
Figure 3. Pathogenic DNA, RNA, and protein correction from a single in vivo ABE-AAV9 injection of a mouse model of human progeria.
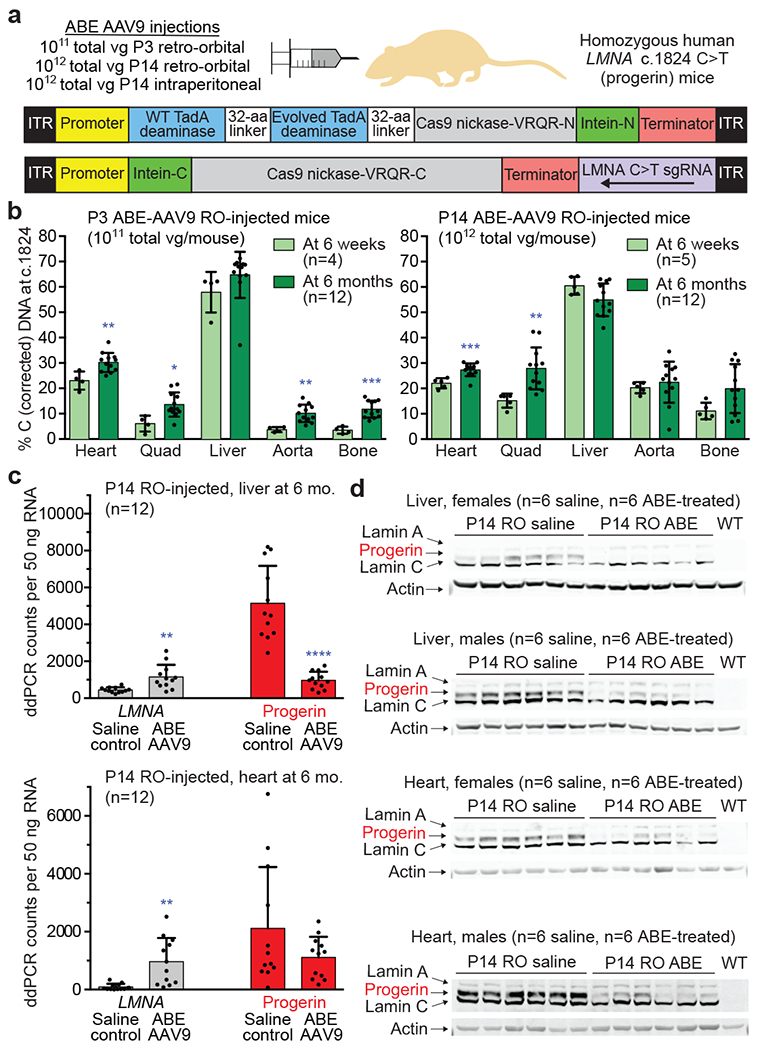
(a) Dual AAV9 encoding split-intein ABE7.10max-VRQR base editor halves26 and the LMNA-targeting sgRNA were injected into progeria mice. P3 retro-orbital (RO) injections (5×1010 of each AAV vg, 1×1011 vg total), P14 RO injections (5×1011 vg of each AAV vg, 1×1012 vg total), or P14 intraperitoneal (IP) injections (5×1011 vg of each AAV, 1×1012 vg total) were administered. (b) DNA editing efficiencies correcting LMNA c.1824 from T (pathogenic) to C (wild-type) for P3 RO-injected mice (left) or P14 RO-injected mice (right) in 6-week- or 6-month-old mice. Editing in P14 IP-injected mice is in Extended Data Fig. 3a. (c) ddPCR counts for human LMNA (grey bars) and progerin (red bars) RNA transcript abundance in P14 RO saline- or ABE-AAV9-injected mice in liver and heart. See Extended Data Fig. 4 for additional data. (d) Western blot analysis of human lamin A, progerin, and lamin C proteins in liver and heart of P14 RO saline- or ABE-AAV9-injected mice. Each lane shows tissue from a different mouse. WT is a C57BL/6 mouse lacking the transgene, showing that the antibody is specific to human lamin proteins. See Extended Data Fig. 5 for additional data. Values and error bars represent mean±SD for the indicated number of biological replicates. *p<0.05, **p<0.01, ***p<0.001, ****p<0.0001 by Student’s unpaired two-sided t-test.