
Figure 2.
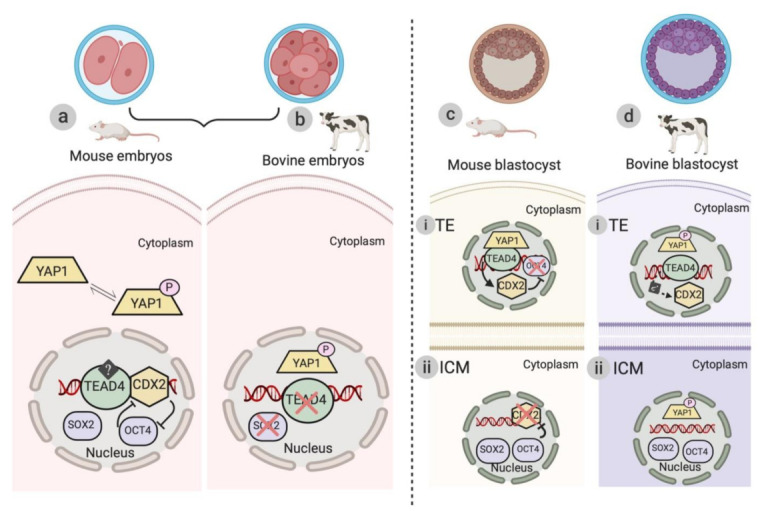
Schematic illustration of the differences in the localization of downstream effectors of the Hippo signaling pathway during murine and bovine embryogenesis. (a) Mechanism of activation of TEAD4 and other transcription factors (CDX2, SOX, and OCT4) of the Hippo signaling pathway during pre-compaction stages (from two-cell to eight-cell stage) of mouse preimplantation embryogenesis. The arrows represent the inhibitory mechanism between CDX2 and OCT4 in the murine model. (b) Protein localization of downstream effectors (p-YAP1, TEAD4, and SOX2) of the Hippo signaling pathway. p-YAP1 is localized in the nucleus and untraceable amounts of TEAD4 and SOX2 are detected during the pre-compaction stages of early bovine embryogenesis. (c and d) Localization of YAP1/p-YAP1 and TEAD4 transcription factors (CDX2, SOX2, and OCT4) in TE (trophectoderm) and ICM (inner cell mass) during blastocyst formation in murine and bovine models, respectively. (c-i) In the outer polar TE cells (Hippo “Off”), YAP1 in collaboration with TEAD4 causes the transcription of CDX2. OCT4 expression is thus inhibited by CDX2, aiding in the process of TE differentiation. (c-ii) In the apolar ICM cells, the Hippo signaling pathway is active (Hippo “On”) and SOX2 and OCT4 are exclusively present in the nucleus. In the ICM, OCT4 supresses the expression of CDX2 and facilitates the formation of ICM. (d-i and ii) Protein localization of downstream effectors (p-YAP1) and TEAD4 transcription factors (CDX2, SOX2, and OCT4) of the Hippo signaling pathway in TE and ICM during bovine blastocyst formation. The dotted line represents the proposed mechanism of TEAD4 and CDX2 interaction during bovine TE differentiation.