
Figure 1.
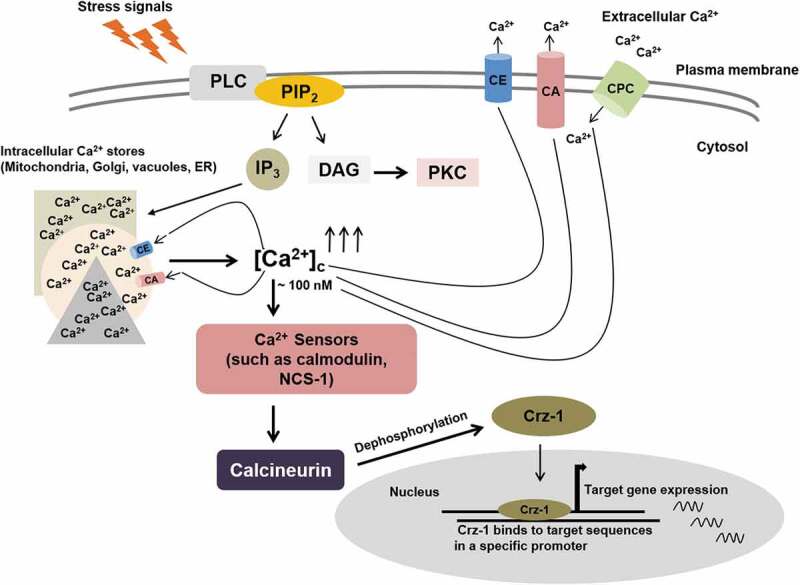
Overview of calcium signalling machinery in fungi. The membrane-bound phosphoinositide-specific phospholipase C (PLC) hydrolyzes phosphatidylinositol-4, 5-bisphosphate (PIP2) to produce two important second messenger molecules inositol 1,4,5-triphosphates (IP3) and diacylglycerol (DAG). IP3 induces Ca2+ release from intracellular stores, including mitochondria, Golgi, vacuoles, and endoplasmic reticulum (ER). DAG activates protein kinase C (PKC) that is involved in various signalling processes (Clapham 2007). PLCs also response to different stress signals (Barman et al. 2018). The resting intracellular free Ca2+ concentration ([Ca2+]c) is about 100 nM, an increase (shown using arrows pointing upward) of the [Ca2+]c is detected by various Ca2+ sensing proteins such as calmodulin (CaM) and NCS-1, which activate specific downstream signalling cascade. Calcineurin-Crz-1 pathway is shown here as an instance of a downstream signalling cascade. Ca2+ and CaM activate the serine/threonine phosphatase calcineurin that dephosphorylates the transcription factor Crz-1 for its nuclear location and subsequent expression of target genes in responses to stimuli. Excess Ca2+ is removed from the cytosol by Ca2+ exchangers (CE) and Ca2+ -ATPases (CA) proteins, whereas Ca2+ permeable channels (CPC) are required for the influx of extracellular Ca2+; and these processes are required to maintain Ca2+ homoeostasis in the cell (Tamuli et al. 2013)