. e. Circulating concentrations of insulin-like growth factor-I and risk of breast cancer. Lancet. 1998;351:1393–6. [DOI] [PubMed] [Google Scholar]
- 10.Isley WL, Underwood LE, Clemmons DR. Changes in Plasma Somatomedin-C in Response to Ingestion of Diets with Variable Protein and Energy Content. Journal of Parenteral and Enteral Nutrition. 1984;8:407–11. [DOI] [PubMed] [Google Scholar]
- 11.Arjmandi BH, Lucas EA, Khalil DA, Devareddy L, Smith BJ, McDonald J, et al. One Year Soy Protein Supplementation has Positive Effects on Bone Formation Markers but not Bone Density in Postmenopausal Women. Nutr J. 2005;4(8):1–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Karabulut M, Sherk VD, Bemben DA, Bemben MG. Inflammation marker, damage market and anabolic hormone responses to resistance training with vascular restriction in older males. Clin Physiol Funct Imaging. 2013;33(5):393–9. [DOI] [PubMed] [Google Scholar]
- 13.Tsai C-L, Wang C-H, Pan C-Y, Chen F-C, Huang T-H, Chou F-Y. Executive function and endocrinological responses to acute resistance exercise. Frontiers in Behavioral Nuroscience. 2014;8:1–12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Stokes K, Nevill M, Frystyk J, Lakomy H, Hall G. Human growth hormone responses to repeated bouts of sprint exercise with different recovery periods between bouts. J Appl Physiol. 2005;99:1254–61. [DOI] [PubMed] [Google Scholar]
- 15.Olesen JL, Heinemeier KM, Geimner C, Kjaer M, Flyvbjerg A, Langberg H. Exercise-dependent IGF-1, IGFBPs, and type I collagen changes in human peritendinous connective tissue determined by microdialysis. J Appl Physiol. 2007;102:214–20. [DOI] [PubMed] [Google Scholar]
- 16.Scofield DE, McClung HL, McClung JP, Kraemer WJ, Rarick KR, Pierce JR, et al. A novel, noninvasive transdermal fluid sampling methodology: IGF-I measurement following exercise. Am J Physiol Regul Integr Comp Physiol. 2011;300:R1326–R32. [DOI] [PubMed] [Google Scholar]
- 17.Cooke MB, La Bounty P, Buford T, Shelmadine B, Redd L, Hudson G, et al. Ingestion of 10 grams of whey protein prior to a single bout of resistance exercise does not augment Akt/mTOR pathway signaling compared to carbohydrate. J Int Soc Sports Nutr. 2011. ;8(18):1–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Paoli A, Pacelli QF, Neri M, Toniolo L, Cancellara P, Canato M, et al. Protein Supplementation Increases Postexercise Plasma Myostatin Concentration After 8 Weeks of Resistance Training in Young Physically Active Subjects. Journal of Medical Food. 2015; 18(1):137–43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Betts JA, Stokes KA, Toone RJ, Williams C. Growth-Hormone Responses to Consecutive Exercise Bouts With Ingestion of Carbohydrate Plus Protein. Int J Sport Nutr Exerc Metab. 2013;23:259–70. [PubMed] [Google Scholar]
- 20.Andersen LB. A maximal cycle exercise protocol to predict maximal oxygen uptake. Scandonavian Journal of Medicine and Science in Sports. 1995;5:143–6. [DOI] [PubMed] [Google Scholar]
- 21.Hackney AC, Viru A. Research Methodology: Endocrinologic Measurements in Exercise Science and Sports Medicine. J Athl Train. 2008;43(6):631–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Hejazi SM. Effects of high intensity interval training on plasma levels of growth hormone and insulin like growth factor-1 in healthy males. Biosci Biotech Res Comm. 2017(Special Issue No 1):199–202. [Google Scholar]
- 23.Kraemer W, Harman F, Vos N, Gordon S, Nindl B, Marx J, et al. Effects of Exercise and Alkalosis on Serum Insulin-Like Growth Factor 1 and IGF-Binding Protein-3. Can J Appl Physiol. 2000;25(2):127–38. [DOI] [PubMed] [Google Scholar]
- 24.Dardevet D, Manin M, Balage M, Sornet C, Grizard J. Influence of low- and high-protein diets on insulin and insulin-like growth factor-1 binding to skeletal muscle and liver in the growing rat. Br J Nutr. 1991;65(1):47–60. [DOI] [PubMed] [Google Scholar]
- 25.Borst SE, Dehoyos DV, Garzarella L, Vincent K, Pollock BH, Lowenthal DT, et al. Effects of resistance training on insulin-like growth factor-I and IGF binding proteins. Med Sci Sports Exerc. 2001;33:648–53. [DOI] [PubMed] [Google Scholar]
- 26.KE Y Growth hormone effects on metabolism, body composition, muscle mass, and strength. Exerc Sport Sci Rev 1994;22:285–312. [PubMed] [Google Scholar]
- 27.De Vol D, Rotwein P, Sadow L, Novakovsky J, Bechtel PJ. Activation of insulin-like growth factor gene expression during work induced muscle hypertrophy. The American Journal of Physiology. 1990;259:E89–E95. [DOI] [PubMed] [Google Scholar]
- 28.Mejri S, Behir F, Ben Rayana MC, Ben Hamida J, Ben Slama C. Effect of training on GH and IGF-1 responses to a submaximal exercise in football players. Eur J Appl Physiol. 2005;95:496–503. [DOI] [PubMed] [Google Scholar]
- 29.Stokes KA, Sykes D, Gilbert KL, Chen J-W, Frystyk J. Brief, high intensity exercise alters serum ghrelin and growth hormone concentrations but not IGF-I, IGF-II or IGF-I bioactivity. Growth Horm IGF Res. 2010;20:289–94. [DOI] [PubMed] [Google Scholar]
- 30.Kraemer W, Gordon S, Fleck S, Marchitelli L, Mello R, Dziados J, et al. Endogenous Anabolic Hormonal and Growth Factor Responses to Heavy Resistance Exercise in Males and Females. Int J Sports Med. 1991;12:228–35. [DOI] [PubMed] [Google Scholar]
- 31.Kawabata T, Suzuki T, Miyagawa T. Effect of Blood Volume on Plasma Volume Shift During Exercise. J Therm Biol. 2004;29(7–8):775–8. [Google Scholar]
- 32.Biotechnology in Growth Regulation. Heap RB, Prosser CG, Lamming GE, editors: Butterworth-Heinemann; 2013. 296 p. [Google Scholar]
- 33.Goodman MH. Endocrinology Concepts for Medical Students. Adv Physiol Educ. 2001;25(4):213–24. [DOI] [PubMed] [Google Scholar]
- 34.Miller BF, Olesen JL, Hansen M, Dossing S, Crameri RM, Welling RJ, et al. Coordinated collagen and muscle protein synthesis in human patella tendon and quadriceps muscle after exercise. J Physiol. 2005;567(3):1021–33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Benjamin F, Miller MH, Olesen Jens L., Flyvbjerg Allan, Schwarz Peter, Babraj John A., Smith Kenneth, Rennie Michael J., Kjaer Michael. No effect of menstrual cycle on myofibrillar and connective tissue protein synthesis in contracting skeletal muscle. Am J Physiol Endocrinol Metab. 2005;290:E163–E8. [DOI] [PubMed] [Google Scholar]
- 36.Nindl BC, Alemany JA, Tuckow AP, Kellogg MD, Sharp MA, Patton JF. Effects of Exercise Mode and Duration on 24-h IGF-I System Recovery Responses. Med Sci Sports Exerc. 2009;41(6):1261–70. [DOI] [PubMed] [Google Scholar]
- 37.Berg U, Enqvist JK, Mattsson CM, Carlsson-Skwirut C, Sundberg CJ, Ekblom B, et al. Lack of sex differences in the IGF-IGFBP response to ultra endurance exercise. Scand J Med Sci Sports. 2008;18:706–14. [DOI] [PubMed] [Google Scholar]
- 38.Nindl BC, Alemany JA, Kellogg MD, Rood J, Allison SA, Young AJ, et al. Utility of circulating IGF-I as a biomarker for assessing body composition changes in men during periods of physical activity, energy and sleep restriction. J Appl Physiol. 2007;103:340–6. [DOI] [PubMed] [Google Scholar]
- 39.Bourrilhon C, Lepers R, Philippe M, Van Beers P, Chennaoui M, Drogou C, et al. Influence of Protein- Versus Carbohydrate-enriched Feedings on Physiological Responses During an Ultraendurance Climbing Race. Horm Metab Res. 2010;42:31–7. [DOI] [PubMed] [Google Scholar]
- 40.Tipton K, Elliott T, Cree M, Wolf S, Sanford A, Wolfe R. Ingestion of casein and whey proteins result in muscle anabolism after resistance exercise. Med Sci Sport Exer. 2004;36(12):2073–81. [DOI] [PubMed] [Google Scholar]
- 41.Josse A, Tang J, Tarnopolsky M, Phillips S. Body composition and strength changes in women with milk and resistance exercise. Med Sci Sport Exer. 2010;42(6):1122–30. [DOI] [PubMed] [Google Scholar]
- 42.Chicharro JL, Lopez-Calderon A, Hoyos J, Martin-Velasco AI, Villa G, Villanua MA, et al. Effects of an Endurance Cycling Competition on Resting Serum Insulin-like Growth Factor I (IGF-I) and its Binding Proteins IGFBP-1 and IGFBP-3. Br J Sports Med. 2001;35(5):303–7. [DOI] [PMC free article] [PubMed] [Google Scholar]