

Abstract
Severe Acute Respiratory Syndrome Coronavirus-2 (SARS-CoV-2) transmission is uncontrolled in many parts of the world, compounded in some areas by higher transmission potential of the B1.1.7 variant now seen in 50 countries. It is unclear whether responses to SARS-CoV-2 vaccines based on the prototypic strain will be impacted by mutations found in B.1.1.7. Here we assessed immune responses following vaccination with mRNA-based vaccine BNT162b2. We measured neutralising antibody responses following a single immunization using pseudoviruses expressing the wild-type Spike protein or the 8 amino acid mutations found in the B.1.1.7 spike protein. The vaccine sera exhibited a broad range of neutralising titres against the wild-type pseudoviruses that were modestly reduced against B.1.1.7 variant. This reduction was also evident in sera from some convalescent patients. Decreased B.1.1.7 neutralisation was also observed with monoclonal antibodies targeting the N-terminal domain (9 out of 10), the Receptor Binding Motif (RBM) (5 out of 31), but not in neutralising mAbs binding outside the RBM. Introduction of the E484K mutation in a B.1.1.7 background to reflect newly emerging viruses in the UK led to a more substantial loss of neutralising activity by vaccine-elicited antibodies and mAbs (19 out of 31) over that conferred by the B.1.1.7 mutations alone. E484K emergence on a B.1.1.7 background represents a threat to the vaccine BNT162b.
Keywords: SARS-CoV-2, COVID-19, antibody, vaccine, neutralising antibodies, mutation, variant
Introduction
The outbreak of a pneumonia of unknown cause in Wuhan, China in December 2019, culminated in a global pandemic due to a novel viral pathogen, now known to be SARS-CoV-21. The unprecedented scientific response to this global challenge has led to the rapid development of vaccines aimed at preventing SARS-COV-2 infection and transmission. Continued viral evolution led to the emergence and selection of SARS-CoV-2 variants with enhanced infectivity/transmissibility2,3 4,5 and ability to circumvent drug6 and immune control7,8.
SARS-CoV-2 vaccines have recently been licensed that target the spike (S) protein, either using mRNA or adenovirus vector technology with protection rates ranging from 62 to 95%9–11. The BNT162b2 vaccine encodes the full-length trimerised S protein of SARS CoV-2 and is formulated in lipid nanoparticles for delivery to cells12. Other vaccines include the Moderna mRNA-1273 vaccine, which is also a lipid nanoparticle formulated S glycoprotein13 and the Oxford-AstraZeneca ChAdOx1 nCoV-19 vaccine (AZD1222) which is a replication-deficient chimpanzee adenoviral vector ChAdOx1, containing the S glycoprotein14. The duration of immunity conferred by these vaccines is as yet unknown. These vaccines were designed against the Wuhan-1 isolate discovered in 2019. Concerns have been raised as to whether these vaccines will be effective against newly emergent SARS-CoV-2 variants, such as B.1.1.7 (N501Y.V1), B.1.351 (N501Y.V2) and P1 (N501Y.V3) that originated in the UK, South Africa, and Brazil and are now being detected all over the world15–17.
In clinical studies of the Pfizer-BioNTech BNT162b2 vaccine, high levels of protection against infection and severe disease were observed after the second dose10. Neutralisating geometric mean titre (GMT) was below cut-off in most cases after prime dose, but as anticipated, titres substantially increased after boost immunization18. In older adults mean GMT was only 12 in a preliminary analysis of 12 participants19 and increased to 109 after the second dose.
In this study, we assess antibody responses against the the B.1.1.7 variant after vaccination with the first and second doses of BNT162b2, showing modest reduction in neutralisation against pseudoviruses bearing B.1.1.7 Spike mutations (ΔH69/V70, Δ144, N501Y, A570D, P681H, T716I, S982A and D1118H). In addition, by using a panel of human neutralising monoclonal antibodies (mAbs) we show that the B.1.1.7 variant can escape neutralisation mediated by most NTD-specific antibodies tested and by a fraction of RBM-specific antibodies. Finally, we show that the recent emergence and transmission of B.1.1.7 viruses bearing the Spike E484K mutation results in significant additional loss of neutralisation by BNT162b2 mRNA-elicited antibodies, convalescent sera and mAbs.
Results
Thirty seven participants had received the first dose of BNT162b2 mRNA vaccine three weeks prior to blood draw for serum and peripheral blood monocnulear cells (PBMC) collection. Median age was 63.5 years (IQR 47–84) and 33% were female. Serum IgG titres to Nucleocapsid (N) protein, S and the S receptor binding domain (RBD) were assayed by particle based flow cytometry on a Luminex analyser (Extended Data Fig. 1a). These data showed S and RBD antibody titres much higher than in healthy controls, but lower than in individuals recovered from COVID-19 and titres observed in therapeutic convalescent plasma. The raised N titres relative to control could be the result of non-specific cross reactivity that is increased following vaccination. However, the antibody response was heterogeneous with almost 100-fold variation in IgG titres to S and RBD across the vaccinated participants.
Using lentiviral pseudotyping we studied WT (wild type bearing D614G) and mutant B.1.1.7 S proteins (Fig. 1a) on the surface of enveloped virions in order to measure neutralisation activity of vaccine-elicited sera. This system has been shown to give results correlating with replication competent authentic virus20,21. Eight out of 37 participants exhibited no appreciable neutralisation against the WT pseudotyped virus following the first dose of vaccines. The vaccine sera exhibited a range of inhibitory dilutions giving 50% neutralisation (ID50) (Fig. 1c–d). The GMT against wild type (WT) following the second dose of vaccine was an order of magnitude higher than after the first dose (318 vs 77) (Fig 1c–f). There was correlation between full length S IgG titres and serum neutralisation titres (Extended Data Fig. 1b). A broad range of T cell responses was measured by IFN gamma FluoroSpot against SARS-CoV-2 peptides in vaccinees. These cellular responses did not correlate with IgG S antibody titres (Extended Data Fig. 1c–d).
Figure 1. Neutralization by first and second dose mRNA vaccine sera against wild type and B.1.1.7 Spike mutant SARS-CoV-2 pseudotyped viruses.

a, Spike in open conformation with a single erect RBD (PDB: 6ZGG) in trimer axis vertical view with the locations of mutated residues highlighted in red spheres and labelled on the monomer with erect RBD. Vaccine first dose (b-c, n=37), second dose (d-e, n=21) and convalescent sera, Conv. (f-g,n=27) against WT and B.1.1.7 Spike mutant with N501Y, A570D, ΔH69/V70, Δ144/145, P681H, T716I, S982A and D1118H. GMT with s.d presented of two independent experiments each with two technical repeats. Wilcoxon matched-pairs signed rank test p-values * <0.05, ** <0.01, ***<0.001, **** <0.0001, ns not significant HS – human AB serum control. Limit of detection for 50% neutralization set at 10.
We then generated mutated pseudoviruses carrying S protein with mutations N501Y, A570D and the H69/V70 deletion. We observed no reduction in the ability of sera from vaccinees to inhibit either WT or mutant virus (Extended Data Fig. 2a, b). A panel of sera from ten recovered individuals also neutralised both wild type and the mutated viruses similarly (Extended Data Fig. 2c). We next completed the full set of eight mutations in the S protein present in B.1.1.7 variant (Fig. 1a), ΔH69/V70, Δ144, N501Y and A570D in the S1 subunit and P681H, T716I, S982A and D1118H in the S2 subunit. All constructs also contained D614G. We found that among 29 individuals with neutralisation activity against the WT three weeks after receiving a single dose of the the BNT162b2 mRNA vaccine, 20 showed evidence of reduction in efficacy of antibodies against the B.1.1.7 mutant (Fig. 1b–c, Extended Data Fig. 3). The mean fold change reduction in sensitivity to first dose vaccine sera of B.1.1.7 compared to WT was approximately 3.2 (SD 5.7). The variation is likely due to the low neutralisation titres following first dose. Following the second dose, GMT was markedly increased compared with first dose titres, and the mean fold change had reduced to 1.9 (SD 0.9) (Fig. 1d–e). Amongst sera from 27 recovered individuals, the GMT at 50% neutralisation was 1334 for WT, significantly higher than post second dose vaccination (Fig. 1f–g). The fold change in ID50 for neutralisation of B.1.1.7 versus wild type (D614G) was 4.5 (Fig. 1f–g and Extended Data Fig. 4).
B.1.1.7 with spike E484K mutation and neutralization by vaccine and convalescent sera
The E484K substitution (Fig. 2a) is antigenically important, and has been reported as an escape mutation for several monoclonal antibodies including C121, C144, REGN10933 and Ly-CoV55522. E484K is also known to be present in the B.1.351 (501Y.V2) and P.1 (501Y.V3) lineages in combination with amino acid replacements at N501 and K417. As of 10th Feb 2021, twenty three English and two Welsh B.1.1.7 sequences from viral isolates contained the E484K substitution (Fig. 2b). The number of B.1.1.7 sequences has been increasing since the start of December 2020 (Fig. 2c). Phylogenetic analysis suggests that there have been multiple independent acquisitions, with one lineage appearing to expand over time, indicating active transmission (Fig. 2b). This has resulted in Public Health England naming this as a variant of concern (VOC 202102/02)23, triggering enhanced public health measures. There are as yet no phenotypic data on the sensitivity to neutralisation for this virus or its spike protein.
Figure 2. E484K appearing in background of B.1.1.7 with evidence of transmission.

a. Representation of Spike RBM:ACE2 interface (PDB: 6M0J) with residues E484, N501 and K417 highlighted as spheres coloured by element b. Maximum likelihood phylogeny of a subset of sequences from the United Kingdom bearing the E484K mutation (green) and lineage B.1.1.7 (blue), with background sequences without RBD mutations in black. As of 11th Feb 2021, 30 sequences from the B.1.1.7 lineage (one cluster of 25 at top of phylogenetic tree) have acquired the E484K mutation (red). c. Sequence accumulation over time in GISAID for UK sequences with B.1.1.7 and E484K. RBD – receptor binding domain; NTD – N terminal domain.
We therefore generated pseudoviruses bearing B.1.1.7 spike mutations with or without additional E484K and tested these against sera obtained after first and second dose mRNA vaccine as well as against convalescent sera. Following second dose, we observed a significant loss of neutralising activity for the pseudovirus with B.1.1.7 spike mutations and E484K (Fig 3d–e). The mean fold change for the E484K B.1.1.7 Spike was 6.7 compared to 1.9 for B.1.1.7, relative to WT (Fig. 3a–c). Similarly when we tested a panel of convalescent sera with a range of neutralisation titres (Fig. 1f–g), we observed additional loss of activity against the mutant B.1.1.7 spike with E484K, with fold change of 11.4 relative to WT (Fig. 3f–g).
Figure 3.
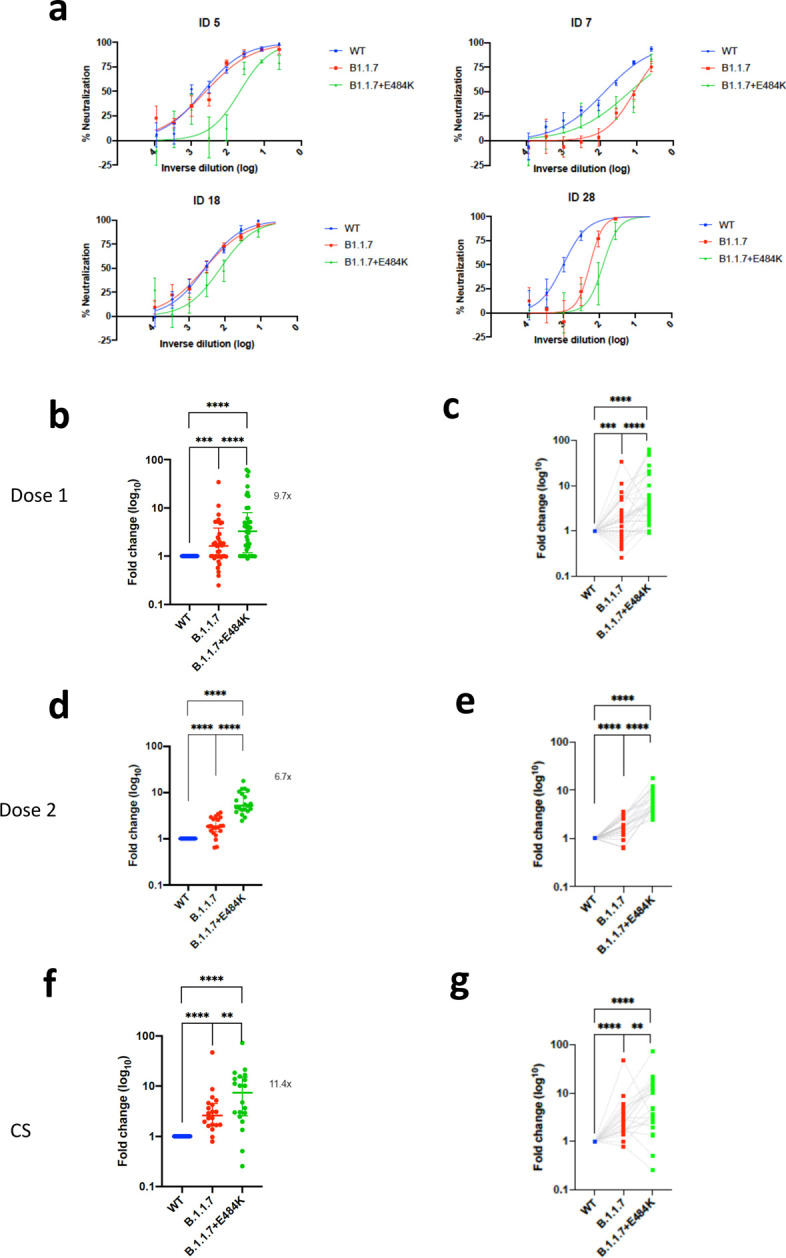
Neutralization potency of mRNA vaccine sera and convalescent sera (pre SARS-CoV-2 B.1.1.7) against pseudotyped virus bearing Spike mutations in the B1.1.7 lineage with and without E484K in the receptor binding domain (all In Spike D614G background). a, Example neutralization curves for vaccinated individuals. Data points represent mean of technical replicates with standard error and are representative of two independent experiments (b-g). 50% neutralisation titre for each virus against sera derived (b,c, n=37) following first vaccination (d,e, n=21) following second vaccination and (f,g, n=20) convalescent sera (CS) expressed as fold change relative to WT. Data points are mean fold change of technical replicates and are representative of two independent experiments. Central bar represents mean with outer bars representing s.d. Wilcoxon matched-pairs signed rank test p-values *<0.05, **<0.01, ***<0.001, ****<0.0001; ns not significant. Limit of detection for 50% neutralization set at 10.
B.1.1.7 variant escape from NTD- and RBM-specific mAb-mediated neutralization.
To investigate the role of the full set of mutations in NTD, RBD and S2 present in the B.1.1.7 variant, we tested 60 mAbs isolated from 15 individuals that recovered from SARS-CoV-2 infection in early 2020 with an in-vitro pseudotyped neutralization assay using VeroE6 target cells expressing Transmembrane protease serine 2 (TMPRSS2, Extended Data Table 1). We found that 20 out of 60 (33.3%) mAbs showed a greater than 2-fold loss of neutralising activity of B.1.1.7 variant compared to WT SARS-CoV-2 (Fig. 4a,b and Extended Data Fig. 5). Remarkably, the B.1.1.7 mutant virus was found to fully escape neutralization by 8 out of 10 NTD-targeting mAbs (80%), and partial escape from an additional mAb (10%) (Fig. 4c). We previously showed that the deletion of residue 144 abrogates binding by 4 out of 6 NTD-specific mAbs tested, possibly accounting for viral neutralization escape by most NTD-specific antibodies24. Of the 31 RBM-targeting mAbs, 5 (16.1%) showed more than 100-fold decrease in B.1.1.7 neutralization, and additional 6 mAbs (19.4%) had a partial 2-to-10-fold reduction (Fig. 4d). Finally, all RBD-specific non-RBM-targeting mAbs tested fully retained B.1.1.7 neutralising activity (Fig. 4e).
Figure 4. Neutralization and binding by a panel of NTD- and RBD-specific mAbs against WT, B.1.1.7 and RBD mutant SARS-CoV-2 viruses.

a, Neutralization of WT D614G (black), B.1.1.7 (blue) and a triple mutant (TM, carrying RBD mutations K417N/E484K/N501Y) (red) pseudotyped SARS-CoV-2-MLVs by 3 selected mAbs (S2E12, S2X333 and S2H14) from one representative experiment. Shown is the mean ± s.d. of 2 technical replicates. b, Neutralization of WT (D614G), B.1.1.7 and TM SARS-CoV-2-MLVs by 60 mAbs targeting NTD (n=10), RBM (n=31) and non-RBM sites in the RBD (n=19). Shown are the mean IC50 values (ng/ml) of n=2 independent experiments. c-e, Neutralization shown as mean IC50 values (upper panel) and mean fold change of B.1.1.7 (blue) or TM (red) relative to WT (lower panel) of NTD (c), RBM (d) and non-RBM (e) mAbs. Lower panel shows IC50 values from 2 independent experiments. f-h, Kinetics of binding of mAbs to WT (black), N501Y (blue) and E484K (red) RBD as measured by bio-layer interferometry (BLI). Shown in (f) are the 4 RBM-targeting mAbs with no reduced binding to N501Y or E484K RBD. Area under the curve (AUC) (g) and AUC fold change (h) of 50 mAbs tested against WT, N501Y and E484K RBD. mAbs with a >1.3 AUC fold change shown in blue and red. mAbs: monoclonal antibodies. NTD: N-terminal domain
To address the role of B.1.1.7 N501Y mutation in the neutralization escape from RBM-specific antibodies, we tested the binding of 50 RBD-specific mAbs to WT and N501Y mutant RBD by biolayer interferometry (Fig. 4f and Extended Data Fig. 6). The 5 RBM-specific mAbs that failed to neutralize B.1.1.7 variant (Fig. 4d) showed a complete loss of binding to N501Y RBD mutant (Fig. 4g–h), demonstrating a role for this mutation as an escape mechanism for certain RBM-targeting mAbs.
The decreased neutralising activity of the immune sera from vaccinees and convalescent patients against B.1.1.7, but not against Δ69/70–501Y-570D mutant (Fig. 1 and Extended Data Fig. 2), could be the result of a loss of neutralising activity of both RBD- and NTD-targeting antibodies, and suggests that the key mutation is Δ144. RBD antibodies against N501Y could play a role in decreased neutralisation by sera, with the overall impact possibly modulated by other mutations present in B.1.1.7, as well as the relative dominance of NTD versus RBM antibodies in polyclonal sera.
To assess the effect of E484K on this panel of mAbs we generated a SARS-CoV-2 pseudotype carrying the K417N, E484K and N501Y mutations (TM). The inclusion of the K417N substitution was prompted by the observation that substitutions at this position have been found in 5 sequences from recent viral isolates within the B.1.1.7 lineage (K417 to N, E or R). This is in keeping with convergent evolution of the virus towards an RBD with N501Y, E484K and K417N/T as evidenced by B.1.351 and P.1 lineages (K417N or K417T, respectively) causing great concern globally. It is therefore important to assess this combination going forward.
Importantly, mutations at K417 are reported to escape neutralization from mAbs, including the recently approved mAb LY-CoV01622,25. Out of the 60 mAbs tested, 20 (33.3%) showed >10 fold loss of neutralising activity of TM mutant compared to WT SARS-CoV-2 (Fig. 4 a–b and Extended Data Fig. 5), and of these 19 are RBM-specific mAbs. As above, we addressed the role of E484K mutation in escape from RBM-specific antibodies, by testing the binding of 50 RBD-specific mAbs to WT and E484K mutant RBD by biolayer interferometry (Fig. 4f and Extended Data Fig. 7). Out of the 19 RBM-specific mAbs that showed reduced or loss of neutralization of TM mutant (Fig. 4d), 16 showed a complete or partial loss of binding to E484K RBD mutant (Fig. 4g–h), consistent with findings that E484K is an important viral escape mutation26, 39, 27. Three of these 16 mAbs also lost binding to an RBD carrying N501Y, indicating that a fraction of RBM antibodies are sensitive to both N501Y and E484K mutations. Similarly, 3 of the 19 mAbs that lost neutralization of TM mutant (S2D8, S2H7 and S2X128) were previously shown to lose binding and neutralization to the K417V mutant, and here shown to be sensitive to either N501Y or E484K mutations.
SARS-CoV-2 B.1.1.7 binds human ACE2 with higher affinity than WT
SARS-CoV-2 and SARS-CoV enter host cells through binding of the S glycoprotein to angiotensin converting enzyme 2 (ACE2)1,28. Previous studies showed that the binding affinity of SARS-CoV for human ACE2 correlated with the rate of viral replication in distinct species, transmissibility and disease severity29–31. However, the picure is unclear for SARS-CoV-2. To understand the potential contribution of receptor interaction to infectivity, we set out to evaluate the influence of the B.1.1.7 RBD substitution N501Y on receptor engagement. We used biolayer interferometry to study binding kinetics and affinity of the purified human ACE2 ectodomain (residues 1–615) to immobilized biotinylated SARS-CoV-2 B.1.1.7 or WT RBDs. We found that ACE2 bound to the B.1.1.7 RBD with an affinity of 22 nM compared to 133 nM for the WT RBD (Extended Data Fig. 8), in agreement with our previous deep-mutational scanning measurements using dimeric ACE232. Although ACE2 bound with comparable on-rates to both RBDs, the observed dissociation rate constant was slower for B.1.1.7 than for the WT RBD (Table 1).
Table 1.
Kinetic analysis of human ACE2 binding to SARS-CoV-2 Wuhan-1, N501Y and N501Y/E484K/K417N (TM) RBDs by biolayer interferometry.
| SARS-CoV-2 RBD WT | SARS-CoV-2 RBD N501Y | SARS-CoV-2 RBD TM | ||
|---|---|---|---|---|
| KD (nM) | hACE2 | 133 | 22 | 64 |
| kon (M−1.s−1) | 1.3*105 | 1.4*105 | 1.3*105 | |
| koff (s−1) | 1.8*10−2 | 3*10−3 | 8.5*10−3 |
Values reported represent the global fit to the data shown in Extended Data Fig. 8.
To understand the impact of TM mutations (K417N, E484K and N501Y), we evaluated binding of ACE2 to the immobilized TM RBD using biolayer interferometry. We determined an ACE2 binding affinity of 64 nM for the TM RBD which is driven by a faster off-rate than observed for the B.1.1.7 RBD but slower than for the WT RBD. Based on our previous deep-mutational scanning measurements using dimeric ACE2, we propose that the K417N mutation is slightly detrimental to ACE2 binding explaining the intermediate affinity determined for the TM RBD compared to the B.1.17 and WT RBDs, likely as a result of disrupting the salt bridge formed with ACE2 residue D30. Enhanced binding of the B.1.1.7 RBD to human ACE2 resulting from the N501Y mutation might participate in the efficient ongoing transmission of this newly emergent SARS-CoV-2 lineage, and possibly reduced opportunity for antibody binding. Although the TM RBD mutations found in B.1.351 are known to participate in immune evasion33,34, the possible contribution to transmissibility of enhanced ACE2 binding relative to WT remains to be determined for this lineage.
Discussion
Serum neutralising activity is a correlate of protection for other respiratory viruses, including influenza35 and respiratory syncytial virus where prohylaxis with monoclonal antibodies has been used in at-risk groups36,37. Neutralising antibody titres appeared to be highly correlated with vaccine protection against SARS-CoV-2 rechallenge in non-human primates, and importantly, there was no correlation between T cell responses (as measured by ELISpot) and protection38. Moreover, passive transfer of purified polyclonal IgGs from convalescent macaques protected naïve macaques against subsequent SARS-CoV-2 challenge39. Coupled with multiple reports of re-infection, there has therefore been significant attention placed on virus neutralisation.
This study reports on the neutralisation by sera collected after both the first and second doses of the BNT162b2 vaccine. The participants of this study were older adults, in line with the targeting of this age group in the initial rollout of the vaccination campaign in the UK. Participants showed similar neutralising activity against wild type pseudovirus as in the phase I/II study12. This is relevant for the UK and other countries planning to extend the gap between doses of mRNA and adenovirus based vaccines from 3 to 12 weeks, despite lack of data for this schedule for mRNA vaccines in particular.
The three mutations in S1 (N501Y, A570D, ΔH69/V70) did not appear to impact neutralisation in a pseudovirus assay, consistent with data on N501Y having little effect on nuetralisation by convalescent and post vaccination sera40. However, we demonstrated that a pseudovirus bearing S protein with the full set of mutations present in the B.1.1.7 variant (i.e., ΔH69/V70, Δ144, N501Y, A570D, P681H, T716I, S982A, D1118H) did result in small reduction in neutralisation by sera from vaccinees that was more marked following the first dose than the second dose. This could be related to increased breadth/potency/concentration of antibodies following the boost dose. A reduction in neutralization titres from mRNA-elicited antibodies in volunteers who received two doses (using both mRNA-1273 and BNT162b2 vaccines) was also observed by Wang et al.41 using pseudoviruses carrying the N501Y mutation. Other studies also reported small reduction of neutralization against the B.1.1.7 variant against sera from individuals vaccinated with two doses of BNT162b242 and mRNA-127343. Xie et al did not find an effect of N501Y alone in the context of BNT162b2 vaccine sera44.
The reduced neutralising activity observed with polyclonal antibodies elicited by mRNA vaccines observed in this study is further supported by the loss of neutralising activity observed with human mAbs directed to both RBD and, to a major extent, to NTD. In the study by Wang et al., 6 out 17 RDB-specific mAbs isolated from mRNA-1273 vaccinated individuals showed more than 100-fold neutralisation loss against N501Y mutant, a finding that is consistent with the loss of neutralisation by 5 out 29 RBM-specific mAbs described in this study. However, the contribution of N501Y to loss of neutralisation activity of polyclonal vaccine and convalescent sera is less clear, and interactions with other mutations likely.
Multiple variants, including the 501Y.V2 and B.1.1.7 lineages, harbor multiple mutations as well as deletions in NTD, most of which are located in a site of vulnerability that is targeted by all known NTD-specific neutralising antibodies24,45. The role of NTD-specific neutralising antibodies might be under-estimated, in part by the use of neutralization assays based on target cells over-expressing ACE2 receptor. NTD-specific mAbs were suggested to interfere with viral entry based on other accessory receptors, such as DC-SIGN and L-SIGN46, and their neutralization potency was found to be dependent on different in vitro culture conditions24. The observation that 9 out of 10 NTD-specific neutralising antibodies failed to show a complete or near-complete loss of neutralising activity against B.1.1.7 indicates that this new variant may have evolved also to escape from this class of antibodies, that may have a yet unrecognized role in protective immunity. Wibmer et al.34 have also recently reported the loss of neutralization of 501Y.V2 by the NTD-specific mAb 4A8, likely driven by the R246I mutation. This result is in line with the lack of neutralization of B.1.1.7 by the 4A8 mAb observed in this study, likely caused by Δ144 due to loss of binding24. Finally, the role of NTD mutations (in particular, L18F, Δ242–244 and R246I) was further supported by the marked loss of neutralization observed by Wibmer et al.34 against 501Y.V2 compared to the chimeric pseudotyped viral particle carrying only the RBD mutations K417N, E484K and N501Y. Taken together, the presence of multiple escape mutations in NTD is supportive of the hypothesis that this region of the spike, in addition to RBM, is also under immune pressure.
Worryingly, we have shown that there are multiple B.1.1.7 sequences in the UK bearing E484K with early evidence of transmission as well as independent aquisitions. We measured further reduction neutralisation titers by vaccine sera when E484K was present alongside the B.1.1.7 S mutations. Wu and co-authors43 have also shown that variants carrying the E484K mutation resulted in 3-to-6 fold reduction in neutralization by sera from mRNA-1273 vaccinated individuals. Consistently, in this study we found that approximately 50% of the RBM mAbs tested lost neutralising activity against SARS-CoV-2 carrying E484K. E484K has been shown to impact neutralisation by monoclonal antibodies or convalescent sera, especially in combination with N501Y and K417N16,26,47–49. Wang et al also showed reduced neutralisation by mRNA vaccine sera against E484K bearing pseudovirus34.
Evidence for the importance role of NTD deletions in combination with E484K in immune escape is provided by Andreano et al.27 who describe the emergence of Δ140 in virus co-incubated with potently neutralising convalescent plasma, causing a 4-fold reduction in neutralization titre. This Δ140 mutant subsequently acquired E484K which resulted in a further 4-fold drop in neutralization titre indicating a two residue change across NTD and RBD represents an effective pathway of escape that can dramatically inhibit the polyclonal response.
Our study was limited by modest sample size. Although the spike pseudotyping system has been shown to faithfully represent full length infectious virus, there may be determinants outside the S that influence escape from antibody neutralization either directly or indirectly in a live replication competent system. On the other hand live virus systems allow replication and therefore mutations to occur, and rigorous sequencing at multiple steps is needed.
Vaccines are a key part of a long term strategy to bring SARS-CoV-2 transmission under control. Our data suggest that vaccine escape to current Spike directed vaccines designed against the Wuhan strain will be inevitable, particularly given that E484K is emerging independently and recurrently on a B.1.1.7 (501Y.V1) background, and given the rapid global spread of B.1.1.7. Other major variants with E484K such as 501Y.V2 and V3 are also spreading regionally. This should be mitigated by designing next generation vaccines with mutated S sequences and using alternative viral antigens.
MATERIALS AND METHODS
Participant recruitment and ethics
Participants who had received the first dose of vaccine and individuals with COVID-19 (Coronavirus Disease-19) were consented into the COVID-19 cohort of the NIHR Bioresource. The study was approved by the East of England – Cambridge Central Research Ethics Committee (17/EE/0025).
SARS-CoV-2 serology by multiplex particle-based flow cytometry (Luminex):
Recombinant SARS-CoV-2 N, S and RBD were covalently coupled to distinct carboxylated bead sets (Luminex; Netherlands) to form a 3-plex and analyzed as previously described (Xiong et al. 2020). Specific binding was reported as mean fluorescence intensities (MFI). Linear regression was used to explore the association between antibody response, T cell response and serum neutralisation in Stata 13. The Pearson correlation coefficient was reported.
Recombinant expression of SARS-CoV-2-specific mAbs.
Human mAbs were isolated from plasma cells or memory B cells of SARS-CoV-2 immune donors, as previously described50–52. Recombinant antibodies were expressed in ExpiCHO cells at 37°C and 8% CO2. Cells were transfected using ExpiFectamine. Transfected cells were supplemented 1 day after transfection with ExpiCHO Feed and ExpiFectamine CHO Enhancer. Cell culture supernatant was collected eight days after transfection and filtered through a 0.2 μm filter. Recombinant antibodies were affinity purified on an ÄKTA xpress fast protein liquid chromatography (FPLC) device using 5 mL HiTrap™ MabSelect™ PrismA columns followed by buffer exchange to Histidine buffer (20 mM Histidine, 8% sucrose, pH 6) using HiPrep 26/10 desalting columns
Generation of S mutants
Amino acid substitutions were introduced into the D614G pCDNA_SARS-CoV-2_S plasmid as previously described53 using the QuikChange Lightening Site-Directed Mutagenesis kit, following the manufacturer’s instructions (Agilent Technologies, Inc., Santa Clara, CA). Sequences were checked by Sanger sequencing.
Preparation of B.1.1.7 or TM SARS-CoV-2 S glycoprotein-encoding-plasmid used to produce SARS-CoV-2-MLV based on overlap extension PCR. Briefly, a modification of the overlap extension PCR protocol54 was used to introduce the nine mutations of the B.1.1.7 lineage or the three mutations in TM mutant in the SARS-CoV-2 S gene. In a first step, 9 DNA fragments with overlap sequences were amplified by PCR from a plasmid (phCMV1, Genlantis) encoding the full-length SARS-CoV-2 S gene (BetaCoV/Wuhan-Hu-1/2019, accession number mn908947). The mutations (del-69/70, del-144, N501Y, A570D, D614G, P681H, S982A, T716I and D1118H or K417N, E484K and N501Y) were introduced by amplification with primers with similar Tm. Deletion of the C-terminal 21 amino acids was introduced to increase surface expression of the recombinant S55. Next, 3 contiguous overlapping fragments were fused by a first overlap PCR (step 2) using the utmost external primers of each set, resulting in 3 larger fragments with overlapping sequences. A final overlap PCR (step 3) was performed on the 3 large fragments using the utmost external primers to amplify the full-length S gene and the flanking sequences including the restriction sites KpnI and NotI. This fragment was digested and cloned into the expression plasmid phCMV1. For all PCR reactions the Q5 Hot Start High fidelity DNA polymerase was used (New England Biolabs Inc.), according to the manufacturer’s instructions and adapting the elongation time to the size of the amplicon. After each PCR step the amplified regions were separated on agarose gel and purified using Illustra GFX™ PCR DNA and Gel Band Purification Kit (Merck KGaA).
Pseudotype virus preparation
Viral vectors were prepared by transfection of 293T cells by using Fugene HD transfection reagent (Promega). 293T cells were transfected with a mixture of 11ul of Fugene HD, 1μg of pCDNAΔ19spike-HA, 1ug of p8.91 HIV-1 gag-pol expression vector56,57, and 1.5μg of pCSFLW (expressing the firefly luciferase reporter gene with the HIV-1 packaging signal). Viral supernatant was collected at 48 and 72h after transfection, filtered through 0.45um filter and stored at −80°C. The 50% tissue culture infectious dose (TCID50) of SARS-CoV-2 pseudovirus was determined using Steady-Glo Luciferase assay system (Promega).
Serum/plasma pseudotype neutralization assay
Spike pseudotype assays have been shown to have similar characteristics as neutralisation testing using fully infectious wild type SARS-CoV-220. Virus neutralisation assays were performed on 293T cell transiently transfected with ACE2 and TMPRSS2 using SARS-CoV-2 spike pseudotyped virus expressing luciferase58. Pseudotyped virus was incubated with serial dilution of heat inactivated human serum samples or sera from vaccinees in duplicate for 1h at 37°C. Virus and cell only controls were also included. Then, freshly trypsinized 293T ACE2/TMPRSS2 expressing cells were added to each well. Following 48h incubation in a 5% CO2 environment at 37°C, luminescence was measured using the Steady-Glo or Bright-Glo Luciferase assay system (Promega). Neutralization was calculated relative to virus only controls. Dilution curves were presented as a mean neutralization with standard error of the mean (SEM). ID50 values were calculated in GraphPad Prism. The ID50 withing groups were summarised as a geometric mean titre and statistical comparison between groups were made with Wilxocon ranked sign test. In addition, the impact of the mutations on the neutralising effect of the sera were expressed as fold change (FC) of ID50 of the wild-type compared to mutant pseudotyped virus. Statistical difference in the mean FC between groups was determined using a 2-tailed t-test.
IFNγ FluoroSpot assays
Frozen PBMCs were rapidly thawed, and the freezing medium was diluted into 10ml of TexMACS media (Miltenyi Biotech), centrifuged and resuspended in 10ml of fresh media with 10U/ml DNase (Benzonase, Merck-Millipore via Sigma-Aldrich), PBMCs were incubated at 37°C for 1h, followed by centrifugation and resuspension in fresh media supplemented with 5% Human AB serum (Sigma Aldrich) before being counted. PBMCs were stained with 2ul of each antibody: anti-CD3-fluorescein isothiocyanate (FITC), clone UCHT1; anti-CD4-phycoerythrin (PE), clone RPA-T4; anti-CD8a-peridinin-chlorophyll protein - cyanine 5.5 (PerCP Cy5.5), clone RPA-8a (all BioLegend, London, UK), LIVE/DEAD Fixable Far Red Dead Cell Stain Kit (Thermo Fisher Scientific). PBMC phenotyping was performed on the BD Accuri C6 flow cytometer. Data were analysed with FlowJo v10 (Becton Dickinson, Wokingham, UK). 1.5 to 2.5 × 105 PBMCs were incubated in pre-coated Fluorospot plates (Human IFNγ FLUOROSPOT (Mabtech AB, Nacka Strand, Sweden)) in triplicate with peptide mixes specific for Spike, Nucleocapsid and Membrane proteins of SARS-CoV-2 (final peptide concentration 1μg/ml/peptide, Miltenyi Biotech) and an unstimulated and positive control mix (containing anti-CD3 (Mabtech AB), Staphylococcus Enterotoxin B (SEB), Phytohaemagglutinin (PHA) (all Sigma Aldrich)) at 37°C in a humidified CO2 atmosphere for 48 hours. The cells and medium were decanted from the plate and the assay developed following the manufacturer’s instructions. Developed plates were read using an AID iSpot reader (Oxford Biosystems, Oxford, UK) and counted using AID EliSpot v7 software (Autoimmun Diagnostika GmbH, Strasberg, Germany). All data were then corrected for background cytokine production and expressed as spot forming units (SFU)/Million PBMC or CD3 T cells. The association between spike Tcell response, spike specific antibody response and serum neutralisation was deterimined using linear regression and the Pearson correlation coefficient between these variables were determined using Stata 13.
Ab discovery and recombinant expression
Human mAbs were isolated from plasma cells or memory B cells of SARS-CoV or SARS-CoV-2 immune donors, as previously described48,56–58. Recombinant antibodies were expressed in ExpiCHO cells at 37°C and 8% CO2. Cells were transfected using ExpiFectamine. Transfected cells were supplemented 1 day after transfection with ExpiCHO Feed and ExpiFectamine CHO Enhancer. Cell culture supernatant was collected eight days after transfection and filtered through a 0.2 μm filter. Recombinant antibodies were affinity purified on an ÄKTA xpress FPLC device using 5 mL HiTrap™ MabSelect™ PrismA columns followed by buffer exchange to Histidine buffer (20 mM Histidine, 8% sucrose, pH 6) using HiPrep 26/10 desalting columns.
MAbs pseudovirus neutralization assay
MLV-based SARS-CoV-2 S-glycoprotein-pseudotyped viruses were prepared as previously described (Pinto et al., 2020). HEK293T/17cells were cotransfected with a WT, B.1.1.7 or TM SARS-CoV-2 spike glycoprotein-encoding-plasmid, an MLV Gag-Pol packaging construct and the MLV transfer vector encoding a luciferase reporter using X-tremeGENE HP transfection reagent (Roche) according to the manufacturer’s instructions. Cells were cultured for 72 h at 37°C with 5% CO2 before harvesting the supernatant. VeroE6 stably expressing human TMPRSS2 were cultured in Dulbecco’s Modified Eagle’s Medium (DMEM) containing 10% fetal bovine serum (FBS), 1% penicillin–streptomycin (100 I.U. penicillin/mL, 100 μg/mL), 8 μg/mL puromycin and plated into 96-well plates for 16–24 h. Pseudovirus with serial dilution of mAbs was incubated for 1 h at 37°C and then added to the wells after washing 2 times with DMEM. After 2–3 h DMEM containing 20% FBS and 2% penicillin–streptomycin was added to the cells. Following 48–72 h of infection, Bio-Glo (Promega) was added to the cells and incubated in the dark for 15 min before reading luminescence with Synergy H1 microplate reader (BioTek). Measurements were done in duplicate and relative luciferase units were converted to percent neutralization and plotted with a non-linear regression model to determine IC50 values using GraphPad PRISM software (version 9.0.0).
Antibody binding measurements using bio-layer interferometry (BLI)
MAbs were diluted to 3 μg/ml in kinetic buffer (PBS supplemented with 0.01% BSA) and immobilized on Protein A Biosensors (FortéBio). Antibody-coated biosensors were incubated for 3 min with a solution containing 5 μg/ml of WT, N501Y or E484K SARS-CoV-2 RBD in kinetic buffer, followed by a 3-min dissociation step. Change in molecules bound to the biosensors caused a shift in the interference pattern that was recorded in real time using an Octet RED96 system (FortéBio). The binding response over time was used to calculate the area under the curve (AUC) using GraphPad PRISM software (version 9.0.0).
Production of SARS-CoV-2 and B.1.1.7 receptor binding domains and human ACE2
The SARS-CoV-2 RBD (BEI NR-52422) construct was synthesized by GenScript into CMVR with an N-terminal mu-phosphatase signal peptide and a C-terminal octa-histidine tag (GHHHHHHHH) and an avi tag. The boundaries of the construct are N-328RFPN331 and 528KKST531-C59. The B.1.1.7 RBD gene was synthesized by GenScript into pCMVR with the same boundaries and construct details with a mutation at N501Y. These plasmids were transiently transfected into Expi293F cells using Expi293F expression medium (Life Technologies) at 37°C 8% CO2 rotating at 150 rpm. The cultures were transfected using PEI cultivated for 5 days. Supernatants were clarified by centrifugation (10 min at 4000×g) prior to loading onto a nickel-NTA column (GE). Purified protein was biotinylated overnight using BirA (Biotin ligase) prior to size exclusion chromatography (SEC) into phosphate buffered saline (PBS). Human ACE2-Fc (residues 1–615 with a C-terminal thrombin cleavage site and human Fc tag) were synthesized by Twist. Clarified supernatants were affinity purified using a Protein A column (GE LifeSciences) directly neutralized and buffer exchanged. The Fc tag was removed by thrombin cleavage in a reaction mixture containing 3 mg of recombinant ACE2-FC ectodomain and 10 μg of thrombin in 20 mM Tris-HCl pH8.0, 150 mM NaCl and 2.5 mM CaCl2.The reaction mixture was incubated at 25°C overnight and re-loaded on a Protein A column to remove uncleaved protein and the Fc tag. The cleaved protein was further purified by gel filtration using a Superdex 200 column 10/300 GL (GE Life Sciences) equilibrated in PBS.
Protein affinity measurement using bio-layer interferometry
Biotinylated RBD (WT, N501Y, or TM) were immobilized at 5 ng/uL in undiluted 10X Kinetics Buffer (Pall) to SA sensors until a load level of 1.1nm. A dilution series of either monomeric ACE2 or Fab in undiluted kinetics buffer starting at 1000–50nM was used for 300–600 seconds to determine protein-protein affinity. The data were baseline subtracted and the plots fitted using the Pall FortéBio/Sartorius analysis software (version 12.0). Data were plotted in Prism.
Statistical analysis
Linear regression was used to explore the association between antibody response, T cell response and serum neutralisation in Stata 13. The Pearson correlation coefficient was reported.
Neutralisation data analysis
Neutralization was calculated relative to virus only controls. Dilution curves were presented as a mean neutralization with standard error of the mean (SEM). IC50 values were calculated in GraphPad Prism. The inhibitory dilution (ID50) within groups were summarised as a geometric mean titre and statistical comparison between groups were made with Wilxocon ranked sign test. In addition, the impact of the mutations on the neutralising effect of the sera were expressed as fold change of ID50 of the wild-type compared to mutant pseudotyped virus. Statistical difference in the mean FC between groups was determined using a 2-tailed t-test
IFNγ FluoroSpot assay data analysis
The association between spike Tcell response, spike specific antibody response and serum neutralisation was determined using linear regression and the Pearson correlation coefficient between these variables were determined using Stata 13.
Data availability.
The neutralization and BLI data shown in Fig. 4 and Extended Data Fig. 5–7 can be found in Source Data Fig. 4. Other data are available from the corresponding author on request.
Extended Data
Extended Data Figure 1: Immune responses three weeks after first dose of Pfizer SARS-CoV-2 vaccine BNT162b2.

a, Serum IgG responses against N protein, Spike and the Spike Receptor Binding Domain (RBD) from first vaccine participants (green), recovered COVID-19 cases (red), 3 convalescent plasma units and healthy controls (grey) as measured by a flow cytometry based Luminex assay. MFI, mean fluorescence intensity. Geometric mean titre (GMT with standard deviation (s.d) of two technical repeats presented. b, Relationship between serum IgG responses as measured by flow cytometry and serum neutralisation ID50. c, Relationship between serum neutralisation ID50 and T cell responses against SARS-CoV-2 by IFN gamma ELISpot. SFU: spot forming units. d, Relationship between serum IgG responses and T cell responses. Simple linear regression is presented with Pearson correlation (r), P-value (p) and regression coefficient/slope (β).
Extended data Fig 2. Neutralization by first dose BNT162b2 vaccine and convalescent sera against wild type and mutant (N501Y, A570D, ΔH69/V70) SARS-CoV-2 pseudotyped viruses:

(a-b) Vaccine sera dilution for 50% neutralization against WT and Spike mutant with N501Y, A570D, ΔH69/V70. Geometric mean titre (GMT) + s.d of two independent experiments with two technical repeats presented. (c-d) Convalescent sera dilution for 50% neutralization against WT and Spike mutant with N501Y, A570D, ΔH69/V70. GMT + s.d of representative experiment with two technical repeats presented. e, Representative curves of convalescent serum log10 inverse dilution against % neutralization for WT v N501Y, A570D, ΔH69/V70. Where a curve is shifted to the right this indicates the virus is less sensitive to the neutralizing antibodies in the serum. Data are means of technical replicates and error bars represent standard error of the mean. Data are representative of 2 independent experiments. Limit of detection for 50% neutralization set at 10.
Extended Data Fig. 3. Representative neutralization curves of BNT162b2 vaccine sera against pseudovirus virus bearing eight Spike mutations present in B.1.1.7 versus wild type (all In Spike D614G background).

Indicated is serum log10 inverse dilution against % neutralization. Where a curve is shifted to the right this indicates the virus is less sensitive to the neutralizing antibodies in the serum. Data are for first dose of vaccine (D1). Data points represent means of technical replicates and error bars represent standard error of the mean. Limit of detection for 50% neutralization set at 10.
Extended Data Fig. 4. Representative neutralization curves of convalescent sera against wild type and B.1.1.7 Spike mutant SARS-CoV-2 pseudoviruses.

Indicated is serum log10 inverse dilution against % neutralization. Where a curve is shifted to the right this indicates the virus is less sensitive to the neutralizing antibodies in the serum. Data points represent means of technical replicates and error bars represent standard error of the mean. Limit of detection for 50% neutralization set at 10.
Extended Data Fig. 5.

Neutralisation of WT (D614G), B.1.1.7 and TM (N501Y, E484K, K417N) SARS-CoV-2 Spike pseudotyped virus by a panel of 57 monoclonal antibodies (mAbs). a-c, Neutralisation of WT (black), B.1.1.7 (blue) and TM (red) SARS-CoV-2-MLV by 9 NTD-targeting (a), 29 RBM-targeting (b) and 19 non-RBM-targeting (c) mAbs.
Extended Data Fig. 6. Kinetics of binding to WT and N501Y SARS-CoV-2 RBD of 43 RBD-specific mAbs.

a-b, Binding to WT (black) and N501Y (blue) RBD by 22 RBM-targeting (a) and 21 non-RBM-targeting (b) mAbs. An antibody of irrelevant specificity was included as negative control. mAbs: monoclonal antibodies
Extended Data Fig. 7. Kinetics of binding to WT and E484K SARS-CoV-2 RBD of 46 RBD-specific mAbs.

a-b, Binding to WT (black) and E484K (red) RBD by 27 RBM-targeting (a) and 19 non-RBM-targeting (b) mAbs. An antibody of irrelevant specificity was included as negative control. mAbs: monoclonal antibodies
Extended Data Fig. 8. Binding of human ACE2 to SARS-CoV-2 WT, N501Y, TM (N501Y, E484K, K417N) RBDs.
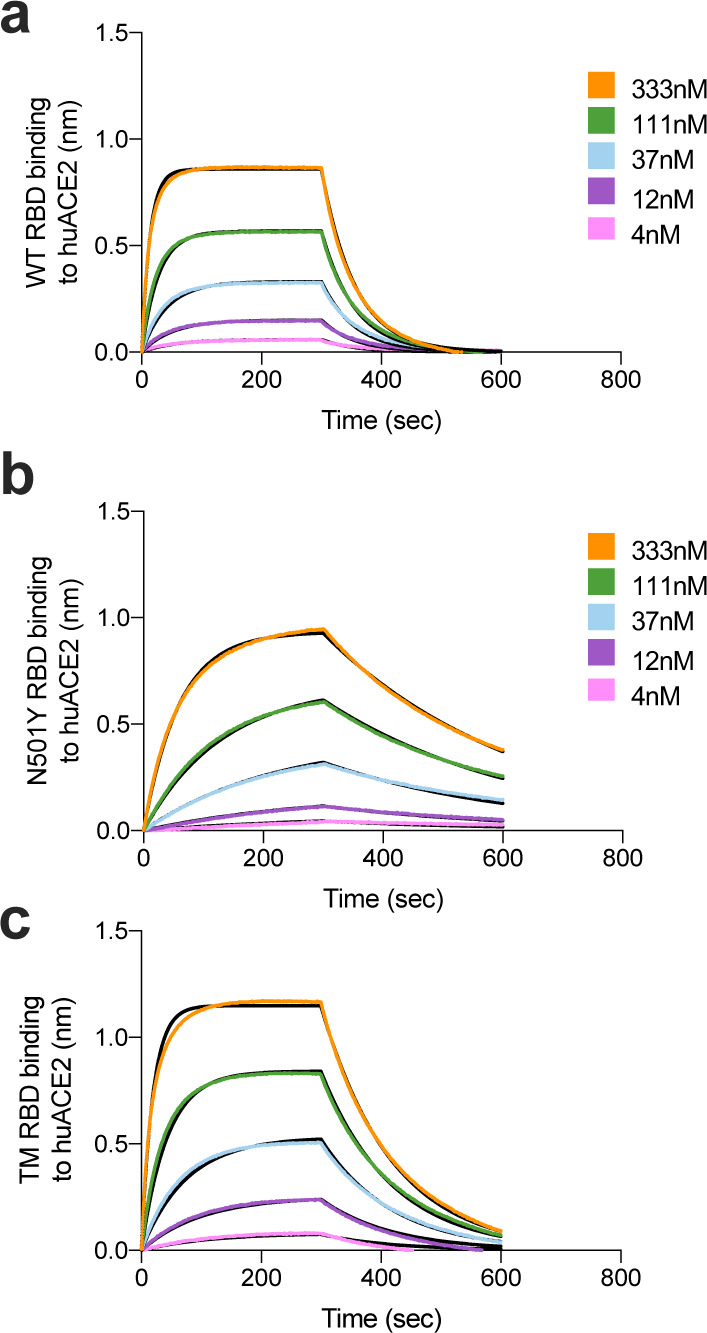
a-b. BLI binding analysis of the human ACE2 ectodomain (residues 1–615) to immobilized SARS-CoV-2 WT RBD (a) and B.1.1.7 RBD (c). Black lines correspond to a global fit of the data using a 1:1 binding model. RBD: receptor binding domain.
Extended Data Table 1.
Neutralization, V gene usage and other properties of tested mAbs.
| mAb | Domain (site) | VH usage (% id.) | Source (DSO) | IC50 WT (ng/ml) | IC50 B.1.1.7 (ng/ml) | ACE2 blocking | SARS-CoV | Escape residues | Ref. |
|---|---|---|---|---|---|---|---|---|---|
| 4A8 | NTD (i) | 1–24 | N/A | 38 | Neg. | S12P; C136Y; Y144del; H146Y; K147T; R246A | 60 | ||
| S2L26 | NTD (i) | 1–24 (97.2) | Hosp. (52) | 70 | - | Neg. | N/A | 24 | |
| S2L50 | NTD (i) | 4–59 (95.4) | Hosp. (52) | 264 | 50 | Neg. | N/A | 24 | |
| S2M28 | NTD (i) | 3–33 (97.6) | Hosp. (46) | 295 | 12′207 | Neg. | P9S/Q;S12P; C15F/R; L18P; Y28C; A123T; C136Y; G142D; Y144del; K147Q/T; R246G;P251L; G252C | 24 | |
| S2X107 | NTD (i) | 4-38-2 (97) | Sympt. (75) | 388 | Neg. | N/A | 24 | ||
| S2X124 | NTD (i) | 3–30 (99) | Sympt. (75) | 221 | - | Neg. | - | N/A | 24 |
| S2X158 | NTD (i) | 1–24 (96.3) | Sympt. (75) | 56 | - | Neg. | - | N/A | 24 |
| S2X28 | NTD (i) | 3–30 (97.9) | Sympt. (48) | 1’399 | Neg. | P9S; S12P; C15W; L18P; C136G/Y; F140S; L141S; G142C/D; Y144C/N; K147T/Q/E; R158G; L244S; R246G | 24 | ||
| S2X303 | NTD (i) | 2–5 (95.9) | Sympt. (125) | 69 | - | Neg. | - | N/A | 24 |
| S2X333 | NTD (i) | 3–33 (96.5) | Sympt. (125) | 66 | Neg. | P9L; S12P; C15S/Y; L18P; C136G/Y; F140C; G142D; K147T | 24 | ||
| S2D106 | RBD (I/RBM) | 1–69 (97.2) | Hosp. (98) | 27 | 20 | Strong | - | N/A | 8 |
| S2D19 | RBD (I/RBM) | 4–31 (99.7) | Hosp. (49) | 128 | 75’200 | Moderate | - | N/A | 8 |
| S2D32 | RBD (I/RBM) | 3–49 (98.3) | Hosp. (49) | 26 | 11 | Strong | - | N/A | 8 |
| S2D65 | RBD (I/RBM) | 3–9 (96.9) | Hosp. (49) | 24 | 12 | Weak | N/A | 8 | |
| S2D8 | RBD (I/RBM) | 3–23 (96.5) | Hosp. (49) | 27 | 58′644 | Strong | - | N/A | 8 |
| S2D97 | RBD (I/RBM) | 2–5 (96.9) | Hosp. (98) | 20 | 17 | Weak | N/A | 8 | |
| S2E11 | RBD (I/RBM) | 4–61 (98.3) | Hosp. (51) | 27 | 16 | Weak | - | N/A | 8 |
| S2E12 | RBD (I/RBM) | 1–58 (97.6) | Hosp. (51) | 27 | 31 | Strong | - | G476S (3x) | 8,61 |
| S2E13 | RBD (I/RBM) | 1–18(96.2) | Hosp. (51) | 34 | 77 | Strong | - | N/A | 8 |
| S2E16 | RBD (I/RBM) | 3–30 (98.3) | Hosp. (51) | 36 | 38 | Strong | - | N/A | 8 |
| S2E23 | RBD (I/RBM) | 3–64 (96.9) | Hosp. (51) | 139 | 180 | Strong | - | N/A | 8 |
| S2H14 | RBD (I/RBM) | 3–15(100) | Sympt. (17) | 460 | 64′463 | Weak | - | N/A | 8,62 |
| S2H19 | RBD (I/RBM) | 3–15(98.6) | Sympt. (45) | 239 | - | Weak | - | N/A | 8 |
| S2H58 | RBD (I/RBM) | 1–2(97.9) | Sympt. (45) | 27 | 14 | Strong | - | N/A | 8 |
| S2H7 | RBD (I/RBM) | 3–66 (98.3) | Sympt. (17) | 492 | 573 | Weak | N/A | 8 | |
| S2H70 | RBD (I/RBM) | 1–2 (99) | Sympt. (45) | 147 | 65 | Weak | - | N/A | 8 |
| S2H71 | RBD (I/RBM) | 2–5 (99) | Sympt. (45) | 36 | 9 | Moderate | - | N/A | 8 |
| S2M11 | RBD (I/RBM) | 1–2 (96.5) | Hosp. (46) | 11 | 4 | Weak | Y449N; L455F; E484K; E484Q; F490L; F490S; S494P | 8,61 | |
| S2N12 | RBD (I/RBM) | 4–39 (97.6) | Hosp. (51) | 76 | 40 | Strong | - | N/A | 8 |
| S2N22 | RBD (I/RBM) | 3–23 (96.5) | Hosp. (51) | 32 | 21 | Strong | N/A | 8 | |
| S2N28 | RBD (I/RBM) | 3–30 (97.2) | Hosp. (51) | 72 | 21 | Strong | - | N/A | 8 |
| S2X128 | RBD (I/RBM) | 1-69-2 (97.6) | Sympt. (75) | 50 | 112 | Strong | N/A | 8 | |
| S2X16 | RBD (I/RBM) | 1–69 (97.6) | Sympt. (48) | 45 | 103 | Strong | N/A | 8 | |
| S2X192 | RBD (I/RBM) | 1–69 (96.9) | Sympt. (75) | 326 | - | Weak | - | N/A | 8 |
| S2X227 | RBD (I/RBM) | 1–46 (97.9) | Sympt. (75) | 26 | 14 | Strong | N/A | ||
| S2X246 | RBD (I/RBM) | 3–48 (96.2) | Sympt. (75) | 35 | 30 | Strong | N/A | ||
| S2X30 | RBD (I/RBM) | 1–69 (97.9) | Sympt. (48) | 32 | 53 | Strong | - | N/A | 8 |
| S2X324 | RBD (I/RBM) | 2–5 (97.3) | Sympt. (125) | 8 | 23 | Strong | - | N/A | |
| S2X58 | RBD (I/RBM) | 1–46 (99) | Sympt. (48) | 32 | 47 | Strong | N/A | 8 | |
| S2H90 | RBD (II) | 4–61 (96.6) | Sympt. (81) | 77 | 32 | Strong | + | N/A | 8 |
| S2H94 | RBD (II) | 3–23 (93.4) | Sympt. (81) | 123 | 144 | Strong | + | N/A | 8 |
| S2H97 | RBD (V) | 5–51 (98.3) | Sympt. (81) | 513 | 248 | Weak | + | N/A | |
| S2K15 | RBD (II) | 2–26 (99.3) | Sympt. (87) | 361 | 235 | 0 | + | N/A | |
| S2K21 | RBD (II) | 3–33 (96.2) | Sympt. (118) | 201 | 189 | 0 | + | N/A | |
| S2K30 | RBD (II) | 1–2(97.2) | Sympt. (87) | 185 | 134 | 0 | + | N/A | |
| S2K63v2 | RBD (II) | 3-30-3 (95.6) | Sympt. (118) | 144 | 215 | 0 | + | N/A | |
| S2L17 | RBD (?) | 5-10-1(98.3) | Hosp. (51) | 313 | 127 | Moderate | + | N/A | 8 |
| S2L49 | RBD (?) | 3–30 (97.9) | Hosp. (51) | 24 | 32 | Neg. | + | N/A | 8 |
| S2X259 | RBD (IIa) | 1–69 (94.1) | Sympt. (75) | 145 | 91 | Moderate | + | N/A | |
| S2X305 | RBD (?) | 1–2(95.1) | Sympt. (125) | 34 | 21 | Strong | N/A | ||
| S2X35 | RBD (IIa) | 1–18(98.6) | Sympt. (48) | 140 | 143 | Strong | + | N/A | |
| S2X450 | RBD (?) | 2–26 (96.9) | Sympt. (271) | 368 | 198 | Strong | + | N/A | |
| S2X475 | RBD (?) | 3–21 (93.8) | Sympt. (271) | 1′431 | 851 | Strong | + | N/A | |
| S2X607 | RBD (?) | 3–66 (95.4) | Sympt. (271) | 41 | 23 | Strong | N/A | ||
| S2X608 | RBD (?) | 1–33 (93.2) | Sympt. (271) | 21 | 35 | Strong | - | N/A | |
| S2X609 | RBD (?) | 1–69 (93.8) | Sympt. (271) | 47 | 35 | Strong | - | N/A | |
| S2X613 | RBD (I) | 1–2(91.7) | Sympt. (271) | 28 | 19 | Strong | N/A | ||
| S2X615 | RBD (I) | 3–11 (94.8) | Sympt. (271) | 23 | 17 | Strong | - | N/A | |
| S2X619 | RBD (?) | 1–69 (92.7) | Sympt. (271) | 36 | 60 | Strong | - | N/A | |
| S2X620 | RBD (?) | 3–53 (95.1) | Sympt. (271) | 34 | 45 | Strong | N/A |
id., identity. DSO, days after symptom onset. * as described in Piccoli et al and McCallum et al. N/A, not available; -, not neutralising
Acknowledgements
We would like to thank Cambridge University Hospitals NHS Trust Occupational Health Department. We would also like to thank the NIHR Cambridge Clinical Research Facility and staff at CUH and. We would like to thank Eleanor Lim and Georgina Okecha. We thank Dr James Voss for the kind gift of HeLa cells stably expressing ACE2. RKG is supported by a Wellcome Trust Senior Fellowship in Clinical Science (WT108082AIA). LEM is supported by a Medical Research Council Career Development Award (MR/R008698/1). SAK is supported by the Bill and Melinda Gates Foundation via PANGEA grant: OPP1175094. DAC is supported by a Wellcome Trust Clinical PhD Research Fellowship. KGCS is the recipient of a Wellcome Investigator Award (200871/Z/16/Z). This research was supported by the National Institute for Health Research (NIHR) Cambridge Biomedical Research Centre, the Cambridge Clinical Trials Unit (CCTU), and the NIHR BioResource. This study was supported by the National Institute of General Medical Sciences (R01GM120553 to D.V.), the National Institute of Allergy and Infectious Diseases (DP1AI158186 and HHSN272201700059C to D.V.), a Pew Biomedical Scholars Award (D.V.), an Investigators in the Pathogenesis of Infectious Disease Awards from the Burroughs Wellcome Fund (D.V.) and Fast Grants (D.V.). The views expressed are those of the authors and not necessarily those of the NIHR or the Department of Health and Social Care. JAGB is supported by the Medical Research Council (MC_UP_1201/16). IATM is funded by a SANTHE award.
Footnotes
Competing interests
A.D.M., J.B., D.P., C.S.F., S.B., K.C., N.S., E.C., G.S., S.J., A.L., H.W.V., M.S.P., L.P. and D.C. are employees of Vir Biotechnology and may hold shares in Vir Biotechnology. H.W.V. is a founder of PierianDx and Casma Therapeutics. Neither company provided funding for this work or is performing related work. D.V. is a consultant for Vir Biotechnology Inc. The Veesler laboratory has received a sponsored research agreement from Vir Biotechnology Inc. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest. RKG has received consulting fees from UMOVIS Lab, Gilead and ViiV.
References
- 1.Zhou P. et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 579, 270–273, doi: 10.1038/s41586-020-2012-7 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Davies N. G. et al. Estimated transmissibility and severity of novel SARS-CoV-2 Variant of Concern 202012/01 in England. medRxiv, 2020.2012.2024.20248822, doi: 10.1101/2020.12.24.20248822 (2020). [DOI] [Google Scholar]
- 3.Volz E. et al. Transmission of SARS-CoV-2 Lineage B.1.1.7 in England: Insights from linking epidemiological and genetic data. medRxiv, 2020.2012.2030.20249034, doi: 10.1101/2020.12.30.20249034 (2021). [DOI] [Google Scholar]
- 4.Korber B. et al. Tracking Changes in SARS-CoV-2 Spike: Evidence that D614G Increases Infectivity of the COVID-19 Virus. Cell 182, 812–827 e819, doi: 10.1016/j.cell.2020.06.043 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Yurkovetskiy L. et al. Structural and Functional Analysis of the D614G SARS-CoV-2 Spike Protein Variant. Cell 183, 739–751 e738, doi: 10.1016/j.cell.2020.09.032 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Martinot M. et al. Remdesivir failure with SARS-CoV-2 RNA-dependent RNA-polymerase mutation in a B-cell immunodeficient patient with protracted Covid-19. Clin Infect Dis, doi: 10.1093/cid/ciaa1474 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Kemp S. et al. Neutralising antibodies in Spike mediated SARS-CoV-2 adaptation. medRxiv, 2020.2012.2005.20241927, doi: 10.1101/2020.12.05.20241927 (2020). [DOI] [Google Scholar]
- 8.Thomson E. C. et al. The circulating SARS-CoV-2 spike variant N439K maintains fitness while evading antibody-mediated immunity. bioRxiv, 1–49, doi:papers3://publication/doi/10.1101/2020.11.04.355842 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Baden L. R. et al. Efficacy and Safety of the mRNA-1273 SARS-CoV-2 Vaccine. N Engl J Med, doi: 10.1056/NEJMoa2035389 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Polack F. P. et al. Safety and Efficacy of the BNT162b2 mRNA Covid-19 Vaccine. N Engl J Med 383, 2603–2615, doi: 10.1056/NEJMoa2034577 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Voysey M. et al. Safety and efficacy of the ChAdOx1 nCoV-19 vaccine (AZD1222) against SARS-CoV-2: an interim analysis of four randomised controlled trials in Brazil, South Africa, and the UK. Lancet 397, 99–111, doi: 10.1016/S0140-6736(20)32661-1 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Mulligan M. J. et al. Phase I/II study of COVID-19 RNA vaccine BNT162b1 in adults. Nature 586, 589–593, doi: 10.1038/s41586-020-2639-4 (2020). [DOI] [PubMed] [Google Scholar]
- 13.Corbett K. S. et al. SARS-CoV-2 mRNA Vaccine Development Enabled by Prototype Pathogen Preparedness. bioRxiv, 2020.2006.2011.145920, doi: 10.1101/2020.06.11.145920 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Folegatti P. M. et al. Safety and immunogenicity of the ChAdOx1 nCoV-19 vaccine against SARS-CoV-2: a preliminary report of a phase 1/2, single-blind, randomised controlled trial. Lancet 396, 467–478, doi: 10.1016/S0140-6736(20)31604-4 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Kemp S. A. et al. Recurrent emergence and transmission of a SARS-CoV-2 Spike deletion ΔH69/V70. bioRxiv, 2020.2012.2014.422555, doi: 10.1101/2020.12.14.422555 (2020). [DOI] [Google Scholar]
- 16.Tegally H. et al. Emergence and rapid spread of a new severe acute respiratory syndrome-related coronavirus 2 (SARS-CoV-2) lineage with multiple spike mutations in South Africa. medRxiv, 2020.2012.2021.20248640, doi: 10.1101/2020.12.21.20248640 (2020). [DOI] [Google Scholar]
- 17.Faria N. R. et al. Genomic characterisation of an emergent SARS-CoV-2 lineage in Manaus: preliminary findings, <https://virological.org/t/genomic-characterisation-of-an-emergent-sars-cov-2-lineage-in-manaus-preliminary-findings/586> (2021).
- 18.Jackson L. A. et al. An mRNA Vaccine against SARS-CoV-2 - Preliminary Report. N Engl J Med 383, 1920–1931, doi: 10.1056/NEJMoa2022483 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Walsh E. E. et al. Safety and Immunogenicity of Two RNA-Based Covid-19 Vaccine Candidates. New England Journal of Medicine 383, 2439–2450, doi: 10.1056/NEJMoa2027906 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Schmidt F. et al. Measuring SARS-CoV-2 neutralizing antibody activity using pseudotyped and chimeric viruses. 2020.2006.2008.140871, doi: 10.1101/2020.06.08.140871 %J bioRxiv (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Brouwer P. J. M. et al. Potent neutralizing antibodies from COVID-19 patients define multiple targets of vulnerability. Science 369, 643–650, doi: 10.1126/science.abc5902 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Wang P. L., L; Iketani S, Luo Y; Guo Y; Ho D. Increased Resistance of SARS-CoV-2 Variants B.1.351 and B.1.1.7 to Antibody Neutralization. bioXriv (2021). [Google Scholar]
- 23.PHE. Public Health England statement on Variant of Concern and new Variant Under Investigation, <https://www.gov.uk/government/news/phe-statement-on-variant-of-concern-and-new-variant-under-investigation> (2021).
- 24.McCallum M. et al. N-terminal domain antigenic mapping reveals a site of vulnerability for SARS-CoV-2. bioRxiv, doi: 10.1101/2021.01.14.426475 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Thomson E. C. et al. Circulating SARS-CoV-2 spike N439K variants maintain fitness while evading antibody-mediated immunity. Cell, doi: 10.1016/j.cell.2021.01.037 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Greaney A. J. et al. Comprehensive mapping of mutations to the SARS-CoV-2 receptor-binding domain that affect recognition by polyclonal human serum antibodies. Cell host & microbe, doi: 10.1016/j.chom.2021.02.003 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Andreano E. et al. SARS-CoV-2 escape in vitro from a highly neutralizing COVID-19 convalescent plasma. bioRxiv, 2020.2012.2028.424451, doi: 10.1101/2020.12.28.424451 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Walls A. C. et al. Structure, Function, and Antigenicity of the SARS- CoV-2 Spike Glycoprotein. Cell 181, 281–292.e286, doi:papers3://publication/doi/10.1016/j.cell.2020.02.058 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Guan Y. et al. Isolation and characterization of viruses related to the SARS coronavirus from animals in southern China. Science 302, 276–278, doi: 10.1126/science.1087139 (2003). [DOI] [PubMed] [Google Scholar]
- 30.Li W. et al. Efficient replication of severe acute respiratory syndrome coronavirus in mouse cells is limited by murine angiotensin-converting enzyme 2. J Virol 78, 11429–11433, doi: 10.1128/JVI.78.20.11429-11433.2004 (2004). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Li W. et al. Receptor and viral determinants of SARS-coronavirus adaptation to human ACE2. The EMBO journal 24, 1634–1643 (2005). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Starr T. N. et al. Deep Mutational Scanning of SARS-CoV-2 Receptor Binding Domain Reveals Constraints on Folding and ACE2 Binding. Cell 182, 1295–1310 e1220, doi: 10.1016/j.cell.2020.08.012 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Wang Z. et al. mRNA vaccine-elicited antibodies to SARS-CoV-2 and circulating variants. Nature, doi: 10.1038/s41586-021-03324-6 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Wibmer C. K. et al. SARS-CoV-2 501Y.V2 escapes neutralization by South African COVID-19 donor plasma. bioRxiv, doi: 10.1101/2021.01.18.427166 (2021). [DOI] [PubMed] [Google Scholar]
- 35.Verschoor C. P. et al. Microneutralization assay titres correlate with protection against seasonal influenza H1N1 and H3N2 in children. PloS one 10, e0131531, doi: 10.1371/journal.pone.0131531 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Kulkarni P. S., Hurwitz J. L., Simoes E. A. F. & Piedra P. A. Establishing Correlates of Protection for Vaccine Development: Considerations for the Respiratory Syncytial Virus Vaccine Field. Viral Immunol 31, 195–203, doi: 10.1089/vim.2017.0147 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Goddard N. L., Cooke M. C., Gupta R. K. & Nguyen-Van-Tam J. S. Timing of monoclonal antibody for seasonal RSV prophylaxis in the United Kingdom. Epidemiol Infect 135, 159–162, doi: 10.1017/S0950268806006601 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Mercado N. B. et al. Single-shot Ad26 vaccine protects against SARS-CoV-2 in rhesus macaques. Nature 586, 583–588, doi: 10.1038/s41586-020-2607-z (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.McMahan K. et al. Correlates of protection against SARS-CoV-2 in rhesus macaques. Nature, doi: 10.1038/s41586-020-03041-6 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Rathnasinghe R. et al. The N501Y mutation in SARS-CoV-2 spike leads to morbidity in obese and aged mice and is neutralized by convalescent and post-vaccination human sera. medRxiv, 2021.2001.2019.21249592, doi: 10.1101/2021.01.19.21249592 (2021). [DOI] [Google Scholar]
- 41.Wang Z. et al. mRNA vaccine-elicited antibodies to SARS-CoV-2 and circulating variants. bioRxiv, doi: 10.1101/2021.01.15.426911 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Muik A. et al. Neutralization of SARS-CoV-2 lineage B.1.1.7 pseudovirus by BNT162b2 vaccine-elicited human sera. Science, doi: 10.1126/science.abg6105 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Wu K. et al. mRNA-1273 vaccine induces neutralizing antibodies against spike mutants from global SARS-CoV-2 variants. bioXriv, doi: 10.1101/2021.01.25.427948 (2021). [DOI] [Google Scholar]
- 44.Xie X. et al. Neutralization of N501Y mutant SARS-CoV-2 by BNT162b2 vaccine-elicited sera. bioRxiv, 2021.2001.2007.425740, doi: 10.1101/2021.01.07.425740 (2021). [DOI] [Google Scholar]
- 45.Suryadevara N. et al. Neutralizing and protective human monoclonal antibodies recognizing the N-terminal domain of the SARS-CoV-2 spike protein. bioRxiv, 2021.2001.2019.427324, doi: 10.1101/2021.01.19.427324 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Soh W. T. et al. The N-terminal domain of spike glycoprotein mediates SARS-CoV-2 infection by associating with L-SIGN and DC-SIGN. bioRxiv, 1–30, doi:papers3://publication/doi/10.1101/2020.11.05.369264 (2020). [Google Scholar]
- 47.Greaney A. J. et al. Comprehensive mapping of mutations to the SARS-CoV-2 receptor-binding domain that affect recognition by polyclonal human serum antibodies. bioRxiv, 2020.2012.2031.425021, doi: 10.1101/2020.12.31.425021 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Greaney A. J. et al. Complete mapping of mutations to the SARS-CoV-2 spike receptor-binding domain that escape antibody recognition. Cell Host & Microbe (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Weisblum Y. et al. Escape from neutralizing antibodies by SARS-CoV-2 spike protein variants. Elife 9, e61312, doi: 10.7554/eLife.61312 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Corti D. et al. A neutralizing antibody selected from plasma cells that binds to group 1 and group 2 influenza A hemagglutinins. Science 333, 850–856, doi: 10.1126/science.1205669 (2011). [DOI] [PubMed] [Google Scholar]
- 51.Pinto D. et al. Cross-neutralization of SARS-CoV-2 by a human monoclonal SARS-CoV antibody. Nature 583, 290–295, doi: 10.1038/s41586-020-2349-y (2020). [DOI] [PubMed] [Google Scholar]
- 52.Tortorici M. A. et al. Ultrapotent human antibodies protect against SARS-CoV-2 challenge via multiple mechanisms. Science, doi: 10.1126/science.abe3354 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Gregson J. et al. HIV-1 viral load is elevated in individuals with reverse transcriptase mutation M184V/I during virological failure of first line antiretroviral therapy and is associated with compensatory mutation L74I. Journal of Infectious Diseases (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Forloni M., Liu A. Y. & Wajapeyee N. Creating Insertions or Deletions Using Overlap Extension Polymerase Chain Reaction (PCR) Mutagenesis. Cold Spring Harb Protoc 2018, doi: 10.1101/pdb.prot097758 (2018). [DOI] [PubMed] [Google Scholar]
- 55.Case J. B. et al. Neutralizing Antibody and Soluble ACE2 Inhibition of a Replication-Competent VSV-SARS-CoV-2 and a Clinical Isolate of SARS-CoV-2. Cell Host Microbe 28, 475–485 e475, doi: 10.1016/j.chom.2020.06.021 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Naldini L., Blomer U., Gage F. H., Trono D. & Verma I. M. Efficient transfer, integration, and sustained long-term expression of the transgene in adult rat brains injected with a lentiviral vector. Proceedings of the National Academy of Sciences of the United States of America 93, 11382–11388 (1996). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Gupta R. K. et al. Full length HIV-1 gag determines protease inhibitor susceptibility within in vitro assays. AIDS 24, 1651 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Mlcochova P. et al. Combined point of care nucleic acid and antibody testing for SARS-CoV-2 following emergence of D614G Spike Variant. Cell Rep Med, 100099, doi: 10.1016/j.xcrm.2020.100099 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Walls A. C. et al. Elicitation of Potent Neutralizing Antibody Responses by Designed Protein Nanoparticle Vaccines for SARS-CoV-2. Cell 183, 1367–1382 e1317, doi: 10.1016/j.cell.2020.10.043 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Chi X. et al. A neutralizing human antibody binds to the N-terminal domain of the Spike protein of SARS-CoV-2. Science, eabc6952–6913, doi:papers3://publication/doi/10.1126/science.abc6952 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Tortorici M. A. et al. Ultrapotent human antibodies protect against SARS-CoV-2 challenge via multiple mechanisms. Science 4, eabe3354–3316, doi:papers3://publication/doi/10.1126/science.abe3354 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Piccoli L. et al. Mapping neutralizing and immunodominant sites on the SARS-CoV-2 spike receptor-binding domain by structure-guided high-resolution serology. Cell, 1–55, doi:papers3://publication/doi/10.1016/j.cell.2020.09.037 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The neutralization and BLI data shown in Fig. 4 and Extended Data Fig. 5–7 can be found in Source Data Fig. 4. Other data are available from the corresponding author on request.